Microalgal Species Selection for Biodiesel Production Based on Fuel Properties Derived from Fatty Acid Profiles

Abstract
:1. Introduction
2. Materials and Methodology
2.1. Materials

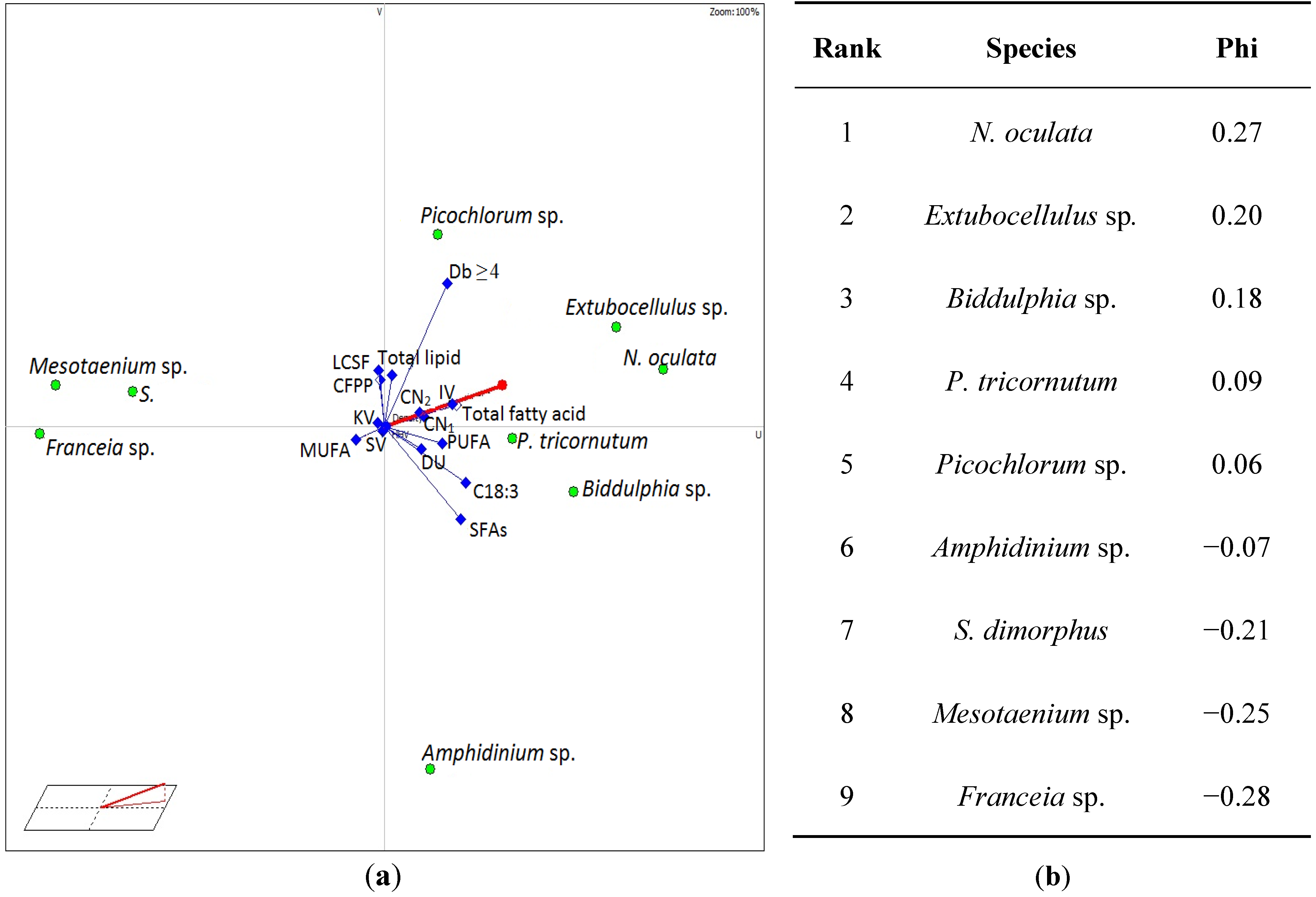
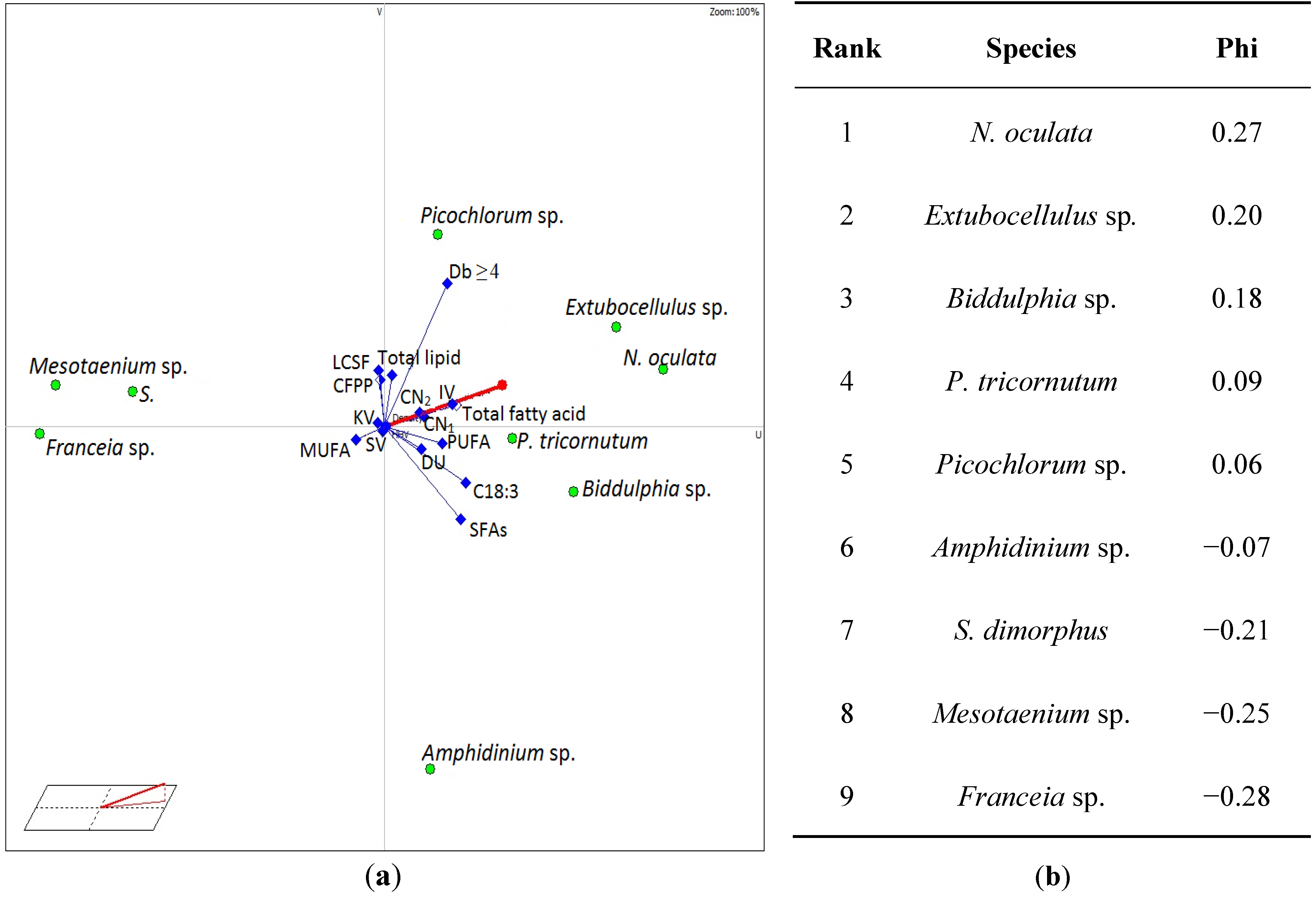{kind=link}
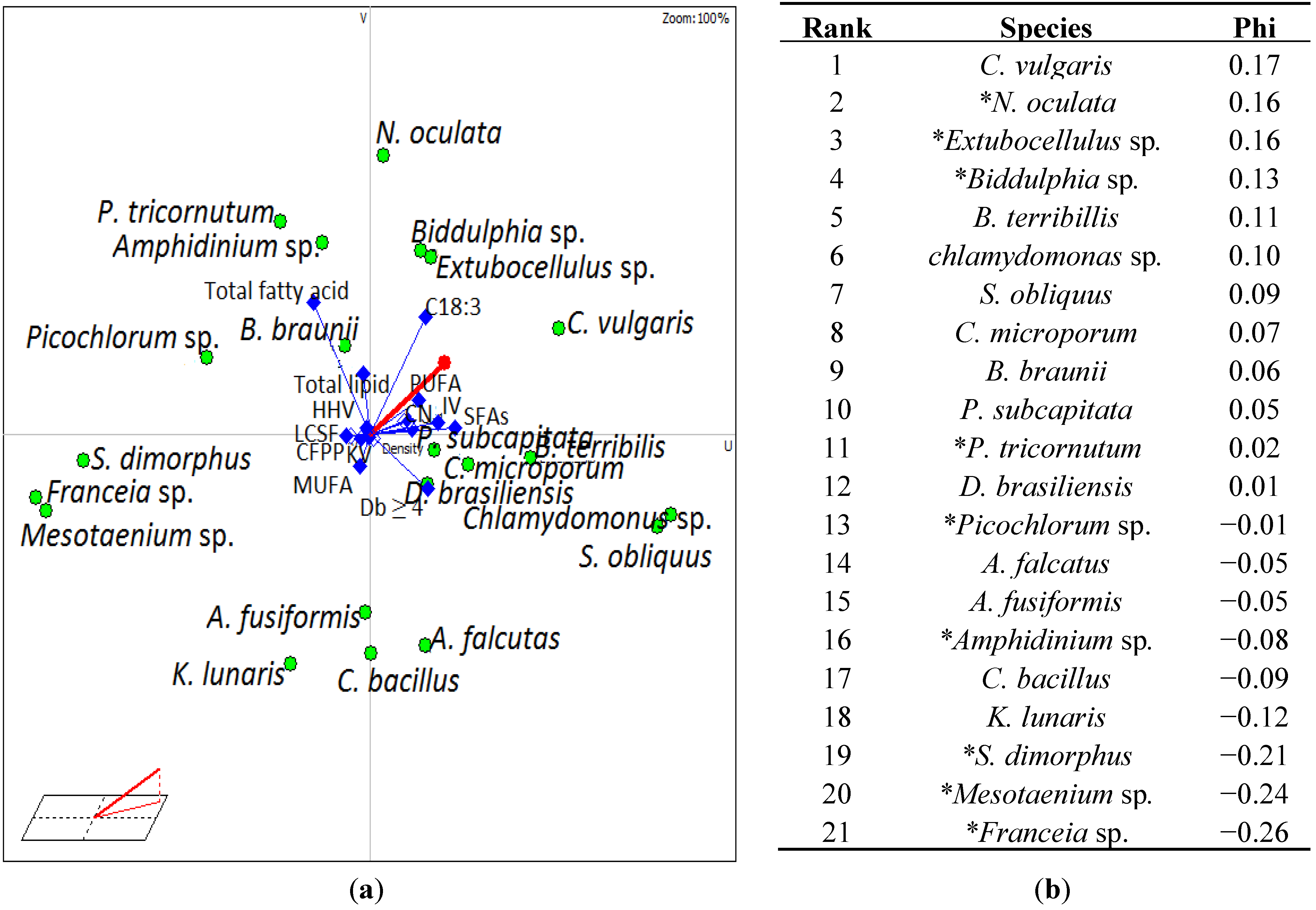{kind=link}
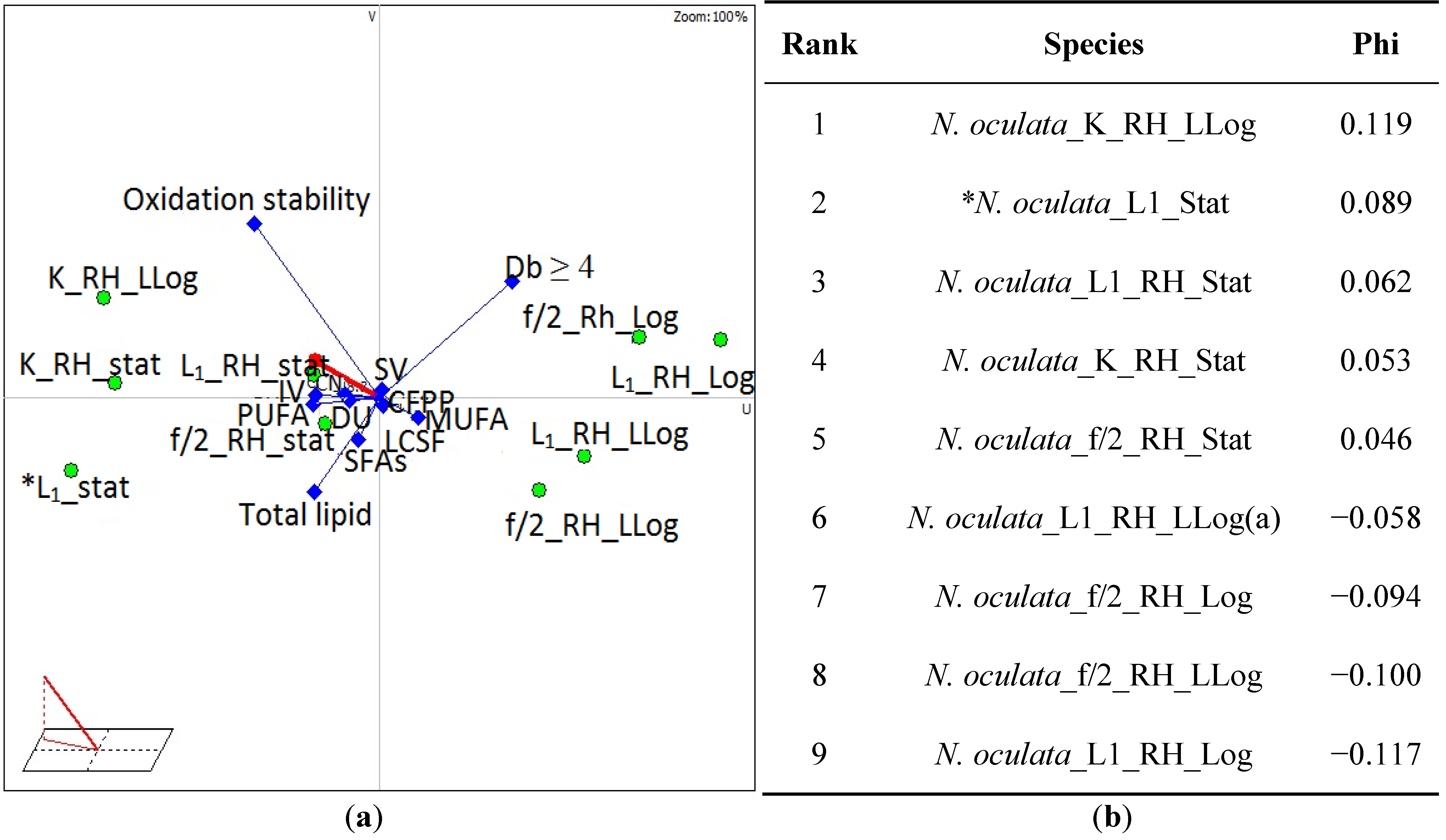{kind=link}
| Sample | Culture medium | Temp. (°C) | Total lipid (mg g−1 dwt) | Total fatty acids * (mg g−1 dwt) |
|---|---|---|---|---|
| Nine species from this study: | ||||
| NQAIF034 Amphidinium sp. | L1 | 24 ± 1 | 189 | 141.0 |
| NQAIF272 Biddulphia sp. | f/2 | 24 ± 1 | 249 | 109.3 |
| NQAIF004 Phaeodactylum tricornutum | f/2 | 24 ± 1 | 217 | 187.3 |
| NQAIF284 Picochlorum sp. | L1 | 24 ± 1 | 305 | 274.8 |
| NQAIF283 Nannochloropsis oculata | L1 | 24 ± 1 | 410 | 267.1 |
| NQAIF254 Extubocellulus sp. | L1 | 24 ± 1 | 270 | 116.9 |
| NQAIF294 Scenedesmus dimorphus | Bold | 24 ± 1 | n.d. | 84.3 |
| NQAIF301 Franceia sp. | Bold | 24 ± 1 | n.d. | 79.7 |
| NQAIF303 Mesotaenium sp. | Bold | 24 ± 1 | n.d. | 76.5 |
| Twelve species from [12]: | ||||
| Ankistrodesmus falcatus | LC | 25 ± 2 | 165 | 17.5 |
| Ankistrodesmus fusiformis | LC | 25 ± 2 | 207 | 27.3 |
| Kirchneriella lunaris | LC | 25 ± 2 | 173 | 30.5 |
| Chlamydomonas sp. | LC | 25 ± 2 | 151 | 14.1 |
| Chlamydocapsa bacillus | LC | 25 ± 2 | 135 | 19.2 |
| Coelastrum microporum | LC | 25 ± 2 | 206 | 49.1 |
| Desmodesmus brasiliensis | LC | 25 ± 2 | 180 | 37.0 |
| Scenedesmus obliquus | LC | 25 ± 2 | 167 | 4.4 |
| Pseudokirchneriella subcapitata | LC | 25 ± 2 | 284 | 36.7 |
| Chlorella vulgaris | CHU 13 | 25 ± 2 | 281 | 75.9 |
| Botryococcus braunii | CHU 13 | 25 ± 2 | 455 | 58.9 |
| Botryococcus terribilis | CHU 13 | 25 ± 2 | 490 | 16.7 |
2.2. Lipid Content, Fatty Acid Methyl Ester Analysis
2.3. Calculation of Fuel Properties from Fatty Acid Profiles
