Manzamenones Inhibit T-Cell Protein Tyrosine Phosphatase

Abstract
:1. Introduction

2. Results and Discussion
3. Experimental Section
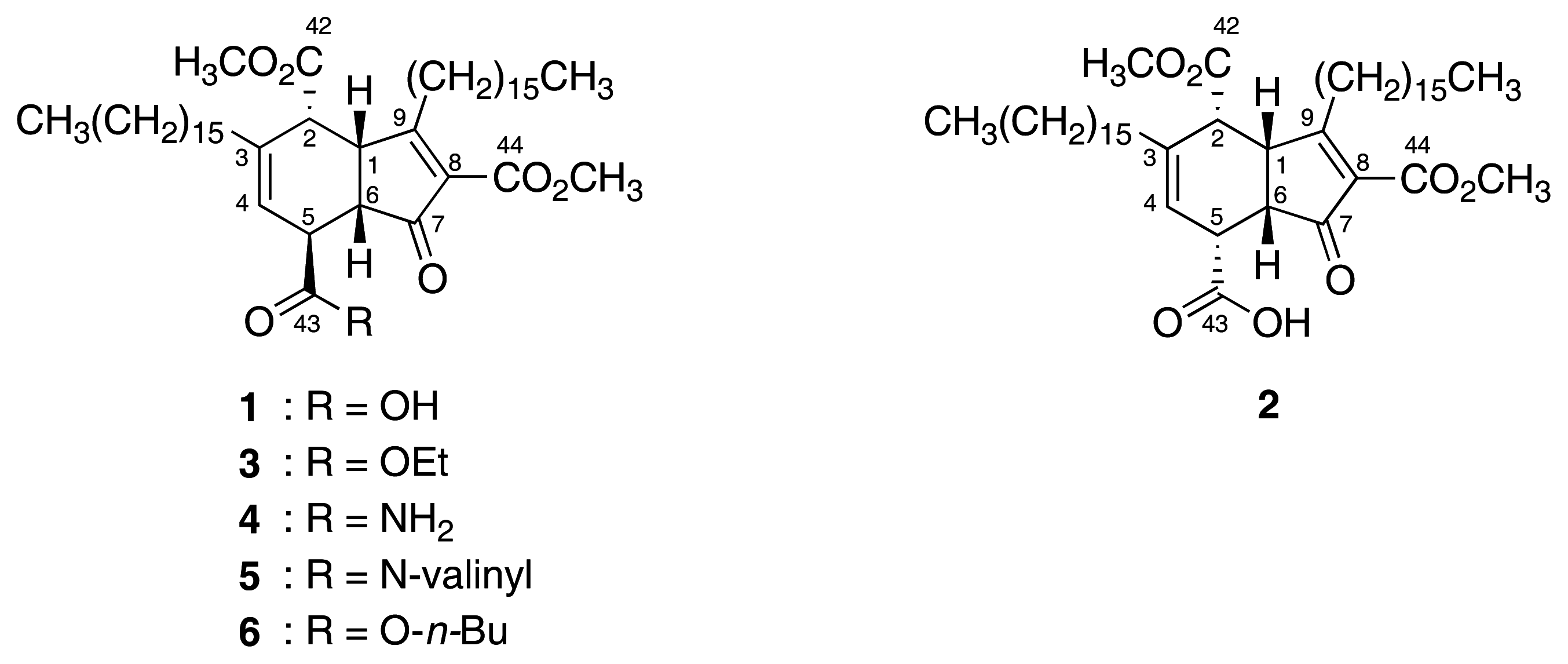
3.1. Compounds
3.2. Bacterial expression of recombinant proteins
3.3. Inhibitory activity against protein tyrosine phosphatases
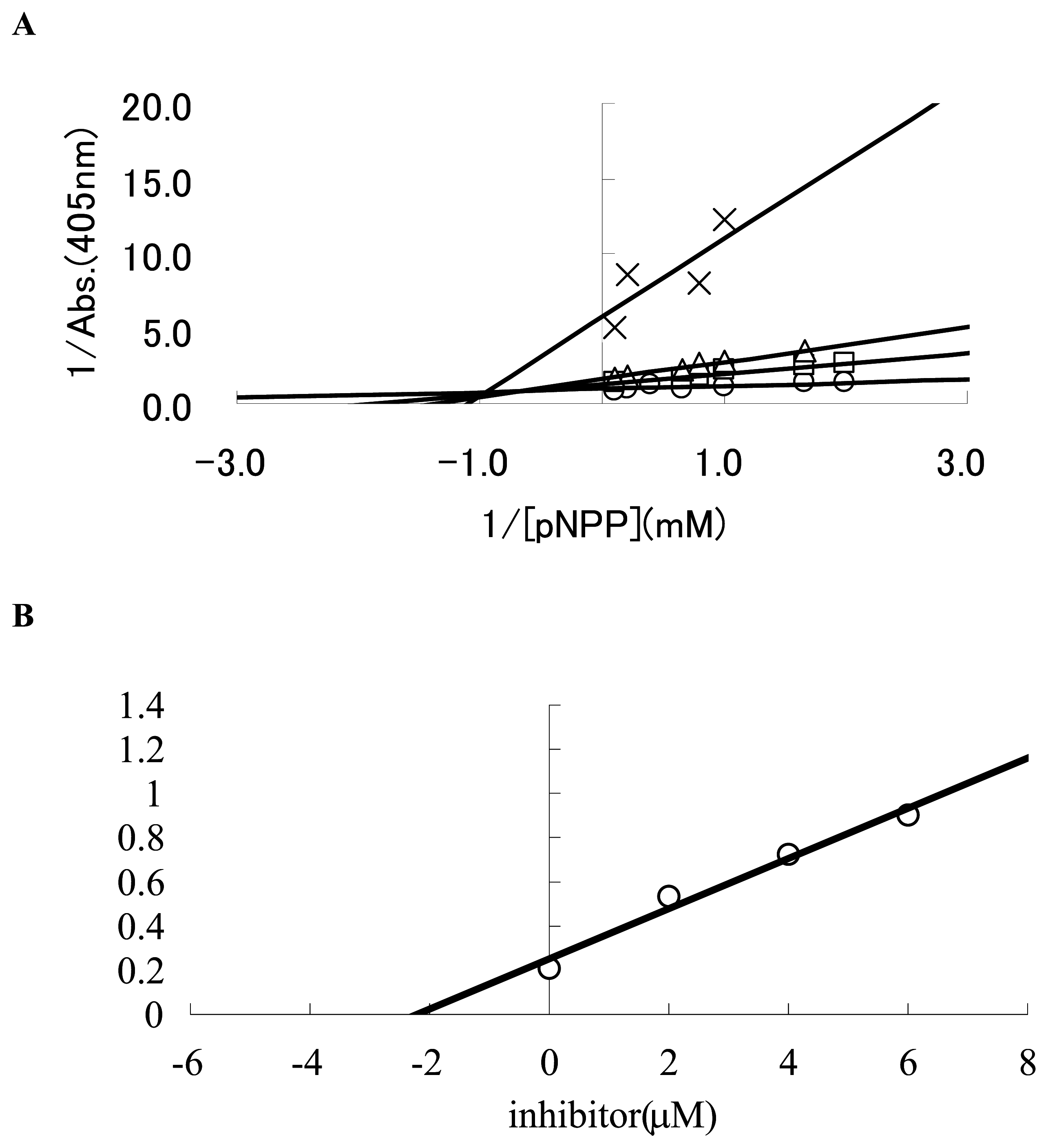
3.4. Determination of Ki values
Acknowledgements
References and Notes
- Tonks, N. K.; Neel, B. G. Combinatorial control of the specificity of protein tyrosine phosphatases. Curr. Opin. Cell. Biol 2001, 13, 182–195. [Google Scholar]
- Zhang, Z. Y. Protein tyrosine phosphatases: prospects for therapeutics. Curr. Opin. Chem. Biol 2001, 5, 416–423. [Google Scholar]
- Alonso, A.; Sasin, J.; Bottini, N.; Friedberg, I.; Friedberg, I.; Osterman, A.; Godzik, A.; Hunter, T.; Dixon, J.; Mustelin, T. Protein tyrosine phosphatases in the human genome. Cell 2004, 117, 699–711. [Google Scholar]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A. L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C. C.; Ramachandran, C.; Gresser, M. J.; Tremblay, M. L.; Kennedy, B. P. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar]
- Klaman, L. D.; Boss, O.; Peroni, O. D.; Kim, J. K.; Martino, J. L.; Zabolotny, J. M.; Moghal, N.; Lubkin, M.; Kim, Y. B.; Sharpe, A. H.; Stricker-Krongrad, A.; Shulman, G. I.; Neel, B. G.; Kahn, B. B. Increased energy expenditure, decreased adiposity, and tissue-specific insulin sensitivity in protein-tyrosine phosphatase 1B-deficient mice. Mol. Cell. Biol 2000, 20, 5479–5489. [Google Scholar]
- Tonks, N. K. PTP-1B: from the sidelines to the front lines. FEBS Lett 2003, 546, 140–148. [Google Scholar]
