Tetrodotoxin – Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication

1
Tokyo Health Care University, 3-11-3, Setagaya, Tokyo 154-8568, Japan
2
Faculty of Fisheries, Nagasaki University, 1-14, Bunkyo-machi, Nagasaki 852-8521, Japan
*
Author to whom correspondence should be addressed.
Mar. Drugs 2008, 6(2), 220-242; https://doi.org/10.3390/md6020220
Submission received: 23 December 2007
/
Revised: 24 March 2008
/
Accepted: 8 April 2008
/
Published: 28 May 2008
(This article belongs to the Special Issue Marine Toxins)
Abstract
:Many pufferfish of the family Tetraodontidae possess a potent neurotoxin, tetrodotoxin (TTX). In marine pufferfish species, toxicity is generally high in the liver and ovary, whereas in brackish water and freshwater species, toxicity is higher in the skin. In 1964, the toxin of the California newt was identified as TTX as well, and since then TTX has been detected in a variety of other organisms. TTX is produced primarily by marine bacteria, and pufferfish accumulate TTX via the food chain that begins with these bacteria. Consequently, pufferfish become non-toxic when they are fed TTX-free diets in an environment in which the invasion of TTX-bearing organisms is completely shut off. Although some researchers claim that the TTX of amphibians is endogenous, we believe that it also has an exogenous origin, i.e., from organisms consumed as food. TTX-bearing animals are equipped with a high tolerance to TTX, and thus retain or accumulate TTX possibly as a biologic defense substance. There have been many cases of human intoxication due to the ingestion of TTX-bearing pufferfish, mainly in Japan, China, and Taiwan, and several victims have died. Several cases of TTX intoxication due to the ingestion of small gastropods, including some lethal cases, were recently reported in China and Taiwan, revealing a serious public health issue.
1. Introduction
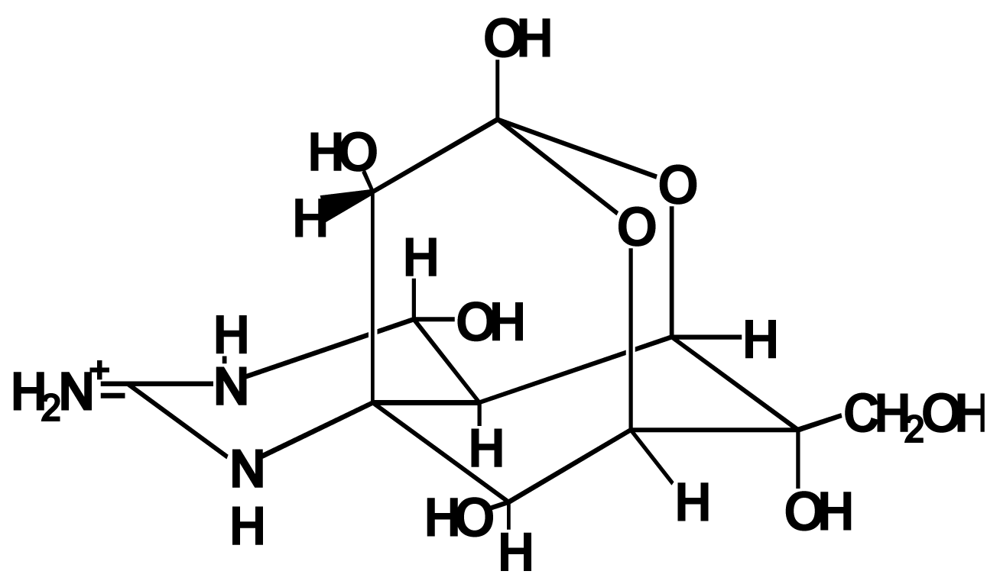
Pufferfish toxin (tetrodotoxin, TTX) is a potent neurotoxin of low molecular weight, which was first isolated in 1950 as a crystalline prism from toxic pufferfish by Yokoo [1]. Its structure (Figure 1) was elucidated by three groups in 1964 [2–4]. TTX inhibits nerve and muscle conduction by selectively blocking sodium channels [5], resulting in respiratory paralysis that causes death. The lethal potency is 5000 to 6000 MU/mg [1 MU (mouse unit) is defined as the amount of toxin required to kill a 20-g male mouse within 30 min after intraperitoneal administration], and the minimum lethal dose (MLD) for humans is estimated to be approximately 10000 MU ( 2 mg) [6]. Various TTX derivatives have so far been separated from pufferfish and/or some other TTX-bearing organisms [7,8].
Many years ago when TTX was believed to be found exclusively in pufferfish, it was controversial whether TTX in the fish was endogenous (produced by the pufferfish itself) or exogenous (taken from the outside and accumulated). Subsequent findings, however, such as the distribution of TTX among many other organisms [9], the TTX intoxication of the trumpet shell following the ingestion of toxic starfish [10], TTX production by marine bacteria [11], and the facts pufferfish become non-toxic when artificially reared with non-toxic diets [12–14] and that such non-toxic pufferfish become toxic when fed TTX-containing diets [15–17], have elucidated that the main mechanism of TTX accumulation in pufferfish is the food chain, consisting of several steps and starting with marine bacteria as a primary source of TTX.
In the present paper, the detailed mechanisms of the distribution and accumulation of TTX are described. We also present some case reports of human intoxication, such as via pufferfish poisoning in Japan, and food poisoning due to the ingestion of small gastropods, which was recently reported in China and Taiwan.
2. TTX distribution in aquatic organisms
2.1. Distribution of TTX in pufferfish species
The toxicity of the Japanese marine pufferfish was extensively studied by Tani, who reported that 14 of the 21 species examined were toxic [18]. Later, 8 pufferfish species were added to the list of toxic species [19–22], and a total of 22 species are presently listed as TTX-bearing marine pufferfish (Table 1), all belonging to the Tetraodontidae family.
Lagocephalus wheeleri and L. gloveri of the same family are usually regarded as non-toxic species, although they occasionally show weak toxicity [23]. All species of the Diodontidae and Ostraciontidae families are TTX-free, though the skin of Ostraciontidae contains an ichthyotoxic and hemolytic substance called pahutoxin [24,25]. Food poisoning cases due to ingestion of the liver of Ostraciontid boxfish sometimes occur in Japan, but the causative agent is a toxin with delayed hemolytic activity, possibly palytoxin or a similar substance, and not TTX or pahutoxin [26]. Thai brackish water pufferfish Tetraodon nigroviridis and T. steindachneri, and freshwater pufferfish T. fangi, T. leiurus, and T. suwatii are also toxic. The toxin of brackish water species was identified as TTX [27,28], but in the freshwater species, saxitoxins (STXs), toxins that belong to the paralytic shellfish poison (PSP) family, were detected as the main toxic principles [29,30]. Freshwater pufferfish, Colomesus asellus from Brazil [31] and T. turgidus from Cambodia [32], also possess PSP. Recently, some marine pufferfish (Arothron spp.) from the Philippines [33], and Arothron firmamentum from Japanese waters [34] were also reported to possess STXs as the major toxins. Furthermore, palytoxin-like substance(s), in addition to a small amount of PSP, were detected in a Bangladeshi freshwater pufferfish belonging to the genus Tetraodon [35].
The generally large individual and regional variations in pufferfish toxicity [20] are too great to be explained by endogenous factors alone, i.e., differences in physiologic conditions among individual pufferfish, suggesting their exogenous intoxication.
2.2. Distribution of TTX in pufferfish bodies
The distribution of TTX in pufferfish bodies appears to be species-specific (Table 1). In marine species, the liver and ovary generally have the highest toxicity, followed by intestines and skin. Muscles and/or testis are non-toxic or weakly toxic, except for in Lagocephalus lunaris and Chelonodon patoca, and are regarded as edible by the Japanese Ministry of Health, Labour, and Welfare, even in many toxic species. In contrast, in C. patoca inhabiting coastal or brackish waters of the Okinawa and Amami Islands [36], and in Thai/Bangladeshi/Cambodian brackish water and freshwater pufferfish, the skin has the highest toxicity [27–29,37].
Our recent immunohistochemical investigations using a monoclonal anti-TTX antibody clarified the intratissue microdistribution of TTX in the skin and ovary of TTX-bearing pufferfish [38–40]. Species-specificity was also recognized in the microdistribution, i.e., TTX was localized in the nucleus, yolk vesicles, and yolk granules of oocytes in the ovary of the marine species T. vermicularis, whereas it was detected in the connective tissues and nucleus of some oocytes in C. patoca [39]. TTX-bearing glands were observed in the skin of T. vermicularis, and TTX-bearing secretory cells (succiform cells) were observed in brackish water species such as C. patoca, T. steindachneri, and T. nigroviridis [38–40].
In the toxic marine pufferfish, the liver generally shows very high toxicity throughout the year except in the spawning season, during which the ovary becomes highly toxic by accumulating TTX transferred from the liver [20]. TTX in the eggs spawned from the ovary could be involved in protecting the eggs from predators. On the other hand, when toxic pufferfish encounter enemies, their bodies swell to two or three times their usual size and TTX is excreted from their skin to repel the enemies [41,42]. These observations suggest that the pufferfish accumulate TTX as a biologic defense agent.
2.3. Distribution of TTX in animals other than pufferfish
TTX was long believed to be present only in pufferfish. Since Mosher et al. detected TTX in the eggs of the California newt Taricha torosa in 1964 [43], however, TTX has been detected in a wide variety of animals, as shown in Table 2 [9].
It is quite unlikely that these TTX-bearers belonging to particular species of different phyla possess a common gene that codes for TTX production. The trumpet shell Charonia sauliae accumulates TTX by ingesting toxic starfish [10], supporting the hypothesis that the TTX of pufferfish is not endogenous, but comes via the food chain. The exact origin of TTX in the food chain, however, remains unknown. Because the ecologic environments of TTX-bearing animals seem to have no common factor other than being closely related to an aquatic system, bacteria, omnipresent organisms that commonly inhabit the aquatic system, are implicated as the primary source of TTX.