3. Results and Discussion
3.1. Lipid Content
3.2. FAME Composition
| FAME | Amphidinium sp. | Biddulphia sp. | Phaeodactylum tricornutum | Picochlorum sp. | Nannochlopsis oculata. | Extubocellulus sp. | Scenedesmus dimorphos | Franceia sp. | Mesotaenium. sp. |
|---|---|---|---|---|---|---|---|---|---|
| C8:0 | 0.0 | 0.0 | 0.0 | 0.3 | 0.5 | 0.0 | 0.0 | 0.0 | 0.0 |
| C10:0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| C12:0 | 0.0 | 0.5 | 0.0 | 0.0 | 1.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| C13:0 | 0.0 | 0.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| C 14:0 | 1.1 | 23.4 | 6.1 | 1.5 | 15.3 | 7.6 | 0.4 | 0.4 | 0.4 |
| C 14:1 | 0.0 | 0.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| C 15:0 | 0.0 | 2.4 | 0.8 | 0.7 | 1.4 | 0.5 | 0.3 | 0.5 | 0.4 |
| C 15:1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.9 | 1.7 | 1.7 |
| C 16:0 | 49.6 | 25.7 | 43.8 | 45.9 | 85.4 | 29.4 | 13.1 | 10.0 | 10.1 |
| C16:1 (7) | 1.5 | 36.3 | 89.3 | 3.3 | 78.4 | 69.6 | 1.3 | 1.1 | 0.9 |
| C 16:1 (9) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 3.0 | 4.6 | 3.6 |
| C16:2 (7,10) | 0.0 | 1.9 | 2.6 | 13.9 | 0.0 | 3.1 | 1.7 | 1.2 | 1.7 |
| C16:2 (9,12) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 | 0.6 | 0.5 |
| C16:3 (cis 6,9,12) | 0.0 | 4.9 | 0.0 | 9.5 | 0.0 | 0.0 | 0.5 | 0.5 | 0.5 |
| C16:3 (7, 10, 13) | 0.0 | 0.0 | 7.9 | 0.0 | 0.0 | 0.0 | 1.3 | 1.0 | 1.4 |
| C16:4 (4,7,10,13) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 12.9 | 13.2 | 12.3 |
| C17:0 | 0.0 | 0.0 | 0.0 | 1.0 | 1.0 | 0.0 | 0.3 | 0.4 | 0.0 |
| C17:1 | 0.0 | 0.0 | 0.0 | 0.8 | 0.8 | 0.0 | 0.0 | 0.0 | 0.0 |
| C 18:0 | 5.7 | 0.8 | 1.5 | 9.3 | 2.6 | 1.1 | 0.5 | 0.4 | 0.4 |
| C 18:1 (9) | 26.9 | 1.6 | 6.7 | 42.4 | 53.3 | 3.7 | 5.8 | 3.4 | 4.3 |
| C 18:1 (x) | 0.0 | 0.6 | 4.2 | 0.0 | 0.0 | 0.0 | 1.5 | 1.8 | 1.4 |
