Aqueous Vernomia amygdalina Extracts Alter MCF-7 Cell Membrane Permeability and Efflux

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
Materials and Methods
V. amygdalina Preparation
Cell Culture
Cell Proliferation Studies
DNA Synthesis Assay
Membrane Permeability and Efflux Determination
Statistical Analysis
Results
Time-Dependent Cell Growth
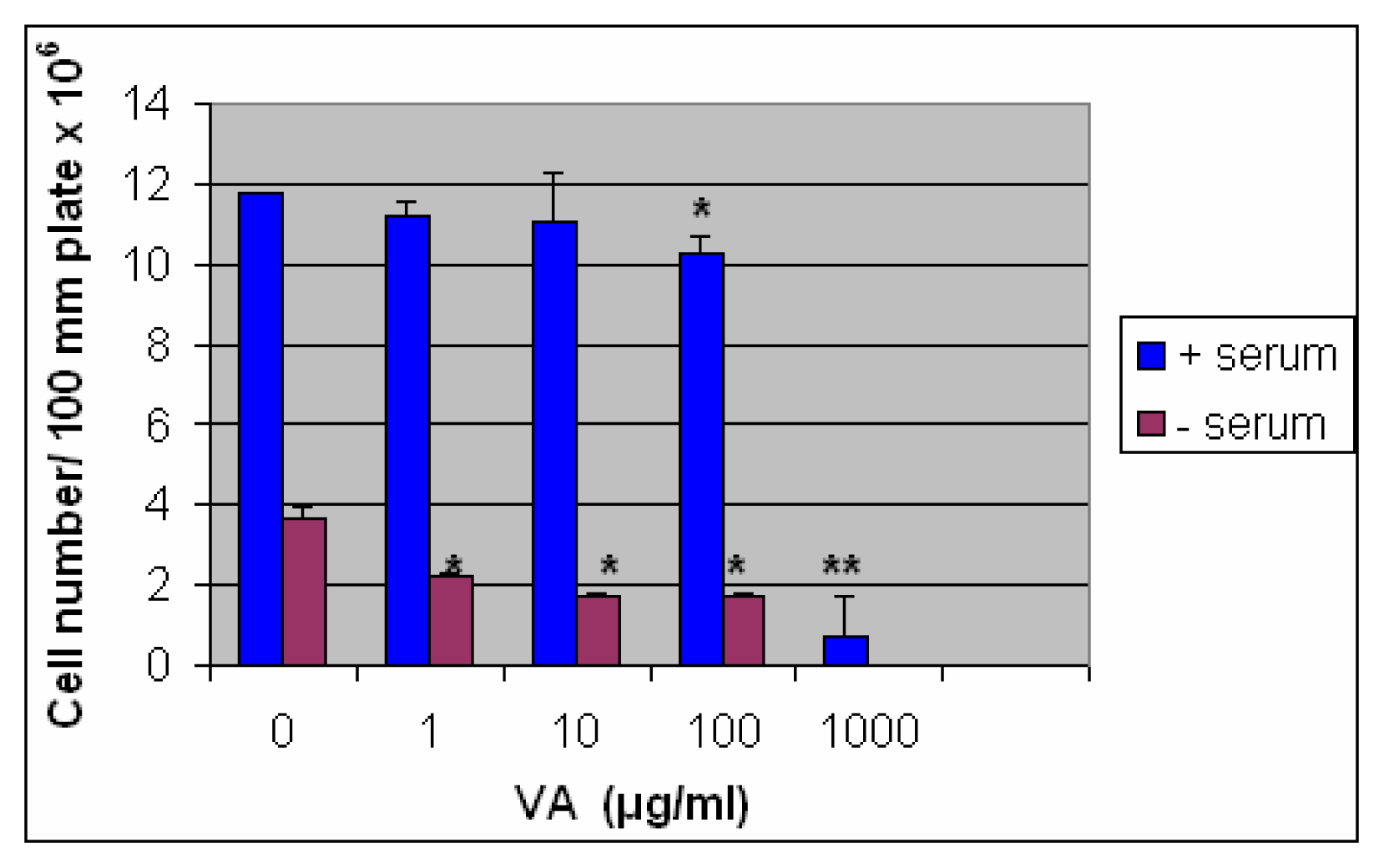
Inhibition of Cell Growth by VA in the Presence and Absence of Serum