3. Mechanism of TTX accumulation in TTX-bearing organisms
3.1. TTX producers
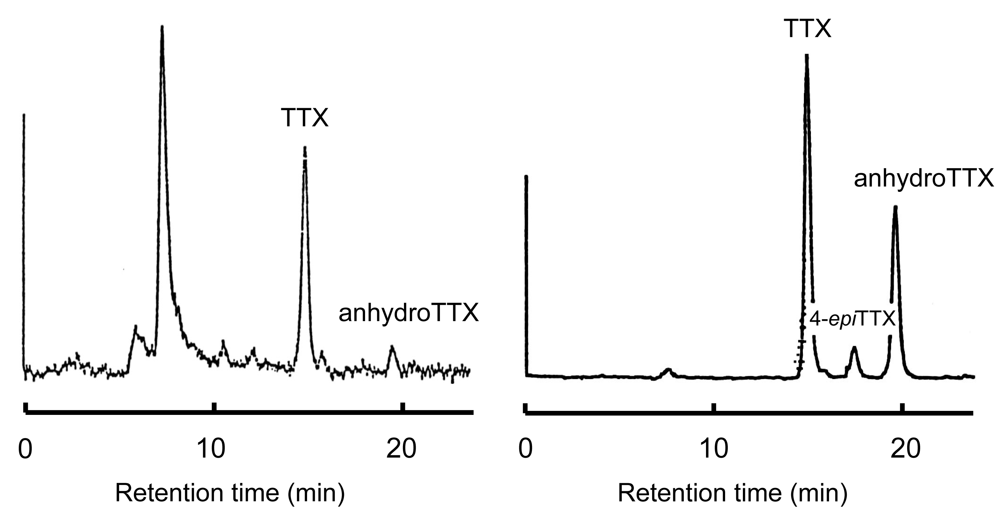
Based on the above suggestion that bacteria might be the primary source of TTX, in the 1980s we investigated the TTX production of marine bacteria. Intestinal bacteria from several TTX-bearing organisms were isolated, cultured, and examined for lethal potency to mice. Vibrio alginolyticus from the toxic starfish Astropecten polyacanthus [73,74], one Vibrio strain from the xanthid crab Atergatis floridus [11,74], and another Vibrio strain from the toxic marine pufferfish Takifugu snyderi [74] had a paralytic toxicity of 213, 30, and 3 MU per 500 ml of medium, respectively. The toxic principles produced peaks identical to those of authentic TTX and anhydrotetrodotoxin (anhydroTTX) in high performance liquid chromatography with post-column fluorescence derivatization (Figure 2)[74]. In addition, gas chromatography/mass spectrometry analyses revealed that they produce a C9-base, the degradation product of TTX, on alkaline hydrolysis [75], leading us to conclude that the isolated bacteria produce TTX. Shewanella alga and Alteromonas tetraodonis isolated from a red alga Jania sp. [76], Shewanella putrefaciens from a TTX-bearing marine puffer Takifugu niphobles [77], and some other marine bacteria isolated from TTX-bearing organisms [78] also produce TTX.
3.2. Mechanism of TTX accumulation in pufferfish
As mentioned above, many years of studies on TTX have revealed that (1) pufferfish toxicity shows remarkable individual and regional variations, (2) TTX is distributed over various organisms, including food animals of pufferfish, (3) the trumpet shell accumulates TTX by ingesting toxic starfish, and (4) marine bacteria primarily produce TTX, suggesting that pufferfish do not synthesize TTX, but accumulate it through the food chain, which starts from marine bacteria. If this is the case, pufferfish should become non-toxic when fed TTX-free diets in an environment in which the invasion of TTX-bearing organisms has been eliminated.
To test this hypothesis, we collected more than 6000 specimens of the marine pufferfish Takifugu rubripes that were cultured with non-TTX-containing diets in such environments (net cages on the sea or land aquaria located in eight prefectures of Japan) and examined the toxicity of their livers and some other body parts, including gonads and muscle, using an accepted TTX bioassay [14]. All specimens were non-toxic, i.e., their toxicity was under the detection limit (2–10 MU/g) (Tables 3 and 4). Furthermore, liquid chromatography/mass spectrometry analysis demonstrated that none of the 20 liver specimens from Nagasaki and 5 liver specimens from Yobuko contained detectable amounts of TTX, i.e., their TTX content was below 0.1–0.4 MU/g.
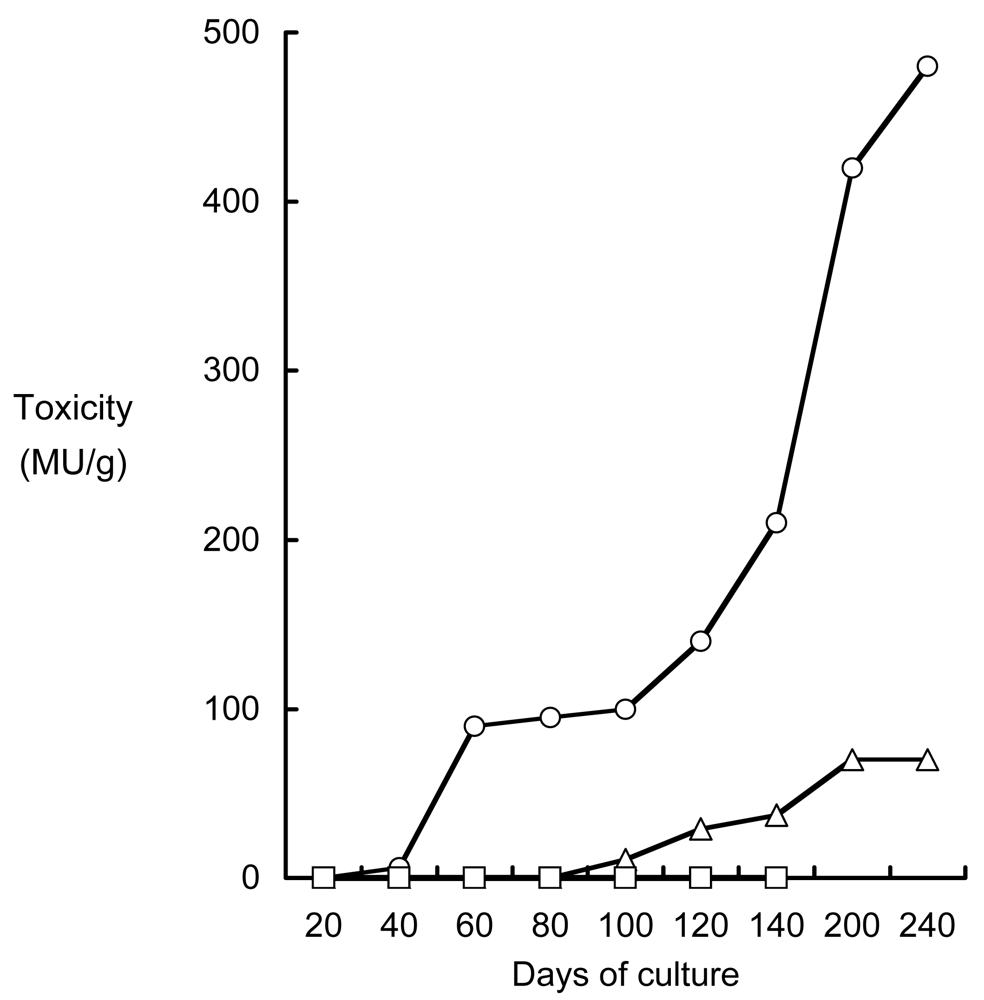
When non-toxic cultured pufferfish T. rubripes were reared on TTX-containing diets, however, they became toxic within 40 days when fed 4 MU TTX/g body mass/day, and in 100 days when fed 0.5 MU TTX/g body mass/day. Thereafter, the toxicity continuously increased up to the end of experimental period (Figure 3) [79]. In contrast, three non-pufferfish species were not intoxicated with TTX, even after culturing for 139 days when fed 4 MU TTX/g body mass/day. These species seem not only to exclude, but also to detoxify, TTX.
In a similar feeding experiment using a PSP [consisting of STX and neosaxitoxin (neoSTX)]-containing diet for 60 days, non-toxic pufferfish were also intoxicated with PSP. Their toxicity scores, however, were only 21% to 53% of those of the fish fed with the same dosage of TTX as controls (unpublished data). Thus, the TTX-bearing marine pufferfish tend to selectively accumulate more TTX than PSP.
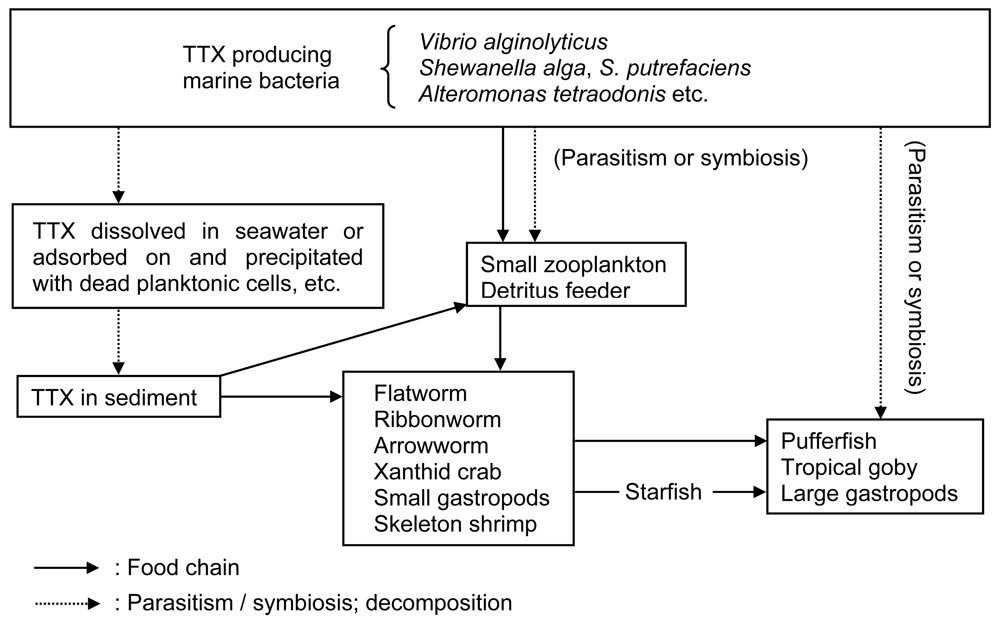
These results clearly demonstrate that the TTX of toxic pufferfish is not endogenous, but is derived from the food chain (Figure 4). Another possibility is that the TTX is produced by symbiotic or parasitic bacteria that are directly accumulated inside the puffer body and not obtained via the food chain. The amount of TTX produced by bacteria, however, appears to be too little to account for the TTX accumulation in pufferfish, therefore bioconcentration likely has a large role in the accumulation of TTX in pufferfish. Furthermore, based on the observation that 6000 specimens of pufferfish reared over 1 year in net cages or land aquaria were non-toxic, the TTX produced in the intestines of pufferfish by marine bacteria must be a minor contributor, or rather negligible, to account for the TTX accumulation in pufferfish.