| C 18:2 (cis-9,12) | 0.0 | 0.0 | 0.0 | 97.6 | 3.4 | 0.0 | 10.6 | 6.6 | 8.9 |
| C18:3 all cis 6,9,12 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.8 | 1.0 | 0.7 |
| C 18:3 (all cis-9,12,15) | 0.0 | 0.0 | 0.0 | 40.6 | 0.0 | 0.0 | 20.7 | 25.1 | 22.9 |
| C18:4 (6,9,12,15) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 2.9 | 2.9 | 2.3 |
| C 20:0 | 7.9 | 0.0 | 0.0 | 5.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| C 20:2 (cis-11,14) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.6 | 0.0 | 0.0 |
| C 20:4 | 1.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| C 20:5 (all cis-5,8,11,14,17) | 16.6 | 10.5 | 22.5 | 0.0 | 21.9 | 0.0 | 0.0 | 0.7 | 0.6 |
| C 22:0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.6 | 0.0 | 0.0 |
| C 22:6 | 28.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| C 24:0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.3 | 0.6 | 0.0 |
| C 24:1 (cis-15) | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.8 | 0.0 | 0.0 |
| SFAs (%) | 46.3 | 48.8 | 28.1 | 23.7 | 40.5 | 33.6 | 18.9 | 15.9 | 15.4 |
| MUFAs (%) | 20.4 | 35.4 | 54.1 | 17.0 | 50.0 | 63.8 | 17.4 | 16.2 | 15.9 |
| PUFAs (%) | 33.3 | 15.8 | 17.8 | 59.2 | 9.5 | 2.7 | 63.7 | 67.9 | 68.7 |
3.3. Fuel Properties
| Algae species | DU | LCSF | CFPP (°C) | IV (g I2100g−1 fat) | SV (mg KOHg−1) | CN1 | CN2 | SFAs (%) | MUFA (%) | PUFA (%) | Kinematic viscosity (υ) (mm2 s−1) | Density (ρ) (g cm−3) | HHV (MJ kg−1) | C18:3 (wt%) | Db ≥ 4 (wt%) | Oxidation Stb. a (h) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biodiesel Standard EN 14214 | - | - | ≤5/≤−20 | ≤120 | - | ≥51 | ≥51 | - | - | - | 3.5–5.0 | 0.86–0.90 | NA | ≤12 | ≤1 | ≥ 6 | |
| Biodiesel Standard ASTM D6751−02 | - | - | NA | NA | - | ≥47 | ≥47 | - | - | - | 1.9–6.0 | NA | NA | - | - | - | |
| Min/max | max | max | max | max | max | min | min | min | max | max | max | max | min | max | max | ||