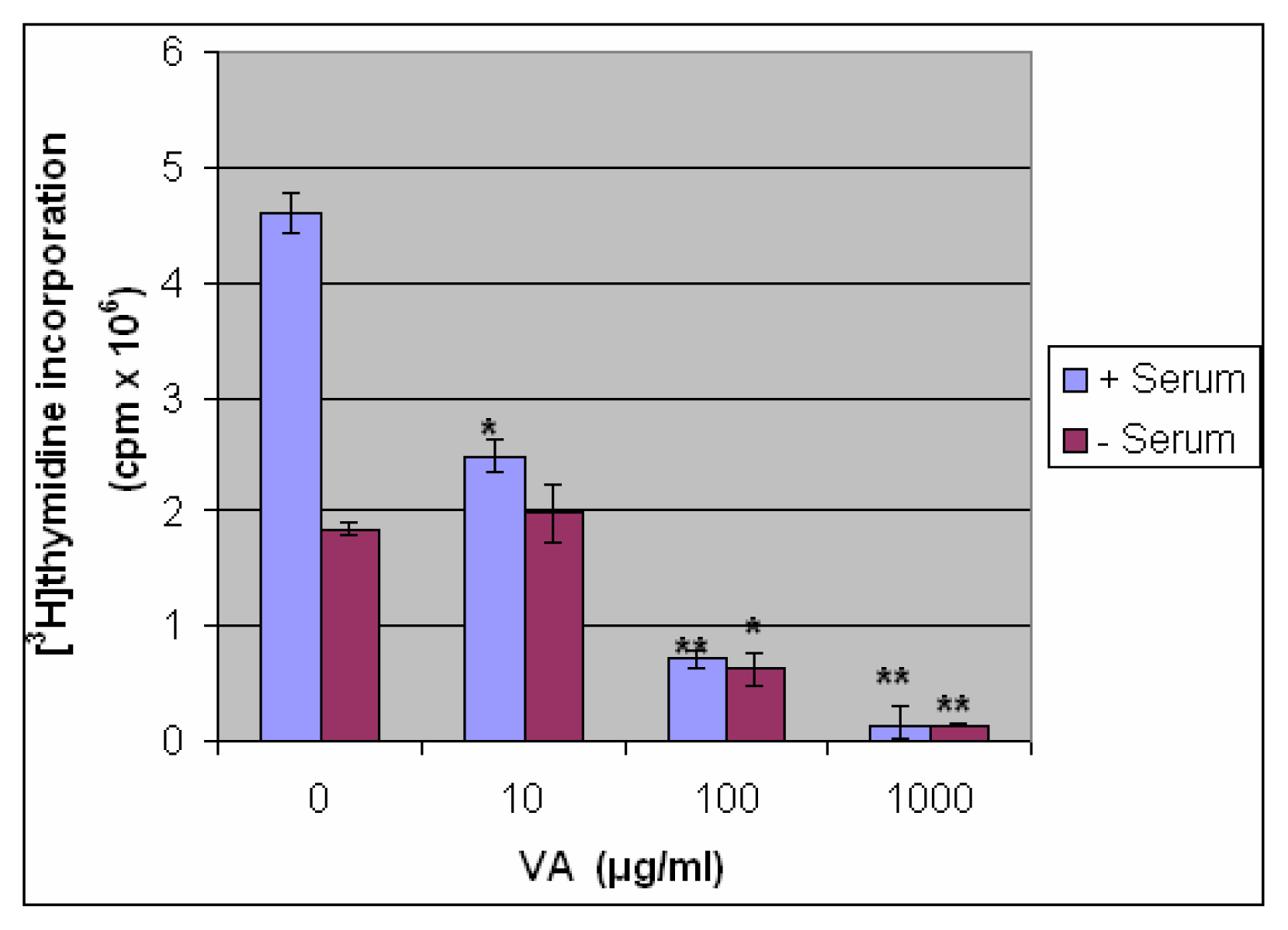
Inhibition of DNA Synthesis by VA in the Presence and Absence of Serum
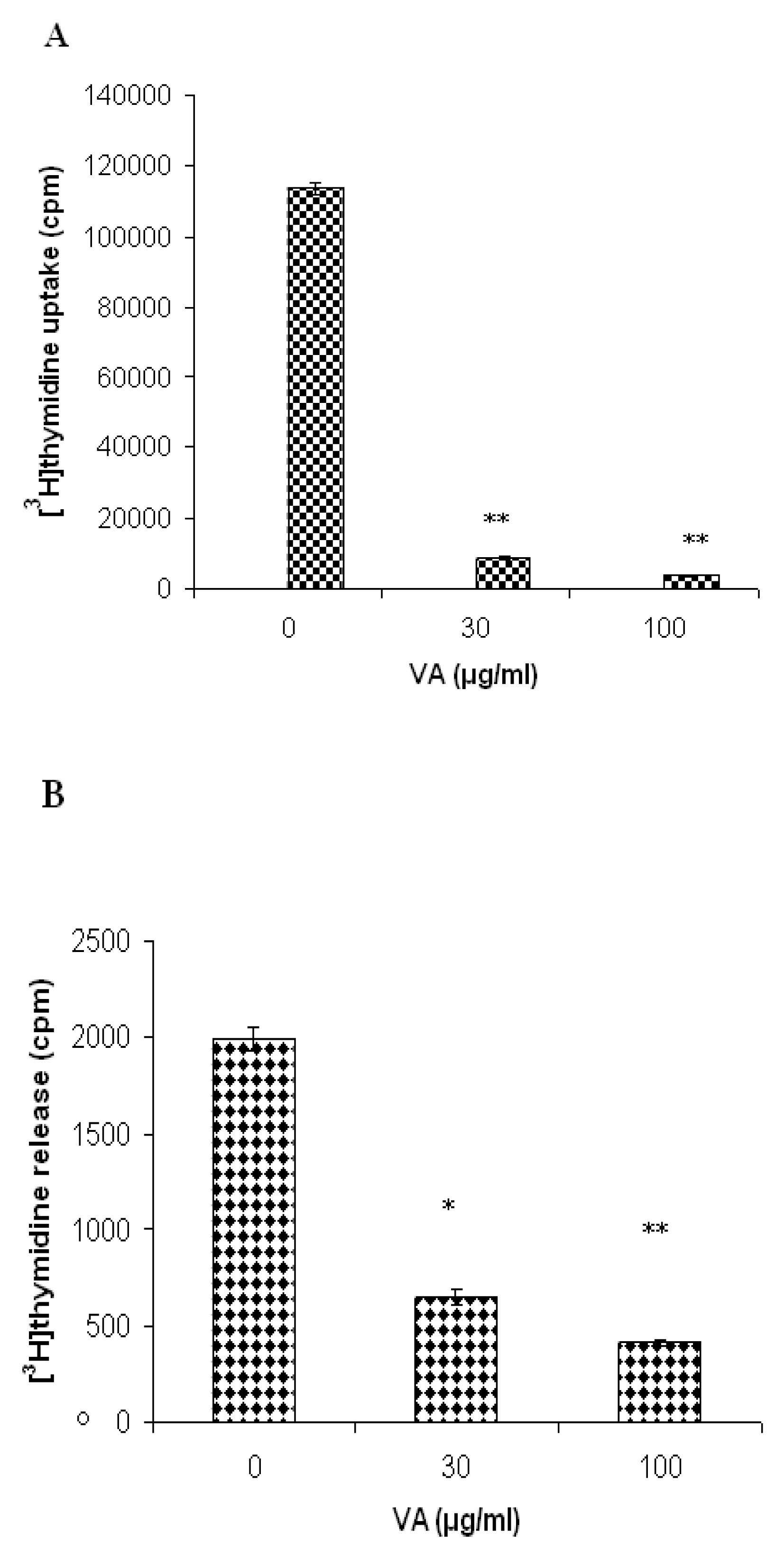
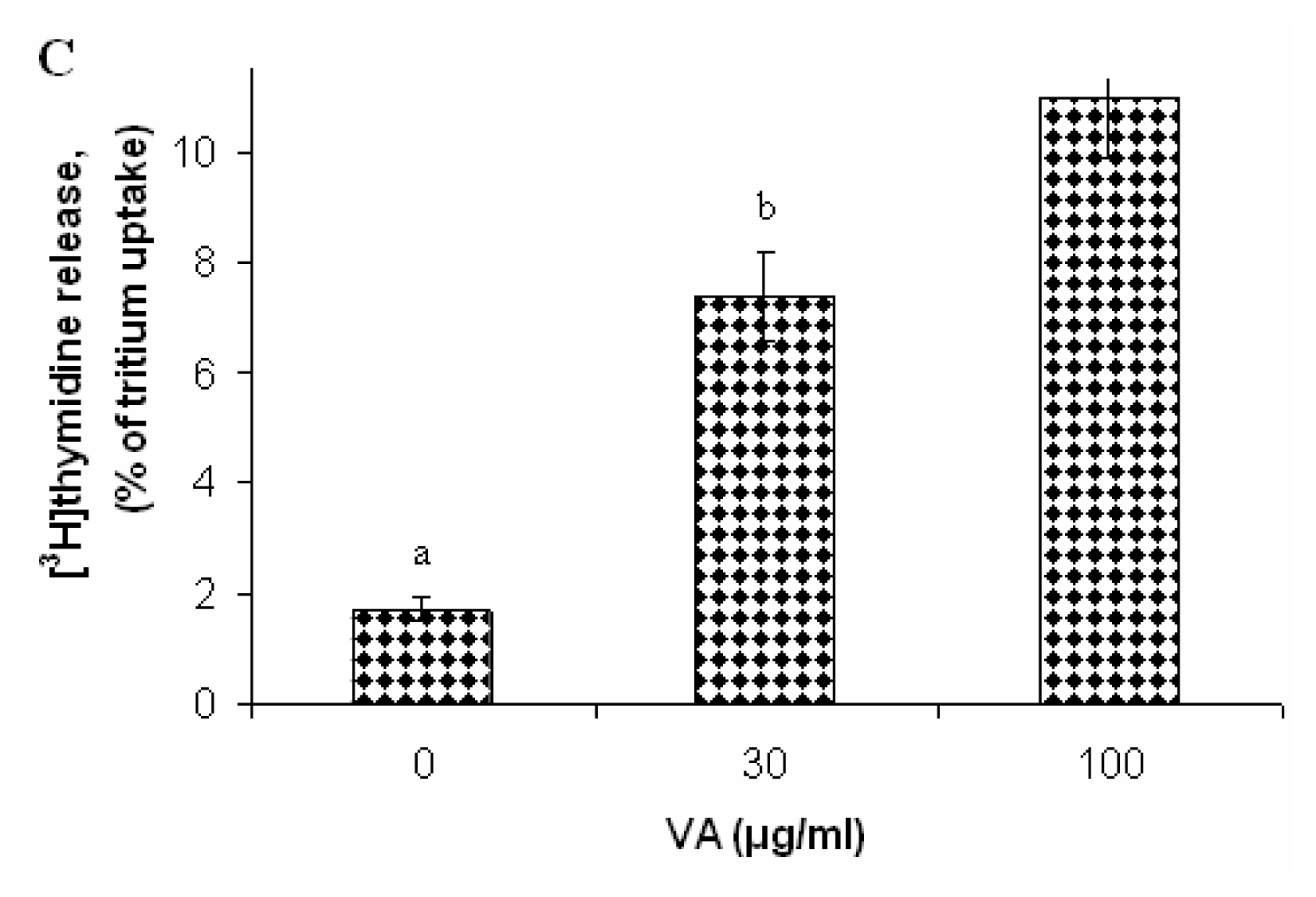
Effects of V. Amygdalina Treatment on Membrane Permeability and Efflux
Discussion





Acknowledgements
References
- American Institute for Cancer Research (AIRC), Food, nutrition and the prevention of cancer: a global perspective (summary); Washington DC, 1997; pp. 9–15.