3.3. Mechanism of TTX accumulation in amphibians
Some studies of the intoxication mechanism have been performed in amphibians, especially newts. Shimizu and Kobayashi [80] first observed that when various radioactive precursors were administered to two species of newts, Taricha torosa and T. granulosa, none of the precursors were incorporated into TTX molecules, although the authors did not clearly conclude whether TTX in the newts was endogenous or exogenous. On the other hand, Hanifin et al. claim that newts can biosynthesize TTX by themselves, because TTX levels in the dorsal skin of T. granulosa significantly increased under artificial rearing with non-toxic diets for 1 year [81], and because the newts released a large amount of TTX from their skin after receiving an electric shock, but the TTX levels in the dorsal skin significantly recovered during the 9 months thereafter [82].
During our studies on the toxicity and toxin profiles of the Japanese newt Cynops pyrrhogaster [83,84], we observed that the eggs of the newt retained a certain amount of TTX inherited from their parents, which gradually disappeared in the larval stage. In the juvenile stage in a terrestrial environment, the newt abruptly begins to accumulate TTX again, and becomes highly toxic in the adult stage. When artificially reared with non-toxic diets for 5 years after hatching, however, the newt became non-toxic (< 2 MU/g). The non-toxic newt, when orally administered TTX or 6-epitetrodotoxin (6-epiTTX), accumulated approximately 50% of the administered toxins. These observations strongly suggest that the TTX of C. pyrrhogaster is exogenous, i.e., the newt begins to accumulate TTX after metamorphosis, with their food organisms as its origin.
Daly et al. [85] examined the intoxication mechanism of frogs. They hatched eggs of the Panamanian frog Atelopus varius which possesses both bufadienolides (a type of steroid) and TTX in the skin, and artificially reared them for 25 to 45 months. In the skin of the adult frogs, bufadienolides were detected at the same level as in wild specimens, but TTX was not detected, leading them to conclude that bufadienolides are endogenous and the TTX is exogenous, possibly derived from food organisms or environmental factors such as symbiotic bacteria.
3.4. Resistance of animals to TTX
Three species each of toxic pufferfish [T. niphobles, T. pardalis, and T. rubribes (cultured specimens)], non-toxic pufferfish (L. wheeleri, L. gloveri, and Liosaccus cutaneus), and non-toxic general fish (Oplegnathus punctatus, O. fasciatus, and Girella punctata) were examined for their resistance to TTX [86]. The MLD of TTX administered intraperitoneally to the three toxic species of pufferfish was 300 to 750 times greater than that of mice (Table 5). Due to this high resistance, the Takifugu species can accumulate huge amounts of TTX. Non-toxic pufferfish showed 13 to 15 times stronger resistance against TTX than mice. These pufferfish are generally non-toxic, but can become weakly toxic in some habitats [23]. They can accumulate a limited amount of TTX due to the medium level of resistance to TTX. In contrast, general non-toxic fish had the same low resistance levels as mice, i.e., they cannot accumulate TTX. The goby Yongeichthys criniger (formerly Gobius criniger) also has high resistance to TTX, and can accumulate TTX [63]. TTX-bearing xanthid crabs [87] and the Japanese newt [84] exhibit even greater resistance to TTX than pufferfish. Based on these findings, it is clear that for TTX-bearing animals to accumulate a certain level of TTX in their body, they must be endowed with high tolerance to TTX.
The mechanism of TTX resistance in pufferfish and rewts has been explained based mainly on the TTX-resistant sodium channels found in the animals, in which the aromatic amino acid commonly located in the p-loop region of domain I in TTX-sensitive sodium channels is replaced by a nonaromatic amino acid, resulting in their extremely low affinity to TTX [88–91]. Garter snakes and clams can also acquire similar sodium channel mutation-based TTX/PSP resistance by interacting with their toxic food organisms, TTX-bearing newts and PSP-producing dinoflagellates, respectively [92,93]. On the other hand, the shore crab Hemigrapsus sanguineus possesses TTX-binding proteins in its hemolymph, which confer TTX resistance on this non-toxic crab [94,95]. Pufferfish and newts may be equipped not only with TTX-resistant sodium channels, but also with other mechanisms whereby they mask and/or modulate TTX molecules inside their body.
4. Cases of human intoxication by TTX
4.1. Poisonings due to pufferfish
4.1.1. Cases in Japan
Many Japanese know that pufferfish, especially their livers (“kimo”), are very toxic. Nevertheless, there are more than a few “kimo” fans who dare to ingest the liver, believing that the toxin can be eliminated by their own special or traditional detoxification methods. Accordingly, food poisonings due mainly to ingestion of the liver were occurring very frequently in Japan [6]. This situation prompted the Japanese Ministry of Health and Welfare (presently the Ministry of Health, Labour, and Welfare) to publish a guideline for edible pufferfish in 1983, with updates in 1993 and 1995 (Table 6), and to prohibit pufferfish liver from being served in restaurants or in the market. Many cases of puffer poisoning, however, continue to occur every year due to the consumption of homemade “kimo” dishes, which are prepared using wild fish that is not purchased commercially. In Japan, over the past 13 years (1995–2007), mortality due to pufferfish poisoning has been 6.4% (Table 7).
Case report of pufferfish poisoning [96]
In the early morning of a day in October 1996, a 48-year-old man in Nagasaki, Nagasaki Prefecture, caught a wild marine pufferfish Takifugu poecilonotus, and ate more than four slices of slightly cooked “kimo” with some flesh in the evening. Thirty to 60 minutes after ingestion, he began to suffer from numbness in his hands and limbs, followed by cyanosis and respiratory failure during the next 60 minutes. Although immediately hospitalized, he died during the following hour.
In this case, toxicity scores of the leftover liver and flesh were determined to be 715-4260 and less than 5 MU/g, respectively, and TTX was identified as the toxic principle [96]. The victim was estimated to have ingested more than 10000 MU, the amount equivalent to the MLD of TTX in humans, and the cause of his death was concluded to be TTX contained in the liver of wild T. poecilonotus.
4.1.2. Cases in Taiwan/China
In Taiwan and China, though people do not eat pufferfish as often as the Japanese, many food poisoning cases due to ingestion of wild pufferfish have also occurred. According to the record of TTX poisoning in Taiwan, there are some cases caused by the mistaken ingestion of muscles of a pufferfish species with toxic muscle, by ingesting puffer roe that had been sold as a fake of dried mullet roe called “karasumi”, or by ingesting a dried dressed fish fillet produced from toxic pufferfish by a food processing company [97–99].
Case report of poisoning cases in Taiwan [100]
A food poisoning incident following fish ingestion occurred in Chunghua Prefecture, western Taiwan in January 2000. A total of five victims (4 men, 58–64 years old and 1 woman, 46 years old) were reported. Symptoms included paralysis, coma, nausea, vomiting, ataxia, aphasia, and difficult respiration. Among these victims, two men suffered from more serious symptoms and were treated with intravenous fluids, mechanical ventilation, and intensive treatment in the hospital. They were then discharged uneventfully after 1 week of management.
According to the victims, the causative fish, which had been collected from the coastal area of Chunghua Prefecture, might be the marine pufferfish Takifugu niphobles. They retained a small piece (about 11 g) of the cooked fish liver, which, along with eight live specimens of T. niphobles, was then assayed for toxicity. The results indicated that the toxicity of the cooked fish liver was 280 MU/g, and all the T. niphobles specimens had high toxicity (>850 MU/g) in their liver [100]. The toxin of both samples was identified as TTX. Mitochondrial DNA of the leftover and T. niphobles specimens showed the same sequence genotype and the same single restriction site for BsaI, indicating that T. niphobles was responsible for the intoxication caused by TTX [101].
4.1.3. Cases in other countries
Ten human fatalities attributable to the consumption of marine pufferfish have been reported in the United States [102], among which four cases occurred in Hawaii during 1908–1925. The Arothron sp. was responsible for all the poisonings [103]. In 1986, a further poisoning incident occurred in Hawaii due to consumption of the liver of Diodon hystrix, affecting one man [104]. Although he was not admitted to the hospital until 24 hours after exposure, he recovered within 1 week. There is also a case, in which two Dutch sailors died within 17 to 20 minutes after ingesting the liver of a South African marine pufferfish [6].
In Bangladesh, a food poisoning incident due to ingestion of the roe of the marine pufferfish Takifugu oblongus occurred in November 1998, affecting eight people and resulting in five deaths [105]. Their symptoms were dyspnea, numbness of the lips, paralysis, and stomachache followed by vomiting, which appeared 2 hours after ingestion. Two victims became unconscious within 5 hours of exposure. On the way to the hospital, two of the eight patients died and the remaining six were admitted. Three of the six patients died in the hospital and three recovered.
4.2. Poisoning due to small gastropods
In China and Taiwan, food poisoning incidents attributable to the ingestion of small gastropods such as Niotha clathrata, Natica vitellus, Oliva miniacea, O. hirasei, Nassarius glans, and Zeuxis samiplicutus, occur frequently [108–112]. In China, more than 300 people were poisoned, and 16 died during 1977–2001. In June 2001, a poisoning incident due to ingestion of a small gastropod of the family Nassariidae, Z. samiplicutus occurred in South Zheijiang, Mainland China. About 30 victims were reported to have symptoms, including paralysis, coma, nausea, vomiting, ataxia, and aphasia. The causative agent was identified as TTX by thin-layer chromatography, electrophoresis, and high performance liquid chromatography [113,114].
It is astonishing that there continues to be many victims, as well as deaths, due to the ingestion of commercially cooked small gastropods in China, which may become a serious public health issue. Moreover, this type of poisoning has spread from the coast of Zheijiang to other areas of the East China Sea and Yellow Sea. Very recently (July 2007) in Nagasaki, Japan, a food poisoning incident occurred due to the same type of small gastropod, Alectrion glans. An investigation revealed that a large amount of TTX (600 MU/g) remained in the leftover cooked gastropods, including muscles and digestive glands (unpublished data). Although the TTX accumulation mechanism in the gastropods remains uncertain, we suspect that the gastropods might become toxic by feeding on the toxic viscera of pufferfish that died after spawning.