| Threshold value for PROMETHEE | - | - | 5 | 120 | - | 47 | 47 | - | - | - | - | 0.90 | - | 12 | 1 | - | |
| Nine species from this study: | |||||||||||||||||
| A | Nannochloropsis oculata | 69 | 3.7 | −4.8 | 81 | 203 | 55.0 | 57.9 | 40.5 | 50.0 | 9.5 | 4.2 | 0.88 | 39.8 | 0 | 8.3 | 95.7 |
| Extubocellulus sp. | 69 | 3 | −7.0 | 65 | 209 | 57.8 | 60.9 | 33.6 | 63.8 | 2.7 | 3.92 | 0.89 | 40.1 | 0 | 0 | − | |
| Biddulphia sp. | 67 | 2.7 | −7.9 | 88 | 210 | 52.5 | 54.6 | 48.8 | 35.4 | 15.8 | 3.71 | 0.89 | 40.0 | 0 | 9.6 | − | |
| B | Phaeodactylum tricornutum | 90 | 2.8 | −7.8 | 114 | 204 | 47.3 | 50.3 | 28.1 | 54.1 | 17.8 | 3.74 | 0.89 | 39.8 | 0 | 12.1 | − |
| C | Picochlorum sp. | 136 | 5.5 | 0.7 | 135 | 195 | 44.0 | 48.9 | 23.7 | 17.0 | 59.2 | 3.99 | 0.89 | 39.9 | 14.9 | 0 | 4.9 |
| Amphidinium sp. | 87 | 11.3 | 19.1 | 159 | 188 | 39.5 | 42.9 | 46.3 | 20.4 | 33.3 | 4.13 | 0.9 | 40.3 | 0 | 32.1 | − | |
| Scenedesmus dimorphus | 145 | 3.8 | −4.6 | 184 | 196 | 32.9 | 37.1 | 18.9 | 17.4 | 63.7 | 3.63 | 0.91 | 40.2 | 26 | 19.1 | 5.6 | |
| Franceia sp. | 152 | 3.1 | −6.7 | 206 | 198 | 27.7 | 33.3 | 15.9 | 16.2 | 67.9 | 3.49 | 0.91 | 40.4 | 33.5 | 21.6 | 5.4 | |
| Mesotaenium sp. | 153 | 1.6 | −11.4 | 202 | 200 | 28.3 | 33.4 | 15.4 | 15.9 | 68.7 | 3.45 | 0.91 | 40.2 | 31.4 | 20.3 | 5.3 | |
| Twelve species from literature [12]: | |||||||||||||||||
| Ankistrodesmus falcatus | 85 | 4.38 | −2.7 | 96 | 191 | 53.2 | 49.3 | 41.4 | 28.4 | 30.2 | 3.68 | 0.82 | 36.6 | 26.86 | 0 | 6.7 | |
| Ankistrodesmus fusiformis | 99 | 3.75 | −4.7 | 108 | 189 | 50.8 | 47.4 | 37.3 | 22.4 | 40.2 | 3.65 | 0.82 | 36.9 | 26.5 | 0 | 5.6 | |
| Kirchneriella lunaris | 111 | 3.53 | −5.4 | 130 | 192 | 45.4 | 45.5 | 32.1 | 23.1 | 44.8 | 3.70 | 0.85 | 38.2 | 39.6 | 0 | 5.3 | |
| Chlamydomonas sp. | 27 | 10.8 | 17.6 | 26 | 206 | 66.9 | 62.4 | 78.6 | 14.6 | 6.8 | 3.93 | 0.81 | 36.5 | 2.76 | 0 | 20.2 | |