- Zhang, Z. Y. Protein tyrosine phosphatases: structure and function, substrate specificity, and inhibitor development. Annu. Rev. Pharmacol. Toxicol 2002, 42, 209–234. [Google Scholar]
- Andersen, J. N.; Jansen, P. G.; Echwald, S. M.; Mortensen, O. H.; Fukada, T.; Del Vecchio, R.; Tonks, N. K.; Moller, N. P. A genomic perspective on protein tyrosine phosphatases: gene structure, pseudogenes, and genetic disease linkage. FASEB J 2004, 18, 8–30. [Google Scholar]
- Heinonen, K. M.; Nestel, F. P.; Newell, E. W.; Charette, G.; Seemayer, T. A.; Tremblay, M. L.; Lapp, W. S. T-cell protein tyrosine phosphatase deletion results in progressive systemic inflammatory disease. Blood 2004, 103, 3457–3464. [Google Scholar]
- Galic, S.; Hauser, C.; Kahn, B. B.; Haj, F. G.; Neel, B. G.; Tonks, N. K.; Tiganis, T. Coordinated regulation of insulin signaling by the protein tyrosine phosphatases PTP1B and TCPTP. Mol. Cell. Biol 2005, 25, 819–829. [Google Scholar]
- Tsukamoto, S.; Takeuchi, S.; Ishibashi, M.; Kobayashi, J. Manzamenones A ~ F from the Okinawan marine sponges Plakortis sp.: novel dimeric fatty acid-derivatives possessing a bicyclo[4.3.0]nonane skeleton. J. Org. Chem 1992, 57, 5255–5260. [Google Scholar]
- Whitehead and co-workers have accomplished the total synthesis of manzamenones A, C, and F, and revised the stereostructure of manzamenone A to be 1 Al-Busafi, S.; Drew, M. G. B.; Sanders, T.; Whitehead, R. C. Pre-disposition in synthesis: an approach to the manzamenones. Tetrahedron Lett. 1998, 39, 1647–1650. [Google Scholar]Al-Busafi, S.; Whitehead, R. C. Predisposition in synthesis: efficient routes to (±)-untenone A and (±)-manzamenones A, C and F. Tetrahedron Lett. 2000, 41, 3467–3470. [Google Scholar]