- Jemal, T.; Thomas, A.; Murray, T.; Thun, M. Cancer Statistics. CA Cancer J. Clin 2002, 52, 23–47. [Google Scholar]
- Richardson, M. A.; Sanders, T.; Palmer, J. L.; Greisinger, A.; Singletary, S. E. Complementary/ Alternative medicine use in a comprehensive cancer center and the implication for oncology. J. Clin. Oncol 2000, 18, 2505–2514. [Google Scholar]
- Zeegers, M.P.; Goldbohm, R. A.; Van den Brant, P. A. Consumption of vegetables and fruits and urothelial cancer incidence: a prospective study. Cancer Epidemiol. Biomarkers Prev 2001, 10, 1121–1128. [Google Scholar]
- Pezzuto, J. M. Plant-derived anticancer agents. Biochem. Pharmacol 1997, 53, 121–133. [Google Scholar]
- Kobayashi, T.; Nakata, T.; Kuzumaki, T. Effect of flavonoids on cell cycle progression in prostate cancer cells. Cancer Lett 2000, 176, 17–23. [Google Scholar]
- Block, G.; Patterson, B.; Subar, A. Fruits, vegetables, and cancer prevention: a review of the epidemiological evidence. Nutr. Cancer 1992, 18, 1–29. [Google Scholar]
- Fahey, J. W.; Zhang, Y.; Talalay, P. Broccoli sprouts: an exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc Natl. Acad. Sci. USA 1997, 94, 10367–10372. [Google Scholar]
- Maoka, T.; Mochida, K.; Kozuka, M.; Ito, Y.; Fujiwara, Y.; Hashimoto, K.; Enjo, F.; Ogata, M.; Nobukuni, Y.; Tokuda, H.; Nishino, H. Cancer chemopreventive activity of carotenoids in the fruits of red paprika capsicum annuum L. Cancer Lett 2001, 172, 103–109. [Google Scholar]
- Terry, P.; Lagergren, J.; Hansen, H.; Wolk, A.; Nyren, O. Fruits and vegetables consumption in the prevention of esophageal and cardiac cancers. Eur. J. Cancer Prev 2001, 10, 365–369. [Google Scholar]
- Rock, C. L.; Moskowitz, A.; Huizar, B.; Saenz, C. C.; Clark, J. T.; Daly, T. L.; Chin, H.; Behling, C.; Ruffin, M. T. 4th. High vegetable and fruit diet intervention in premenopausal women with cervical intraepithelial neoplasia. J. Am. Diet Assoc 2001, 101, 1167–1174. [Google Scholar]
- Simopoulos, A. P. The Mediterranean diets: what is so special about the diet of Greece? The scientific evidence. J. Nutr 2001, 131 11 Suppl, 3065S–3073S. [Google Scholar]
- Singletary, K. Natural products and cancer chemoprevention. J. Nutr 2000, 130, 465–466. [Google Scholar]
- Chung, S. Y.; Sung, M. K.; Kim, N. H.; Jang, J. O. Inhibition of P-glycoprotein by natural products in human breast cancer cells. Arch. Pharm. Res 2005, 28(7), 823–828. [Google Scholar]
- Jakubilova, J.; Bao, Y.; Sedlac, J. Isothiocyanates induce cell cycle arrest, apoptosis and mitochondrial potential depolarization in HL-60 and multidrug-resistant cell lines. Anticancer Res 2005, 25(5), 3375–3386. [Google Scholar]
- Dragsted, L.; Srube, M.; Larsen, J. Cancer-preventive factors in fruits and vegetables: biochemical and biological background. Parmacol. Toxicol 1993, 72 suppl 1, 116–135. [Google Scholar]
- Izevbigie, E. B. Discovery of water-soluble anticancer agents (Edotides) from a vegetable found in Benin City Nigeria. Exp. Biol. Med 2003, 228, 293–298. [Google Scholar]
- Izevbigie, E. B.; Bryant, J. L.; Walker, A. A Novel inhibitor of extracellular signal-regulated kinases and human breast cancer cell growth. Exp. Biol. Med 2004, 229, 163–169. [Google Scholar]
- Jahansson, S.; Gullbo, J.; Lindholm, P.; Ek, B.; Thunberg, E.; Samuelsson, G.; Larsson, R.; Bohlin, L.; Claeson, P. Small, novel proteins from the mistletoe phoradendron tomentosum exhibit highly selective cytotoxicity to human breast cancer cells. Cell Mol. Life Sci 2003, 60, 165–75. [Google Scholar]