4.3. Poisoning due to other TTX-bearing animals
In December 1979, a 41-year-old man ate a boiled digestive gland (about 60 g) of the trumpet shell Charonia sauliae, which he had caught in the coastal waters of Shimizu, Shizuoka Prefecture. Thirty minutes after ingestion, he was hospitalized and received artificial respiration. Although he remained unconscious for 2 days, he recovered fully in 5 days.
The leftover digestive gland had a toxicity score of 17000 MU, and the responsible toxin was identified as TTX [47]. Thus, it was estimated that the victim might have ingested an amount of TTX almost equivalent to the MLD. TTX was also identified as the causative agent in the leftovers from similar TTX poisoning cases in Wakayama Prefecture in 1982 and Miyazaki Prefecture in 1987 [115].
Poisonings due to the ingestion of eggs of the horseshoe crab Carcinoscorpius rotundicauda have occasionally been reported in Thailand [116]. The symptoms of the victims were mostly similar to those caused by TTX or PSP. C. rotundicauda toxin consists mainly of TTX and anhydroTTX [59,117,118], but sometimes contains STX and neoSTX [119]. Hence, the responsible toxin for horseshoe crab poisonings is considered to be TTX and/or PSP.
5. Conclusion
As described above, a large variety of organisms besides pufferfish possess TTX. By knowing the inter- or intra-tissue distribution of TTX, it is possible to estimate the functions of TTX in these organisms. For example, marine pufferfish (Table 1) and flatworms [9,120] retain high toxicity in their eggs, and pufferfish and newts are equipped with TTX-secreting glands or cells in their skin [38–40,121], suggesting that TTX is present as a defensive substance to protect their eggs or themselves from external enemies. In this context, we recently found that the immunity of cultured pufferfish is significantly activated when they are reared with TTX-containing diets [122], though the mechanism remains to be clarified. On the other hand, blue-ringed octopus and ribbon worms possess TTX in the post salivary gland and proboscis, respectively [54,120], indicating that they utilize TTX to capture their prey.
The TTX accumulation mechanism in pufferfish can be explained by bioconcentration via the food chain starting from bacteria. It remains to be elucidated how TTX molecules are absorbed, transferred, and retained/eliminated after entering into the pufferfish body via the toxic food organisms. We recently found that when intramuscularly administered to the non-toxic cultured specimens of TTX-bearing marine pufferfish, TTX is rapidly transferred to the skin and liver through the blood (unpublished data). A similar transfer is also observed when PSP is administered to the non-toxic cultured specimens of PSP-bearing freshwater pufferfish [32]. On the other hand, Nagashima et al. [123,124] reported that liver tissue slices of pufferfish accumulate considerable amounts of TTX in vitro when incubated with TTX-containing medium. These phenomena were not observed in non-toxic fish, suggesting the presence of a specific mechanism in the pufferfish liver or skin whereby the fish selectively and/or actively accumulate TTX/PSP. TTX-binding proteins exist in the blood plasma of pufferfish [125,126], which may be implicated in the transportation of TTX. In this connection, Lee et al. [127] identified novel genes that are possibly related to TTX accumulation in pufferfish.
Not only pufferfish, but various other animals induce TTX intoxication in humans. The symptoms in the patients appear shortly after ingestion of the causative food, and the time to fatality is also very short (average 6 hours). There are currently no antidotes or antitoxins to TTX, and there is no treatment for the seriously TTX-intoxicated patient except for artificial respiration. A monoclonal anti-TTX antibody was recently developed [128], but it can be used only as a reagent for research, and is not expected to have clinical efficacy. There is a possibility that TTX poisoning may increase or diversify further in the future due to changes in the marine environment and the increase in international commerce of fishery products.
TTX has a very important role in neurophysiology, and in other fields of study as a research reagent (a specific sodium channel blocker). In China, it is used as a clinical medicine (an analgesic for terminal cancer patients). In Japan, TTX was previously clinically applied as an analgesic for neuralgia and rheumatism. A thorough understanding of the biochemical properties and mechanisms of TTX accumulation will facilitate the development of technology to use TTX or TTX-bearing organisms effectively as reagents and medicines, or to produce non-toxic cultured pufferfish whose liver can be used as a poison-free food.
References
- Yokoo, A. Chemical studies on pufferfish toxin (3) – separation of spheroidine. Nippon Kagaku Zasshi 1950, 71, 590–592. [Google Scholar]
- Tsuda, K; Ikuma, S; Kawamura, M; Tachikawa, R; Sakai, K; Tamura, C; Amakasu, D. Tetrodotoxin. VII. On the structures of tetrodotoxin and its derivatives. Chem Pharm Bull 1964, 12, 1357–1374. [Google Scholar]
- Woodward, RB. The structure of tetrodotoxin. Pure Appl Chem 1964, 9, 49–74. [Google Scholar]
- Goto, T; Takahashi, S; Kishi, Y; Hirata, Y. Tetrodotoxin. Tetrahedron 1965, 21, 2059–2088. [Google Scholar]
- Narahashi, T. Pharmacology of tetrodotoxin. J Toxicol-Toxin Rev 2001, 20, 67–84. [Google Scholar]
- Noguchi, T; Ebesu, JSM. Puffer poisoning: epidemiology and treatment. J Toxicol-Toxin Rev 2001, 20, 1–10. [Google Scholar]
- Yotsu-Yamashita, M. Chemistry of puffer fish toxin. J Toxicol-Toxin Rev 2001, 20, 51–66. [Google Scholar]
- Jang, JH; Yotsu-Yamashita, M. 6,11-Dideoxytetrodotoxin from the puffer fish, Fugu pardalis. Toxicon 2007, 50, 947–951. [Google Scholar]
- Miyazawa, K; Noguchi, T. Distribution and origin of tetrodotoxin. J Toxicol-Toxin Rev 2001, 20, 11–33. [Google Scholar]
- Noguchi, T; Narita, H; Maruyama, J; Hashimoto, K. Tetrodotoxin in the starfish Astropecten polyacanthus, in association with toxification of a trumpet shell, “boshubora”, Charonia sauliae. Bull Japan Soc Sci Fish 1982, 48, 1173–1177. [Google Scholar]
- Noguchi, T; Jeon, JK; Arakawa, O; Sugita, H; Deguchi, Y; Shida, Y; Hashimoto, K. Occurrence of tetrodotoxin in Vibrio sp. isolated from intestines of xanthid crab, Atergatis floridus. J Biochem 1986, 99, 311–314. [Google Scholar]
- Matsui, T; Sato, H; Hamada, S; Shimizu, C. Comparison of toxicity of the cultured and wild puffer fish Fugu niphobles. Bull Japan Soc Sci Fish 1982, 48, 253. [Google Scholar]
- Saito, T; Maruyama, J; Kanoh, S; Jeon, JK; Noguchi, T; Harada, T; Murata, O; Hashimoto, K. Toxicity of the cultured pufferfish Fugu rubripes rubripes along with their resistibility against tetrodotoxin. Bull Japan Soc Sci Fish 1984, 50, 1573–1575. [Google Scholar]
- Noguchi, T; Arakawa, O; Takatani, T. Toxicity of pufferfish Takifugu rubripes cultured in netcages at sea or aquaria on land. Comp Biochem Physiol, Part D 2006, 1, 153–157. [Google Scholar]
- Matsui, T; Hamada, S; Konosu, S. Difference in accumulation of puffer fish toxin and crystalline tetrodotoxin in the puffer fish, Fugu rubripes rubripes. Bull Japan Soc Sci Fish 1981, 47, 535–537. [Google Scholar]
- Yamamori, K; Kono, M; Furukawa, K; Matsui, T. The toxification of juvenile cultured kusafugu Takifugu niphobles by oral administration of crystalline tetrodotoxin. J Food Fyg Soc Japan 2004, 45, 73–75. [Google Scholar]
- Honda, S; Arakawa, O; Takatani, T; Tachibana, K; Yagi, M; Tanigawa, A; Noguchi, T. Toxification of cultured puffer fish Takifugu rubripes by feeding on tetrodotoxin-containing diet. Nippon Suisan Gakkaishi 2005, 71, 815–820. [Google Scholar]
- Tani, I. Studies on Japanese Pufffish in Association with Poisoning due to Ingestion of Them; Teikoku Tosho: Tokyo, Japan, 1945; p. 103. [Google Scholar]
- Environmental Health Bureau, Ministry of Health and Welfare, Pufferfishes Available in Japan-An Illustrated Guide to Their Identification; Chuo-Hokishuppan: Tokyo, Japan, 1984; p. 79.