| Chlamydocapsa bacillus | 100 | 3.93 | −4.1 | 109 | 187 | 51.0 | 48.0 | 35.7 | 23.6 | 40.7 | 3.69 | 0.83 | 37.1 | 25.45 | 0 | 5.6 | |
| Coelastrum microporum | 86 | 4.02 | −3.8 | 84 | 195 | 55.4 | 57.2 | 45.9 | 38.0 | 16.1 | 4.15 | 0.86 | 38.8 | 11.1 | 0 | 8.6 | |
| Desmodesmus brasiliensis | 87 | 4.43 | −2.6 | 83 | 195 | 55.6 | 57.9 | 34.5 | 44.1 | 21.4 | 4.18 | 0.86 | 39.0 | 9.43 | 0 | 8.1 | |
| Scenedesmus obliquus | 36 | 8.95 | 11.6 | 34 | 204 | 65.5 | 63.2 | 70.8 | 21.7 | 7.5 | 4.04 | 0.83 | 37.5 | 2.83 | 0 | 18.5 | |
| Pseudokirchneriella subcapitata | 82 | 4.23 | −3.2 | 79 | 194 | 56.7 | 57.5 | 35.4 | 47.4 | 17.3 | 4.14 | 0.85 | 38.3 | 9.87 | 0 | 9.4 | |
| Chlorella vulgaris | 56 | 8.04 | 8.8 | 50 | 189 | 63.8 | 63.3 | 52.2 | 37.5 | 10.3 | 4.28 | 0.84 | 38.1 | 1.57 | 0 | 14.3 | |
| Botryococcus braunii | 99 | 1.51 | −11.7 | 90 | 188 | 55.1 | 58.7 | 9.9 | 79.6 | 10.5 | 4.39 | 0.86 | 39.2 | 5.34 | 0 | 13.8 | |
| Botryococcus terribilis | 67 | 5.08 | −0.5 | 64 | 184 | 61.7 | 59.0 | 43.2 | 44.3 | 12.6 | 4.13 | 0.82 | 37.3 | 7.22 | 0 | 12.2 | |
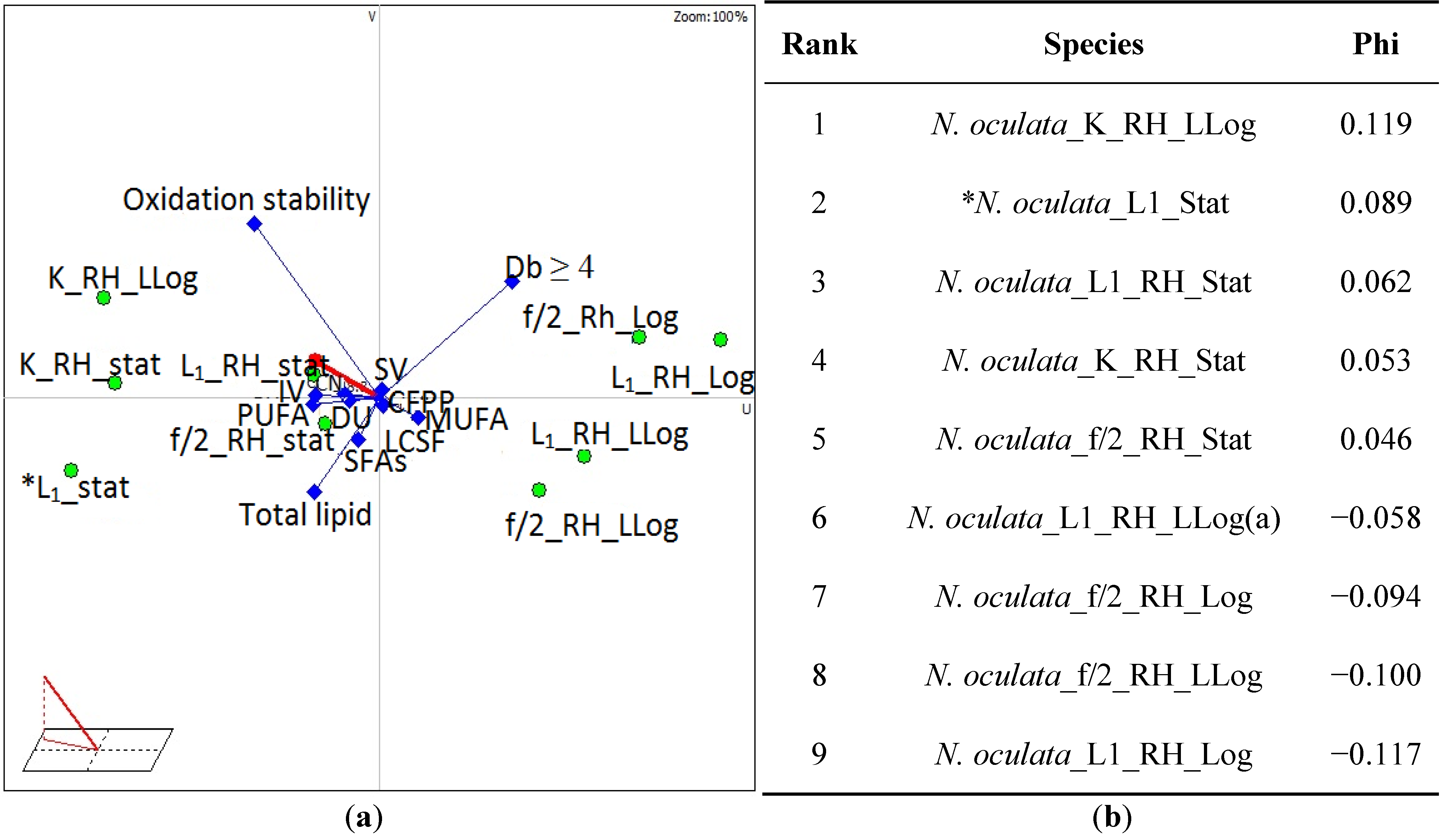
| Algae species | Growth media and growth phase | DU | LCSF | CFPP (°C) | IV (g I2100g−1 fat) | SV (mg KOHg−1) | CN1 | SFAs (%) | MUFA (%) | PUFA (%) | Total lipid (mg g−1 dwt) | C18:3 (wt%) | Db ≥ 4 (wt%) | Oxidation Stb. (h) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min/max | - | max | max | max | max | max | min | min | max | max | min | max | max | min |
| Threshold value for PROMETHEE analysis | - | - | - | 5 | 120 | - | 47 | - | - | - | - | 12 | 1 | 6 |
| * N. oculata | L1_Stat | 69 | 3.7 | −4.8 | 81 | 203 | 55.0 | 40.5 | 50.0 | 9.5 | 410 | 0 | 8.23 | 95.4 |
| N. oculata_RH | L1_Log | 107 | 4.0 | −4.0 | 175 | 195 | 35.0 | 31.9 | 28.6 | 39.4 | 213 | 0 | 37.10 | 56.2 |
| N. oculata_RH | L1_LLog a | 96 | 4.1 | −3.7 | 155 | 198 | 39.9 | 37.7 | 29.1 | 33.4 | 313 | 0 | 31.1 | 56.2 |