- Karkabounas, S.; Assimakopoulos, D.; Malamas, M.; Skaltsounis, A. L.; Leonce, S.; Zelovitis, J.; Stefanou, D.; Evangelou, A. Antiproliferative and anticarcinogenic effects of an aqueous preparation of Abies alba and Viscum album se abies, on a L-1210 malignant cell line and tumor-bearing Wistar rats. Anticancer Res 2000, 20(6B), 4391–4395. [Google Scholar]
- Florack, D. E.; Stickema, W. J. Thionins: properties, possible biological roles and mechanisms of action. Plant Mol. Biol 1994, 26, 25–37. [Google Scholar]
- Froy, O.; Gurevitz, M. Membrane potential modulators: a thread of scarlet from plants to animals. FASEB J 1998, 12, 1793–1796. [Google Scholar]
- Coulon, A.; Berkane, E.; Sautereau, A.M.; Urech, K.; Rouge, P.; Lopez, A. Modes of membrane interaction of a natural cysteine-rich peptides: viscotoxin A3. Biochim. Biophys Acta 2002, 1559(2), 145–159. [Google Scholar]
- Stec, B.; Markman, O.; Rao, U.; Heffron, G.; Henderson, S.; Vernon, L. P.; Brumfeld, V.; Teeter, M.M. Proposal for molecular mechanism of thionins deduced from physico-chemical studies of plant toxins. J. Pept. Res 2004, 64(6), 210–224. [Google Scholar]
- Carrasco, L.; Vazquez, D.; Hernandez-Lucas, C.; Carbonero, P.; Carcia-Olmedo, F. Thionins: plant peptides that modify membrane permeability in cultured mammalian cells. Eur. J. Biochem 1981, 116, 185–189. [Google Scholar]
- Hughes, P.; Dennist, E.; Whitecross, M.; Llewellyn, D.; Gage, P. The cytotoxic plant protein, β-purothionin, forms ion channels in lipid membrane. J. Biol. Chem 2000, 275(2), 823–827. [Google Scholar]
- Garcia-Olmedo, F.; Molina, A.; Alamillo, J.M.; Rodriquez-Palenzuela, P. Plant defense peptides. Biopolymers 1998, 47(6), 479–491. [Google Scholar]
- Taiwo, O.; Xu, H.X.; Lee, S.F. Antibacterial activities of extracts from Nigerian chewing sticks. Phytother. Res 1999, 13(8), 675–679. [Google Scholar]
- Babalola, O. O.; Anetor, J. I.; Adeniyi, F. A. Amelioration of carbon tetrachloride-induced hepatotoxicity by terpenoid extract from leaves of Vernonia amygdalina. African. J. Med. Med. Sci 2001, 30(1–2), 91–93. [Google Scholar]
- Howard, C. B.; Stevens, J.; Izevbigie, E. B.; Walker, A.; McDaniel, O. Time and dose-dependent modulation of phase 1 and phase 2 gene expression in response to treatment of MCF-7 cells with a natural anti-cancer agent. Cell. Mol. Biol 2003, 49(7), 1057–1065. [Google Scholar]
© 2006 MDPI. All rights reserved.
Share and Cite
Opata, M.M.; Izevbigie, E.B. Aqueous Vernomia amygdalina Extracts Alter MCF-7 Cell Membrane Permeability and Efflux. Int. J. Environ. Res. Public Health 2006, 3, 174-179. https://doi.org/10.3390/ijerph2006030019
Opata MM, Izevbigie EB. Aqueous Vernomia amygdalina Extracts Alter MCF-7 Cell Membrane Permeability and Efflux. International Journal of Environmental Research and Public Health. 2006; 3(2):174-179. https://doi.org/10.3390/ijerph2006030019
Chicago/Turabian StyleOpata, Michael M., and Ernest B. Izevbigie. 2006. "Aqueous Vernomia amygdalina Extracts Alter MCF-7 Cell Membrane Permeability and Efflux" International Journal of Environmental Research and Public Health 3, no. 2: 174-179. https://doi.org/10.3390/ijerph2006030019
APA StyleOpata, M. M., & Izevbigie, E. B. (2006). Aqueous Vernomia amygdalina Extracts Alter MCF-7 Cell Membrane Permeability and Efflux. International Journal of Environmental Research and Public Health, 3(2), 174-179. https://doi.org/10.3390/ijerph2006030019