- Aoki, N.; Matsuda, T. A cytosolic protein tyrosine phosphatase PTP1B specifically dephosphorylates and deactivates prolactin-activated STAT5a and STAT5b. J. Biol. Chem 2000, 275, 39718–39726. [Google Scholar]
- Matsuzawa, S.; Tamura, T.; Mizuno, Y.; Kobayashi, S.; Okuyama, H.; Tsukitani, Y.; Uemura, D.; Kikuchi, K. Increase in potential activities of protein phosphatases PP1 and PP2A in lymphoid tissues of autoimmune MRL/MpJ-lpr/lpr mice. J. Biochem 1992, 111, 472–477. [Google Scholar]
- Sample availability: not available.



{kind=link}
{kind=link}
{kind=link}
| IC50 values (μM) | ||||
|---|---|---|---|---|
| Compound | TCPTP | PTP1B | PP1/PP2A | A-kinase |
| 1 | 9.1 | >20 | >50 | >50 |
| 2 | 2.5 | 10.8 | >50 | >50 |
| 3 | 7.6 | >20 | >50 | >50 |
| 4 | >20 | >20 | - | - |
| 5 | 3.2 | 13.5 | >50 | >50 |
| 6 | 12.6 | >20 | - | - |
© 2006 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Wakuda, Y.; Kubota, T.; Shima, H.; Okada, T.; Mitsuhashi, S.; Aoki, N.; Kikuchi, K.; Kobayashi, J. Manzamenones Inhibit T-Cell Protein Tyrosine Phosphatase. Mar. Drugs 2006, 4, 9-14. https://doi.org/10.3390/md401009
Wakuda Y, Kubota T, Shima H, Okada T, Mitsuhashi S, Aoki N, Kikuchi K, Kobayashi J. Manzamenones Inhibit T-Cell Protein Tyrosine Phosphatase. Marine Drugs. 2006; 4(1):9-14. https://doi.org/10.3390/md401009
Chicago/Turabian StyleWakuda, Yusuke, Takaaki Kubota, Hiroshi Shima, Tadashi Okada, Shinya Mitsuhashi, Naohito Aoki, Kunimi Kikuchi, and Jun\'ichi Kobayashi. 2006. "Manzamenones Inhibit T-Cell Protein Tyrosine Phosphatase" Marine Drugs 4, no. 1: 9-14. https://doi.org/10.3390/md401009