- Kanoh, S. Hashimoto, K, Ed.; Distribution of tetrodotoxin in vertebrates. In Recent Advances in Tetrodotoxin Research; Koseisha-Koseikaku: Tokyo, Japan, 1988; pp. 32–44. [Google Scholar]
- Fuchi, Y; Narimatsu, H; Nakama, S; Kotobuki, H; Hirakawa, H; Torishima, Y; Noguchi, T; Ohtomo, N. Tissue distribution of toxicity in a pufferfish, Arothron firmamentum (“hoshifugu”). J Food Hyg Soc Japan 1991, 32, 520–524. [Google Scholar]
- Khora, SS; Isa, J; Yasumoto, T. Toxicity of puffers from Okinawa, Japan. Nippon Suisan Gakkaishi 1991, 57, 163–167. [Google Scholar]
- Hwang, DF; Kao, CY; Yang, HC; Jeng, SS; Noguchi, T; Hashimoto, K. Toxicity of puffer in Taiwan. Nippon Suisan Gakkaishi 1992, 58, 1541–1547. [Google Scholar]
- Boylan, DB; Scheuer, PJ. Pahutoxin: a fish poison. Science 1967, 155, 52. [Google Scholar]
- Thomson, DA. Freudenthal, HD, Ed.; Drugs from the Sea; Marine Technology Society: Washington, D.C., USA, 1968. [Google Scholar]
- Taniyama, S. Toxicity and toxin profile of marine boxfish. In Studies on Parrotfish Poisoning and Similar Incidents Caused by Other Fishes; Ph.D. Thesis, Nagasaki University: Nagasaki, Japan, 2002; pp. 73–80. [Google Scholar]
- Mahmud, Y; Yamamori, K; Noguchi, T. Occurrence of TTX in a brackishwater puffer “midorifugu” Tetraodon nigroviridis, collected from Thailand. J Food Hyg Soc Japan 1999, 40, 363–367. [Google Scholar]
- Mahmud, Y; Yamamori, K; Noguchi, T. Toxicity and tetrodotoxin as the toxic principle of a brackishwater puffer Tetraodon steindachneri, collected from Thailand. J Food Hyg Soc Japan 1999, 40, 391–395. [Google Scholar]
- Kungsuwan, A; Arakawa, O; Promdet, M; Onoue, Y. Occurrence of paralytic shellfish poisons in Thai freshwater puffers. Toxicon 1997, 35, 1341–1346. [Google Scholar]
- Sato, S; Kodama, M; Ogata, T; Saitanu, K; Furuya, M; Hirayama, K; Kakinuma, K. Saxitoxin as toxic principle of a freshwater puffer, Tetraodon fangi, in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar]
- Oliveira, JS; Fernandes, SCR; Schwartz, CA; Bloch, C, Jr; Melo, JAT; Pires, OR, Jr; Freitas, JC. Toxicity and toxin identification in Colomesus asellus, an Amazonian (Brazil) freshwater puffer fish. Toxicon 2006, 48, 55–63. [Google Scholar]
- Ngy, L; Tada, K; Yu, CF; Takatani, T; Arakawa, O. Occurrence of paralytic shellfish toxins in Cambodian Mekong pufferfish Tetraodon turgidus: Selective toxin accumulation in the skin. Toxicon 2008, 51, 280–288. [Google Scholar]
- Sato, S; Ogata, T; Borja, V; Gonzales, C; Fukuyo, Y; Kodama, M. Frequent occurrence of paralytic shellfish poisoning toxins as dominant toxins in marine puffer from tropical water. Toxicon 2000, 38, 1101–1109. [Google Scholar]
- Nakashima, K; Arakawa, O; Taniyama, S; Nonaka, M; Takatani, T; Yamamori, K; Fuchi, Y; Noguchi, T. Occurrence of saxitoxins as a major toxin in the ovary of a marine puffer Arothron firmamentum. Toxicon 2004, 43, 207–212. [Google Scholar]
- Taniyama, S; Mahmud, Y; Tanu, MB; Takatani, T; Arakawa, O; Noguchi, T. Delayed haemolytic activity by the freshwater puffer Tetraodon sp. toxin. Toxicon 2001, 39, 725–727. [Google Scholar]
- Mahmud, Y; Tanu, MB; Takatani, T; Asayama, E; Arakawa, O; Noguchi, T. Chelonodon patoca, a highly toxic marine puffer in Japan. J Nat Toxins 2001, 10, 69–74. [Google Scholar]
- Zaman, L; Arakawa, O; Shimosu, A; Onoue, Y. Occurrence of paralytic shellfish poison in Bangladeshi freshwater puffers. Toxicon 1997, 35, 423–431. [Google Scholar]
- Tanu, MB; Mahmud, Y; Takatani, T; Kawatsu, K; Hamano, Y; Arakawa, O; Noguchi, T. Localization of tetrodotoxin in the skin of a brackishwater puffer Tetraodon steindachneri on the basis of immunohistological study. Toxicon 2002, 40, 103–106. [Google Scholar]
- Mahmud, Y; Okada, K; Takatani, T; Kawatsu, K; Hamano, Y; Arakawa, O; Noguchi, T. Intra-tissue distribution of tetrodotoxin in two marine puffers Takifugu vermicularis and Chelonodon patoca. Toxicon 2003, 41, 13–18. [Google Scholar]
- Mahmud, Y; Arakawa, O; Ichinose, A; Tanu, MB; Takatani, T; Tsuruda, K; Kawatsu, K; Hamano, Y; Noguchi, T. Intracellular visualization of tetrodotoxin (TTX) in the skin of a puffer Tetraodon nigroviridis by immunoenzymatic technique. Toxicon 2003, 41, 605–611. [Google Scholar]
- Kodama, M; Ogata, T; Sato, S. External secretion of tetrodotoxin from puffer fishes stimulated by electric shock. Marine Biol 1985, 87, 199–202. [Google Scholar]
- Saito, T; Noguchi, T; Harada, T; Murata, O; Hashimoto, K. Tetrodotoxin as a biological defense agent for puffers. Bull Japan Soc Sci Fish 1985, 51, 1175–1180. [Google Scholar]
- Mosher, HS; Fuhrman, GJ; Fuhrman, FA; Fischer, HG. Tarichatoxin-tetrodotoxin, a potent neurotoxin. Science 1965, 144, 1100–1110. [Google Scholar]
- Miyazawa, K; Jeon, JK; Maruyama, J; Noguchi, T; Ito, K; Hashimot, K. Occurrence of tetrodotoxin in the flatworm Planocera multitentaculata. Toxicon 1986, 24, 645–650. [Google Scholar]
- Miyazawa, K; Higashiyama, M; Ito, K; Noguchi, T; Arakawa, O; Shida, Y; Hashimoto, K. Tetrodotoxin in two species of ribbon worm (Nemertini), Lineus fuscoviridis and Tubulanus punctatus. Toxicon 1988, 26, 867–874. [Google Scholar]
- Ali, AE; Arakawa, O; Noguchi, T; Miyazawa, K; Shida, Y; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar]
- Narita, H; Noguchi, T; Maruyama, J; Ueda, Y; Hashimoto, K; Watanabe, Y; Hida, K. Occurrence of tetrodotoxin in a trumpet shell, “boshubora” Charonia sauliae. Bull Japan Soc Sci Fish 1981, 47, 935–941. [Google Scholar]
- Noguchi, T; Maruyama, J; Ueda, Y; Hashimoto, K; Harada, T. Occurrence of tetrodotoxin in the Japanese ivory shell Babylonia japonica. Bull Japan Soc Sci Fish 1981, 47, 901–913. [Google Scholar]
- Noguchi, T; Maruyama, J; Narita, H; Hashimoto, K. Occurrence of tetrodotoxin in the gastropod mollusk Tutufa lissostoma (frog shell). Toxicon 1984, 22, 219–226. [Google Scholar]
- Narita, H; Noguchi, T; Maruyama, J; Hashimoto, K. Occurrence of a tetrodotoxin-associated substance in a gastropod, “hanamushirogai” Zeuxis siquijorensis. Bull Japan Soc Sci Fish 1984, 50, 85–88. [Google Scholar]
- Jeon, JK; Narita, H; Nara, M; Noguchi, T; Maruyama, J; Hashimoto, K. Occurrence of tetrodotoxin in a gastropod mollusk, “araregai” Niotha clathrata. Bull Japan Soc Sci Fish 1984, 50, 2099–2102. [Google Scholar]
- Hwang, DF; Chueh, CH; Jeng, SS. Occurrence of tetrodotoxin in the gastropod mollusk Natica lineata (lined moon shell). Toxicon 1990, 28, 21–27. [Google Scholar]
- Narita, H. Tetrodotoxin, with special reference to its origin and the mechanism of toxification. Seikatsu Eisei 1991, 35, 2–15. [Google Scholar]
- Sheumack, DD; Howden, MEH. Maculotoxin: a neurotoxin from the venom glands of the octopus Hapalochlaena maculosa identified as tetrodotoxin. Science 1978, 199, 188–189. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D; Flachsenberger, W. Distribution of tetrodotoxin in the body of the blue-ringed octopus (Hapalochlaena maculosa). Toxicon 2007, 49, 410–412. [Google Scholar]
- Yasumoto, T; Yotsu, M; Endo, A; Murata, M; Kao, CY. Interspecies distribution and biogenetic origin of tetrodotoxin and its derivatives. Pure Appl Chem 1989, 61, 505–508. [Google Scholar]
- Noguchi, T; Uzu, A; Koyama, K; Hashimoto, K. Occurrence of tetrodotoxin as the major toxin in xanthid crab Atergatis floridus. Bull Japan Soc Sci Fish 1983, 49, 1887–1892. [Google Scholar]