| N. oculata_RH | L1_Stat | 75 | 4.9 | −1.1 | 107 | 200 | 49.5 | 44.8 | 35.2 | 20.1 | 327 | 0 | 18.6 | 81.2 |
| N. oculata_RH | f/2_Log | 102 | 4.1 | −3.5 | 163 | 196 | 37.4 | 33.9 | 29.8 | 36.2 | 219 | 0 | 34.1 | 58.7 |
| N. oculata_RH | f/2_LLog | 97 | 4.5 | −2.2 | 154 | 208 | 38.0 | 40.2 | 28.7 | 31.3 | 292 | 0.1 | 19.1 | 56.2 |
| N. oculata_RH | f/2_Stat | 74 | 5.1 | −0.4 | 105 | 200 | 49.9 | 45.9 | 34.1 | 20.0 | 332 | 0 | 18.4 | 76.3 |
| N. oculata_RH | K_LLog | 70 | 4.5 | −2.5 | 83 | 203 | 54.3 | 40.2 | 47.1 | 12.6 | 357 | 0.1 | 11.7 | 150 |
| N. oculata_RH | K_Stat | 79 | 4.5 | −2.4 | 113 | 200 | 48.0 | 40.8 | 48.8 | 10.4 | 378 | 0.1 | 9.4 | 120.5 |
3.4. Selection of Suitable Algae Species for Biodiesel
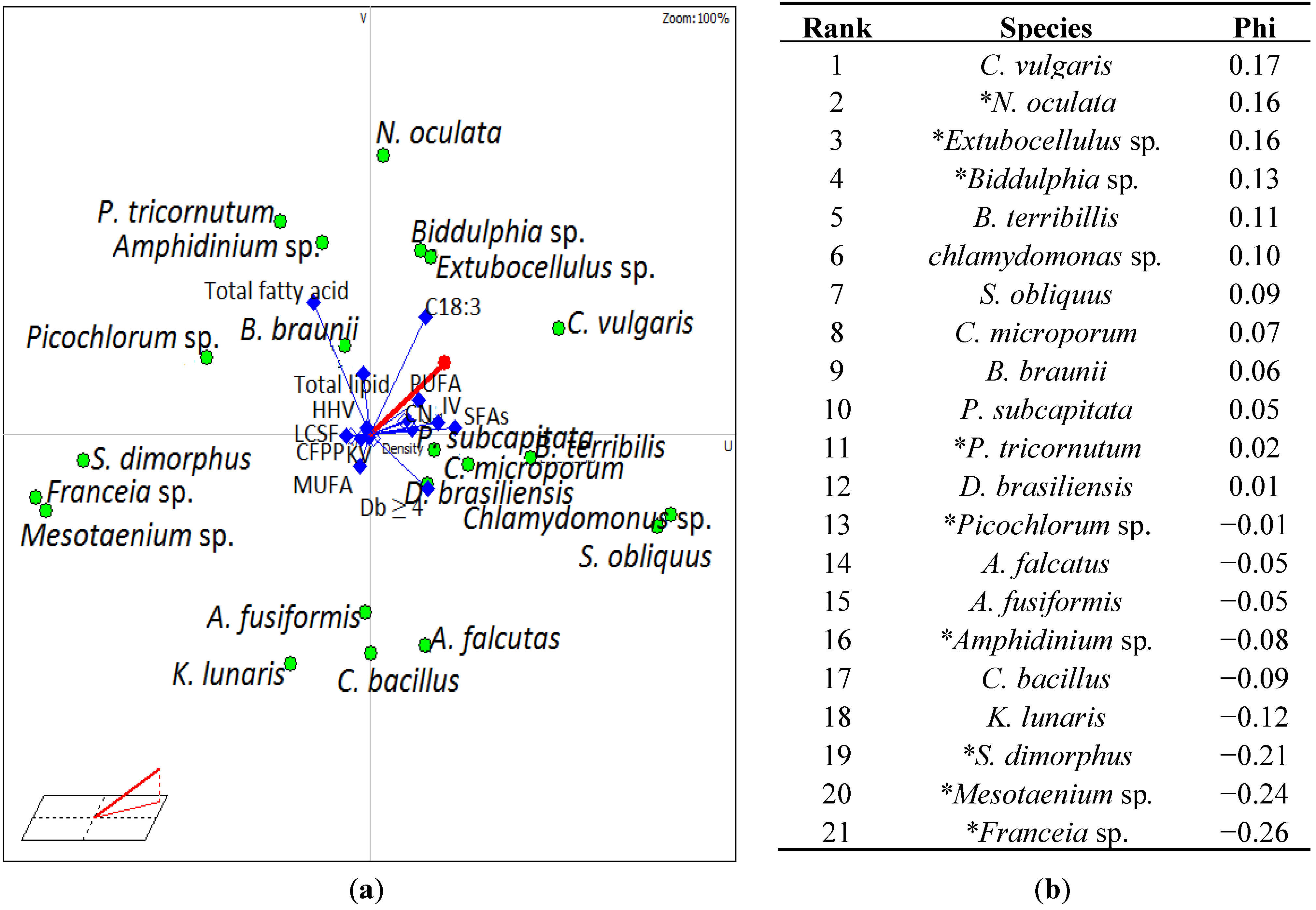
| Species | Comparative Rank shift with different PUFA weighting | |||||
|---|---|---|---|---|---|---|
| All weight = 1 | PUFA weight = 10 | PUFA weight = 30 | PUFA weight = 40 | PUFA weight = 50 | Direction of rank shift | |
| C. vulgaris | 1 | 3 | 4 | 5 | 5 |  |
| *N. oculata | 2 | 2 | 3 | 3 | 4 | |
| *Extubocellulus sp. | 3 | 1 | 1 | 1 | 1 |  |
| *Biddulphia sp. | 4 | 7 | 8 | 8 | 8 | |
| B. terribillis | 5 | 6 | 6 | 7 | 7 | |
| chlamydomonas sp. | 6 | 4 | 2 | 2 | 2 | |
| S. obliquus | 7 | 5 | 5 | 4 | 3 | |
| C. microporum | 8 | 9 | 9 | 9 | 9 |  |
| B. braunii | 9 | 8 | 7 | 6 | 6 | |
| P. subcapitata | 10 | 10 | 10 | 10 | 10 | - |
| *P. tricornutum | 11 | 11 | 11 | 11 | 11 | - |
| D. brasiliensis | 12 | 12 | 12 | 12 | 12 | - |
| *Picochlorum sp. | 13 | 18 | 18 | 18 | 18 | - |
| A. falcatus | 14 | 13 | 13 | 13 | 13 | - |
| A. fusiformis | 15 | 15 | 15 | 15 | 15 | - |
| *Amphidinium sp. | 16 | 14 | 14 | 14 | 14 | - |
| C. bacillus | 17 | 16 | 16 | 16 | 16 | - |
| K. lunaris | 18 | 17 | 17 | 17 | 17 | - |
| *S. dimorphus | 19 | 19 | 19 | 19 | 19 | - |