- Yasumura, D; Oshima, Y; Yasumoto, T; Alcala, AC; Alcala, LC. Tetrodotoxin and paralytic shellfish toxins in Philippine crabs. Agric Biol Chem 1986, 50, 593–598. [Google Scholar]
- Kungsuwan, A; Nagashima, Y; Noguchi, T; Shida, Y; Suvapeepan, S; Suwansakornkul, P; Hashimoto, K. Tetrodotoxin in the horseshoe crab Carcinoscorpius rotundicauda inhabiting Thailand. Nippon Suisan Gakkaishi 1987, 53, 261–266. [Google Scholar]
- Thuesen, EV; Kogure, K; Hashimoto, K; Nemoto, T. Poison arrowworms: a tetrodotoxin venom in the marine phylum Chaetonnatha. J Exp Mar Biol Ecol 1988, 116, 249–256. [Google Scholar]
- Maruyama, J; Noguchi, T; Jeon, JK; Harada, T; Hashimoto, K. Occurrence of tetrodotoxin in the starfish Astropecten latespinosus. Experientia 1984, 40, 1395–1396. [Google Scholar]
- Maruyama, J; Noguchi, T; Narita, H; Jeon, JK; Otsuka, M; Hashimoto, K. Occurrence of tetrodotoxin in a starfish, Astropecten scoparius. Agric Biol Chem 1985, 49, 3069–3070. [Google Scholar]
- Noguchi, T; Hashimoto, Y. Isolation of tetrodotoxin from a goby Gobius criniger. Toxicon 1973, 11, 305–307. [Google Scholar]
- Yotsu, M; Iorizzi, M; Yasumoto, T. Distribution of tetrodotoxin, 6-epitetrodotoxin, and 11-deoxytetrodotoxin in newts. Toxicon 1990, 28, 238–241. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D. The levels of tetrodotoxin and its analogue 6-epitetrodotoxin in the red-spotted newt Notophthalmus viridescens. Toxicon 2001, 39, 1261–1263. [Google Scholar]
- Yasumoto, T; Yotsu, M; Murata, M. New tetrodotoxin analogues from the newt Cynops ensicauda. J Am Chem Soc 1988, 110, 2344–2345. [Google Scholar]
- Yotsu-Yamashita, M; Mebs, D; Kwet, A; Schneider, M. Tetrodotoxin and its analogue 6-epitetrodotoxin in newts (Triturus spp.; Urodela, Salamandridae) from southern Germany. Toxicon 2007, 50, 306–309. [Google Scholar]
- Kim, YH; Brown, GB; Mosher, HS. Tetrodotoxin: occurrence in atelopid frogs of Costa Rica. Science 1975, 189, 151–152. [Google Scholar]
- Daly, JW; Gusovsky, F; Myers, CW; Yotsu-Yamashita, M; Yasumoto, T. First occurrence of tetrodotoxin in a dendrobatid frog (Colostethus inguinalis), with further reports for the bufonid genus Atelopus. Toxicon 1994, 32, 279–285. [Google Scholar]
- Tanu, MB; Mahmud, Y; Tsuruda, K; Arakawa, O; Noguchi, T. Occurrence of tetrodotoxin in the skin of a rhacophoridid frog Polypedates sp. from Bangladesh. Toxicon 2001, 39, 937–941. [Google Scholar]
- Pires, OR, Jr; Sebben, A; Schwartz, EF; Largura, SWR; Bloch, C, Jr; Morales, RAV; Schwartz, CA. Occurrence of tetrodotoxin (TTX) and its analogues in the Brazilian frog Brachycephalus ephippium (Anura: Brachycephalidae). Toxicon 2002, 40, 761–766. [Google Scholar]
- Pires, OR, Jr; Sebben, A; Schwartz, EF; Morales, RAV; Bloch, C, Jr; Schwartz, CA. Further report of the occurrence of tetrodotoxin and new analogues in the Anuran family Brachycephalidae. Toxicon 2005, 45, 73–79. [Google Scholar]
- Narita, H; Matsubara, S; Miwa, N; Akahane, S; Murakami, M; Goto, T; Nara, M; Noguchi, T; Saito, T; Shida, Y; Hashimoto, K. Vibrio alginolyticus, a TTX-producing bacterium isolated from the starfish Astropecten polyacanthus. Bull Japan Soc Sci Fish 1987, 47, 935–941. [Google Scholar]
- Hashimoto, K; Noguchi, T; Watabe, S. Pohland, AE, Ed.; New aspect of tetrodotoxin. In Microbial Toxins in Foods and Feeds; Plenum Press: New York, 1990; pp. 159–172. [Google Scholar]
- Suenaga, K; Kotoku, S. Detection of tetrodotoxin in autopsy material by gas chromatography. Arch Toxicol 1980, 44, 291–297. [Google Scholar]
- Yasumoto, T; Yasumura, D; Yotsu, M; Michishita, T; Endo, A; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric Biol Chem 1986, 50, 793–795. [Google Scholar]
- Matsui, T; Taketsugu, S; Sato, H; Yamamori, K; Kodama, K; Ishi, A; Hirose, H; Shimizu, C. Toxification of cultured puffer fish by the administration of tetrodotoxin producing bacteria. Nippon Suisan Gakkaishi 1990, 56, 705. [Google Scholar]
- Simidu, U; Noguchi, T; Hwang, DF; Shida, Y; Hashimoto, K. Marine bacteria which produce tetrodotoxin. Appl Environ Microbiol 1987, 53, 1714–1715. [Google Scholar]
- Noguchi, T. Hashimoto, K, Ed.; Food chain-associated toxification of tetrodotoxin-bearing animals. In Recent Advances in Tetrodotoxin Research; Koseisha-Koseikaku: Tokyo, 1988; pp. 85–93. [Google Scholar]
- Shimizu, Y; Kobayashi, M. Apparent lack of tetrodotoxin biosynthesis in captured Taricha torosa and Taricha granulosa. Chem Pharm Bull 1983, 31, 3625–3631. [Google Scholar]
- Hanifin, CT; Brodie, ED, III; Brodie, ED, Jr. Tetrodotoxin levels of the rough-skin newt, Taricha granulosa, increase in long-term captivity. Toxicon 2002, 40, 1149–1153. [Google Scholar]
- Cardall, BL; Brodie, ED, Jr; Brodie, ED, III; Hanifin, CT. Secretion and regeneration of tetrodotoxin in the rough-skin newt Taricha granulosa. Toxicon 2004, 44, 933–938. [Google Scholar]
- Tsuruda, K; Arakawa, O; Noguchi, T. Toxicity and toxin profiles of the newt, Cynops pyrrhogaster from western Japan. J Nat Toxins 2001, 10, 79–89. [Google Scholar]
- Arakawa, K. Resistibility against TTX and PSP. In Studies on the Toxicity of a Japanese Newt Cynops pyrrhogaster; Doctoral thesis, Nagasaki University: Nagasaki, 2001; pp. 50–53. [Google Scholar]
- Daly, JW; Padgett, WL; Saunders, RL; Cover, JF, Jr. Absence of tetrodotoxin in a captive-raised riparian frog, Atelopus varius. Toxicon 1997, 35, 705–709. [Google Scholar]
- Saito, T; Noguchi, T; Harada, T; Murata, O; Abe, T; Hashimoto, K. Resistibility of toxic and nontoxic pufferfish against tetrodotoxin. Bull Japan Soc Sci Fish 1985, 51, 1371. [Google Scholar]
- Koyama, K; Noguchi, T; Uzu, A; Hashimoto, K. Resistibility of toxic and nontoxic crabs against paralytic shellfish poison and tetrodotoxin. Bull Japan Soc Sci Fish 1983, 49, 485–489. [Google Scholar]
- Kaneko, Y; Matsumoto, G; Hanyu, Y. TTX resistivity of Na+ channel in newt retinal neuron. Biochem Biophys Res Commun 1997, 240, 651–656. [Google Scholar]
- Yotsu-Yamashita, M; Nishimori, K; Nitanai, Y; Isemura, M; Sugimoto, A; Yasumoto, T. Binding properties of 3H-PbTx-3 and 3H-saxitoxin to brain membranes and to skeletal muscle membranes of puffer fish Fugu pardalis and the primary structure of a voltage-gated Na+ channel α-subunit (fMNa1) from skeletal muscle of F pardalis. Biochem Biophys Res Commun 2000, 267, 403–412. [Google Scholar]
- Venkatesh, B; Lu, SQ; Dandona, N; See, SL; Benner, S; Soong, TW. Genetic basis of tetrodotoxin resistance in pufferfishes. Current Biol 2005, 15, 2069–2072. [Google Scholar]
- Maruta, S; Yamaoka, K; Yotsu-Yamashita, M. Two critical residues in p-loop regions of puffer fish Na+ channels on TTX-sensitivity. Toxicon 2008, 51, 381–387. [Google Scholar]
- Geffeney, SL; Fujimoto, E; Brodie, ED, III; Brodie, ED, Jr; Ruben, PC. Evolutionary diversification of TTX-resistant sodium channels in a predator-prey interaction. Nature 2005, 434, 759–763. [Google Scholar]
- Bricejl, VM; Connell, L; Konoki, K; MacQuarrie, SP; Scheuer, T; Catterall, WA; Trainer, VL. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434, 763–767. [Google Scholar]
- Shiomi, K; Yamaguchi, S; Kikuchi, T; Yamamori, K; Matsui, T. Occurrence of tetrodotxin-binding high molecular weight substances in the body fluid of shore crab (Hemigrapsus sanguineus). Toxicon 1992, 30, 1529–1537. [Google Scholar]
- Nagashima, Y; Yamamoto, K; Shimakura, K; Shiomi, K. A tetrodotoxin-binding protein in the hemolymph of shore crab Hemigrapsus sanguineus: purification and properties.
- Noguchi, T; Akaeda, H. Pufferfish poisoning. Jpn J Toxicol 1998, 11, 339–345. [Google Scholar]
- Du, SS; Fu, YM; Shih, YC; Chang, PC; Chou, SS; Lue, YH; Hwang, DF. First report on suspected food poisoning with ingestion of dried seasoned fish fillet. J Food Drug Anal 1999, 7, 163–167. [Google Scholar]
- Hsieh, YW; Hwang, PA; Pan, HH; Chen, JB; Hwang, DF. Identification of tetrodotoxin and fish species in an adulterated dried mullet roe implicated in food poisoning. J Food Sci 2003, 68, 142–146. [Google Scholar]
- Hwang, DF; Hwang, PA; Tsai, YH; Lu, YH. Identification of tetrodotoxin and fish species in dried dressed fish fillets implicated in food poisoning. J Food Prot 2002, 65, 389–392. [Google Scholar]
- Hwang, DF; Noguchi, T. Tetrodotoxin poisoning. Adv Food Nutr Res 2007, 52, 142–236. [Google Scholar]
- Hsieh, YW; Shiu, YC; Cheng, CA; Chen, SK; Hwang, DF. Identification of toxin and fish species in cooked fish liver implicated in food poisoning. J Food Sci 2002, 67, 948–952. [Google Scholar]
- Ebesu, JSM; Hokama, Y; Noguchi, T. Hui, YH, Kitts, D, Stanfield, PS, Eds.; Tetrodotoxin. In Foodborne Disease Handbook; Marcel Dekker, Inc.: New York, USA, 2000; Volume 2, pp. 274–276. [Google Scholar]
- Helfrich, P. Fish poisoning in Hawaii. Hawaii Med J 1963, 22, 361. [Google Scholar]
- Sims, JK; Ostman, DC. Emergency diagnosis and management of mild human tetrodotoxin. Ann Emerg Med 1986, 15, 1094. [Google Scholar]
- Mahmud, Y; Tanu, MB; Noguchi, T. First occurrence of a food poisoning incident due to ingestion of Takifugu oblongus, along with a toxicological report on three marine puffer species in Bangladesh. J Food Hyg Soc Japan 1999, 40, 473–480. [Google Scholar]
- Laobhripatr, S; Limpakarnjanarat, K; Sangwonloy, O; Sudhasaneya, S; Anuchatvorakul, B; Leelasitor, S; Saitanu, K. Food poisoning due to consumption of the freshwater puffer Tetraodon fungi in Thailand. Toxicon 1990, 28, 1372–1375. [Google Scholar]
- Mahmud, Y; Arakawa, O; Noguchi, T. An epidemic survey on freshwater puffer poisoning in Bangladesh. J Nat Toxins 2000, 9, 319–326. [Google Scholar]
- Hwang, DF; Cheng, CA; Tsai, YH; Shih, DYC; Ko, HC; Yang, RZ; Jeng, SS. Identification of tetrodotoxin and paralytic shellfish toxins in marine gastropods implicated in food poisoning. Fish Sci 1995, 61, 675–679. [Google Scholar]
- Hwang, DF; Shiu, YC; Hwang, PA; Lu, YH. Tetrodotoxin in gastropods snails implicated in food poisoning in northern Taiwan. J Food Sci 2002, 65, 1341–1344. [Google Scholar]
- Hwang, PA; Tsai, YH; Lu, YH; Hwang, DF. Paralytic toxins in three new gastropod Olividae species implicated in food poisoning in southern Taiwan. Toxicon 2003, 41, 529–533. [Google Scholar]
- Hwang, PA; Tsai, YH; Deng, JF; Cheng, CA; Ho, PH; Hwang, DF. Identification of tetrodotoxin in a marine gastropod Nassarius glans responsible for human morbidity and mortality in Taiwan. J Food Prot 2005, 68, 1696–1701. [Google Scholar]
- Shiu, YC; Lu, YH; Tsai, YH; Hwang, DF. Occurrence of tetrodotoxin in the causative gastropod Polinices didyma and another gastropod Natica lineata collected from western Taiwan. J Food Drug Anal 2003, 11, 159–163. [Google Scholar]
- Sui, LM; Chen, K; Hwang, PA; Hwang, DF. Identification of tetrodotoxin in marine gastropod implicated in food poisoning. J Nat Toxins 2002, 11, 213–220. [Google Scholar]
- Shui, LM; Chen, K; Wang, JY; Mei, HZ; Wang, AZ; Lu, YH; Hwang, DF. Paralytic snail poisoning in Zhoushan: A 25-year retrospective analysis. J Food Prot 2003, 66, 110–114. [Google Scholar]
- Maruyama, J; Noguchi, T; Jeon, JK; Yamazaki, K; Hashimoto, K. Another occurrence of tetrodotoxin in a trumpet shell Charonia sauliae. J Food Hyg Soc Japan 1983, 24, 465–468. [Google Scholar]
- Kanchanapongkul, J; Krittayapoositpot, P. An epidemic of tetrodotoxin poisoning following ingestion of the horseshoe crab Carcinoscorpius rotundicauda. Southeast Asian J Trop Med Public Health 1995, 26, 364–367. [Google Scholar]
- Tanu, MB; Noguchi, T. Tetrodotoxin as a toxin principle in the horseshoe crab Carcinoscorpius rotundicauda collected from Bangladesh. J Food Hyg Soc Japan 1999, 40, 426–431. [Google Scholar]
- Ngy, L; Yu, CF; Takatani, T; Arakawa, O. Toxicity assessment for the horseshoe crab Carcinoscorpius rotundicauda collected from Cambodia. Toxicon 2007, 49, 843–847. [Google Scholar]
- Fusetani, N; Endo, H; Hashimoto, K; Kodama, M. Occurrence and properties of toxins in the horseshoe crab Carcinoscorpius rotundicauda. Toxicon 1983, 21, 165–168. [Google Scholar]
- Tanu, MB; Mahmud, Y; Arakawa, O; Takatani, T; Kajihara, H; Kawatsu, K; Hamano, Y; Asakawa, M; Miyazawa, K; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar]
- Tsuruda, K; Arakawa, O; Kawatsu, K; Hamano, Y; Takatani, T; Noguchi, T. Secretory glands of tetrodotoxin in the skin of a Japanese newt Cynops pyrrhogaster. Toxicon 2002, 40, 131–136. [Google Scholar]
- Honda, S; Ichibu, T; Arakawa, O; Takatani, T; Tachibana, K; Yagi, M; Tanigawa, A; Noguchi, T. Antibody productivity against sheep red blood cells and splenocyte proliferation reaction of tiger puffer (Takifugu rubripes) fed with tetrodotoxin-containing diets. Aquaculture Sci 2005, 53, 205–210. [Google Scholar]
- Nagashima, Y; Toyoda, M; Hasobe, M; Shimakura, K; Shiomi, K. In vitro accumulation of tetrodotoxin in pufferfish liver tissue slices. Toxicon 2003, 41, 569–547. [Google Scholar]
- Matsumoto, T; Nagashima, Y; Kusuhara, H; Sugiyama, Y; Ishizaki, S; Shimakura, K; Shiomi, K. Involvement of carrier-mediated transport system in uptake of tetrodotoxin into liver tissue slices of puffer fish Takifugu rubripes. Toxicon 2007, 50, 173–179. [Google Scholar]
- Matsui, T; Yamamori, K; Furukawa, K; Kono, M. Purification and some properties of a tetrodotoxin binding protein from the blood plasma of kusafugu, Takifugu niphobles. Toxicon 2000, 38, 463–468. [Google Scholar]
- Yotsu-Yamashita, M; Sugimoto, A; Terakawa, T; Shoji, Y; Miyazawa, T; Yasumoto, T. Purification, characterization and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur J Biochem 2001, 268, 5937–5946. [Google Scholar]
- Lee, JH; Kondo, H; Sato, S; Akimoto, S; Saito, T; Kodama, M; Watabe, S. Identification of novel genes related to tetrodotoxin intoxication in pufferfish. Toxicon 2007, 49, 939–953. [Google Scholar]
- Kawatsu, K; Hamano, Y; Yoda, T; Terano, Y; Shibata, T. Rapid and highly sensitive enzyme immunoassay for quantitative determination of tetrodotoxin. Jpn J Med Sci Biol 1997, 50, 133–150. [Google Scholar]
Figure 1.
Structure of TTX.

Figure 2.
HPLC of TTX fraction from a Vibrio strain (left) and of authentic TTX (right) [74].
Figure 2.
HPLC of TTX fraction from a Vibrio strain (left) and of authentic TTX (right) [74].

Figure 3.
TTX infestation to the non-toxic cultured pufferfish T. rubripes and non-toxic three species of fish by feeding TTX-containing livers of wild pufferfish. ▵: Cultured pufferfish; TTX dosage of 0.5 MU/g body mass/day. ○: cultured pufferfish; TTX dosage of 4 MU/g body mass/day. □: non-toxic three species of fish; TTX dosage of 4 MU/g body mass/day.
Figure 3.
TTX infestation to the non-toxic cultured pufferfish T. rubripes and non-toxic three species of fish by feeding TTX-containing livers of wild pufferfish. ▵: Cultured pufferfish; TTX dosage of 0.5 MU/g body mass/day. ○: cultured pufferfish; TTX dosage of 4 MU/g body mass/day. □: non-toxic three species of fish; TTX dosage of 4 MU/g body mass/day.

Figure 4.
Proposed mechanism of TTX accumulation in marine animals.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maximal toxicity* | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Family | Habitat | Species | Ovary | Testis | Liver | Skin | Intestine | Muscle | Blood |
| Tetraodontidae | Marine | Japanese pufferfish | |||||||
| Takifugu niphobles | ● | ○ | ● | ◎ | ● | ○ | — | ||
| T. poecilonotus | ● | ◎ | ● | ◎ | ◎ | ○ | — | ||
| T. pardalis | ● | ○ | ● | ◎ | ◎ | × | × | ||
| T. snyderi | ● | × | ● | ◎ | ◎ | ○ | — | ||
| T. porphyreus | ● | × | ● | ◎ | ◎ | × | — | ||
| T. chinensis | ● | — | ● | — | — | — | — | ||
| T. obscurus | ● | × | ◎ | ◎ | ◎ | × | — | ||
| T. exascurus | ● | × | ◎ | ◎ | — | × | — | ||
| T. pseudommus | ● | × | ○ | ○ | ○ | × | — | ||
| T. chrysops | ◎ | × | ◎ | ◎ | ○ | × | × | ||
| T. vermicularis | ◎ | × | ◎ | ◎ | ○ | × | — | ||
| T. rubripes | ◎ | × | ◎ | × | ○ | × | × | ||
| T. xanthopterus | ◎ | × | ◎ | × | ○ | × | — | ||
| T. stictonotus | ◎ | × | ◎ | ○ | × | × | — | ||
| Tetraodon alboreticulatus | ● | — | ○ | ○ | ◎ | ○ | — | ||
| Pleuranacanthus sceleratus | ● | — | ○ | ○ | ◎ | ○ | — | ||
| Chelonodon patoca** | ◎ | ◎ | ◎ | ● | — | ◎ | — | ||
| Arothron firmamentum | ◎ | × | × | ○ | × | × | — | ||
| Canthigaster rivulata | × | — | ○ | ◎ | ○ | × | — | ||
| Lagocephalus lunaris | × | × | × | ◎ | × | ● | — | ||
| L. inermis | × | × | ◎ | × | × | × | — | ||
| L. wheeleri | × | × | × | × | × | × | — | ||
| L. gloveri | × | × | × | × | × | × | — | ||
| Sphoeroides pachygaster | × | × | × | × | × | × | — | ||
| Marine | Chinese pufferfish | ||||||||
| Takifugu flavidus | ● | ◎ | ● | ◎ | ◎ | ○ | — | ||
| Brackish | Thai pufferfish | ||||||||
| Tetraodon nigroviridis | — | — | × | ◎ | ○ | ○ | — | ||
| T. steindachneri | — | — | × | ◎ | × | × | — | ||
| Diodontidae | Marine | Japanese pufferfish | |||||||
| Diodon holocanthus | × | — | × | × | × | × | — | ||
| Chilomycterus affinis | × | — | × | × | × | × | — | ||
| Ostraciidae | Marine | Japanese pufferfish | |||||||
| Ostracion immaculatum | × | × | × | × | × | × | — | ||
| Lactoria diaphana | × | × | × | × | × | × | — | ||
| Aracana aculeata | × | × | × | × | × | × | — | ||
*×: <10 MU/g tissue; ○: 10–100 MU/g tissue (weakly toxic); ◎: 100–1000 MU/g tissue (moderately toxic); ●: >1000 MU/g tissue (strongly toxic), where 1 MU (mouse unit) is defined as the amount of toxin that kills a male mouse of ddY strain (20 g body weight) in 30 min after intraperitoneal administration. The amount is equivalent to about 0.2 μg of TTX. —: no data available.
**Marine to brackish water species.
Table 2.