| *Mesotaenium sp. | 20 | 20 | 21 | 21 | 21 |  |
| *Franceia sp. | 21 | 21 | 20 | 20 | 20 | |
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hossain, A.S.; Salleh, A.; Boyce, A.N.; Chowdhury, P.; Naqiuddin, M. Biodiesel fuel production from algae as renewable energy. Am. J. Biochem. Biotechnol. 2008, 4, 250–254. [Google Scholar] [CrossRef]
- Oncel, S.S. Microalgae for a macroenergy world. Renew. Sustain. Energy Rev. 2013, 26, 241–264. [Google Scholar] [CrossRef]
- Rosillo-Calle, F.; Thrän, D.; Seiffert, M.; Teelucksingh, S. The Potential Roll of Biofuels in Commercial Air Transport—BioJetFuel; IEA Bioenergy Task 40 Sustainable International Bioenergy Trade: Paris, France, 2012; Volume 40. [Google Scholar]
- Rantanen, L.; Linnaila, R.; Aakko, P.; Harju, T. NExBTL—Biodiesel Fuel of the Second Generation. In Proceedings of the Powertrain & Fluid Systems Conference & Exhibition, San Antonio, TX, USA, 24–27 September 2005.
- Kuronen, M.; Mikkonen, S.; Aakko, P.; Murtonen, T. Hydrotreated Vegetable Oil as Fuel for Heavy Duty Diesel Engines. In Proceedings of the Powertrain & Fluid Systems Conference & Exhibition, Detroit, MI, USA, 16–19 April 2007.
- Shay, E.G. Diesel fuel from vegetable oils: Status and opportunities. Biomass Bioenergy 1993, 4, 227–242. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Brennan, L.; Owende, P. Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co-products. Renew. Sustain. Energy Rev. 2010, 14, 557–577. [Google Scholar] [CrossRef]
- Esteban, G.F.; Finlay, B.J. Cryptic freshwater ciliates in a hypersaline lagoon. Protist 2003, 154, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Huerlimann, R.; De Nys, R.; Heimann, K. Growth, lipid content, productivity, and fatty acid composition of tropical microalgae for scale-up production. Biotechnol. Bioeng. 2010, 107, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, A.; Puhan, S.; Nagarajan, G. Relating the cetane number of biodiesel fuels to their fatty acid composition: A critical study. Proc. Inst. Mech. Eng. Part D J. Automob. Eng. 2009, 223, 565–583. [Google Scholar] [CrossRef]