Distribution of TTX in animals other than pufferfish [9].
| Animals | Toxic parts | Maximal toxicity* | Ref | |
|---|---|---|---|---|
| Platyhelminthes | ||||
| Turbellaria | ||||
| Flatworms | Planocera spp. | Whole body | ● | [44] |
| Nemertinea | ||||
| Ribbonworms | Lineus fuscoviridis | Whole body | ● | [45] |
| Tubulanus punctatus | Whole body | ◎ | [45] | |
| Cephalothrix linearis | Whole body | ● | [46] | |
| Mollusca | ||||
| Gastropoda | Charonia sauliae | Digestive gland | ● | [47] |
| Babylonia japonica | Digestive gland | ○ | [48] | |
| Tutufa lissostoma | Digestive gland | ◎ | [49] | |
| Zeuxis siquijorensis | Whole body | ● | [50] | |
| Niotha clathrata | Whole body | ● | [51] | |
| Niotha lineata | Whole body | ● | [52] | |
| Cymatium echo | Digestive gland | ○ | [53] | |
| Pugilina ternotoma | Digestive gland | ○ | [53] | |
| Cephalopoda | Hapalochlaena maculosa | Posterior salivary gland (adult) | ● | [54] |
| Whole body (semi-adult) | ○ | [55] | ||
| Annelida: | ||||
| Polychaeta | Pseudopolamilla occelata | Whole body | ○ | [56] |
| Arthropoda: | ||||
| Xanthidae crabs | Atergatis floridus | Whole body | ○ | [57] |
| Zosimus aeneus | Whole body | ○ | [58] | |
| Horseshoe crab | Carcinoscorpius rotundicauda | Egg | ○ | [59] |
| Chaetognatha: | ||||
| Arrowworms | Parasagitta spp. | Head | ▵ | [60] |
| Flaccisagitta spp. | Head | ▵ | [60] | |
| Echinodermata: | ||||
| Starfish | Astropecten spp. | Whole body | ◎ | [61,62] |
| Vertebrata: | ||||
| Pisces | ||||
| Goby | Yongeichthys criniger | Skin, viscera, gonad | ◎ | [63] |
| Amphibia | ||||
| Newts | Taricha spp. | Skin, egg, ovary, muscle, blood | ◎ | [43] |
| Notophthalmus spp. | Skin, egg ovary | ○ | [64,65] | |
| Cynopsis spp. | Skin, egg, ovary, muscle, blood | ○ | [66] | |
| Triturus spp. | Skin, egg, ovary, muscle, blood | ▵ | [64,67] | |
| Frogs | Atelopus spp. | Skin | ● | [68] |
| Colostethus sp. | Skin | ◎ | [69] | |
| Polypedates sp. | Skin | ◎ | [70] | |
| Brachycephalus spp. | Skin, liver | ◎ | [71,72] | |
*Maximal toxicity is shown by the same symbols as in Table 1.
▵derivatives of TTX were detected (toxicity data are unavailable).
Table 3.
Toxicity of liver of the pufferfish T. rubripes cultured in net cages [14]
| Place of culture | Year of collection | Age | Number of collection* | Toxicity (MU/g) |
|---|---|---|---|---|
| Nagasaki** | 2001 | 1–3 | 494 | < 2 |
| 2002 | 1–3 | 1021 | < 2 | |
| 2003 | 2 | 240 | < 2 | |
| Kumamoto | 2001 | 1–2 | 829 | < 2 |
| 2002 | 2 | 587 | < 2 | |
| 2003 | Unknown | 145 | < 2 | |
| Kagoshima | 1981–83 | 1–2 | 47 | < 10 |
| 1990–91 | 1–2 | 40 | < 5 | |
| 2002 | 2 | 46 | < 2 | |
| Ehime | 2001 | 2 | 379 | < 2 |
| 2002 | 2 | 140 | <2 | |
| Fukui | 1982–83 | 1–2 | 25 | < 10 |
| Wakayama | 1983 | 1 | 12 | < 10 |
| 2002 | 2 | 81 | < 2 | |
| Shizuoka | 2003 | 2 | 70 | <2 |
| Unknown | 2001 | 2 | 101 | <2 |
| Total | 4257 | |||
About 2500 each of fish were reared in a net cage (10 x 10 x 4 m) floating on the sea, whose bottom was more than 10 m apart from the sea bottom. After 1–3 years of culture, the toxicity of T. rubripes liver never exceeded 2–10 MU/g (n=4257).
*Specimens were randomly collected from the several numbers of netcages;
**Partly including the specimens of unknown age.
| Place of culture | Year of collection | Age | Number of collection* | Toxicity (MU/g) |
|---|---|---|---|---|
| Yobuko, Saga | 2001 | 2 | 114 | < 2 |
| 2002 | 2 | 228 | < 2 | |
| 2003 | 2 | 358 | < 2 | |
| 2004 | 2 | 349 | < 2 | |
| 2005 | 2 | 100 | < 2 | |
| 2006 | 2 | 250 | < 2 | |
| 2007 | 2 | 266 | < 4–8 | |
| 2 | 60 | < 2–4 | ||
| 1725 | ||||
About 2500 each of fish were reared in an indoor aquarium of 100 ton with the seawater that was pumped up from the sea, filtered through a membrane, and then disinfected by electrolysis. After 2 years of culture, the toxicity of T. rubripes liver never exceeded 2–8 MU/g (n=1725).
*Specimens were randomly collected from the 1–3 aquaria in every winter of 2001–2007.
| Species | MLD (MU/20 g)* | Ref | |
|---|---|---|---|
| TTX-bearing organisms | |||
| Xanthid crab | Atergatis floridus | 1000 | [87] |
| Tropical goby | Yongeichthys criniger | > 300 | [63] |
| Japanese newt | Cynops pyrrhogaster | > 10000 | [84] |
| Pufferfish | [86] | ||
| Toxic | Takifugu niphobles | 700–750 | |
| T. pardalis | 500–550 | ||
| T. rubripes (culture) | 300–500 | ||
| Generally non-toxic or rarely toxic | Lagocephalus wheeleri | 15–18 | |
| L. gloveri | 19–20 | ||
| Liosaccus cutaneous | 13–15 | ||
| Non-toxic | Ostracion immaculatum | 0.9–1.3 | |
| TTX-free vertebrates | [86] | ||
| Teleosts | Oplegnathus punctatus | 0.8–0.9 | |
| O. fasciatus | 0.8–1.8 | ||
| Girella punctata | 0.3–0.5 | ||
| Land mammal | |||
| Mouse | Mus musculus | 1 | |
*MLD of TTX (MU/20 g body mass) that killed 100% of the test animals by intraperitoneal injection.
| Family | Species | Muscle | Edible part Skin | Male gonad |
|---|---|---|---|---|
| Tetraodontidae | “Kusafugu” Takifugu niphobles | ○ | — | — |
| “Komonfugu” T. poecilonotus | ○ | — | — | |
| “Higanfugu” T. pardalis | ○ | — | — | |
| “Shousaifugu” T. snyderi | ○ | — | ○ | |
| “Nashifugu” T. vermicularisa* | ○ | — | — | |
| “Mafugu” T. porphyreus | ○ | — | ○ | |
| “Mefugu” T. obscurus | ○ | — | ○ | |
| “Akamefugu” T. chrysops | ○ | — | ○ | |
| “Torafugu” T. rubripes | ○ | ○ | ○ | |
| “Karasu” T. chinensis | ○ | ○ | ○ | |
| “Shimafugu” T. xanthopterus | ○ | ○ | ○ | |
| “Gomafugu” T. stictonotus | ○ | — | ○ | |
| “Kanafugu” Lagocephalus inermis | ○ | ○ | ○ | |
| “Shirosabafugu” L. wheeleri | ○ | ○ | ○ | |
| “Kurosabafugu” L. gloveri | ○ | ○ | ○ | |
| “Yoritofugu” Sphoeroides pachygaster | ○ | ○ | ○ | |
| “Sansaifugu” T. flavidus | ○ | — | — | |
| Diodontidae | “Ishigakifugu” Chilomycterus reticulatus | ○ | ○ | ○ |
| “Harisenbon” Diodon holocanthus | ○ | ○ | ○ | |
| “Hitozuraharisenbon” D. liturosus | ○ | ○ | ○ | |
| “Nezumifugu” D. hystrix | ○ | ○ | ○ | |
| Ostracidae | “Hakofugu” Ostraction immaculatum | ○ | — | ○ |
○Edible; —, non-edible.
*Applicable to muscle of the species caught in Shimabara Bay, Tachibana Bay and the Inland Sea of Kagawa and Okayama, and to male gonad in Shimabara Bay and Tachibana Bay.
| Year | Number of incidents | Number of patients | Number of deaths | Mortality (%) |
|---|---|---|---|---|
| 1965 | 106 | 152 | 88 | 57.9 |
| 1970 | 46 | 73 | 33 | 45.2 |
| 1975 | 52 | 75 | 30 | 40.0 |
| 1980 | 46 | 90 | 15 | 16.7 |
| 1985 | 30 | 41 | 9 | 22.0 |
| 1990 | 33 | 55 | 1 | 1.8 |
| 1995 | 30 | 42 | 2 | 4.8 |
| 1996 | 21 | 34 | 3 | 8.8 |
| 1997 | 28 | 44 | 6 | 13.6 |
| 1998 | 27 | 39 | 4 | 10.3 |
| 1999 | 20 | 34 | 2 | 5.9 |
| 2000 | 29 | 40 | 0 | 0.0 |
| 2001 | 31 | 52 | 3 | 5.8 |
| 2002 | 32 | 49 | 5 | 10.2 |
| 2003 | 28 | 35 | 3 | 8.6 |
| 2004 | 43 | 58 | 2 | 3.4 |
| 2005 | 40 | 49 | 2 | 4.1 |
| 2006 | 25 | 32 | 1 | 3.1 |
| 2007 | 24 | 38 | 2 | 5.3 |
Share and Cite
MDPI and ACS Style
Noguch, T.; Arakawa, O. Tetrodotoxin – Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication. Mar. Drugs 2008, 6, 220-242. https://doi.org/10.3390/md6020220
AMA Style
Noguch T, Arakawa O. Tetrodotoxin – Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication. Marine Drugs. 2008; 6(2):220-242. https://doi.org/10.3390/md6020220
Chicago/Turabian StyleNoguch, Tamao, and Osamu Arakawa. 2008. "Tetrodotoxin – Distribution and Accumulation in Aquatic Organisms, and Cases of Human Intoxication" Marine Drugs 6, no. 2: 220-242. https://doi.org/10.3390/md6020220