- Nascimento, I.A.; Marques, S.S.I.; Cabanelas, I.T.D.; Pereira, S.A.; Druzian, J.I.; de Souza, C.O.; Vich, D.V.; de Carvalho, G.C.; Nascimento, M.A. Screening microalgae strains for biodiesel production: Lipid productivity and estimation of fuel quality based on fatty acids profiles as selective criteria. Bioenergy Res. 2013, 6, 1–13. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Huang, B.-Y.; Chiang, T.-H.; Tang, T.-C. Fuel properties of microalgae (Chlorella protothecoides) oil biodiesel and its blends with petroleum diesel. Fuel 2012, 94, 270–273. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernandez, C.M.; Casas, A.; Rodriguez, L.; Perez, A. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Krisnangkura, K. A simple method for estimation of cetane index of vegetable oil methyl esters. J. Am. Oil Chem. Soc. 1986, 63, 552–553. [Google Scholar] [CrossRef]
- Gouw, T.; Vlugter, J. Physical properties of fatty acid methyl esters. I. Density and molar volume. J. Am. Oil Chem. Soc. 1964, 41, 142–145. [Google Scholar] [CrossRef]
- Allen, C.; Watts, K.; Ackman, R.; Pegg, M. Predicting the viscosity of biodiesel fuels from their fatty acid ester composition. Fuel 1999, 78, 1319–1326. [Google Scholar] [CrossRef]
- Hoekman, S.K.; Broch, A.; Robbins, C.; Ceniceros, E.; Natarajan, M. Review of biodiesel composition, properties, and specifications. Renew. Sustain. Energy Rev. 2012, 16, 143–169. [Google Scholar] [CrossRef]
- Barabás, I.; Todoruţ, I.A. Biodiesel Quality, Standards and Properties. In Biodiesel-Quality, Emissions and By-Products; Montero, G., Stoytcheva, M., Eds.; InTech.: Rijeka, Croacia, 2011; pp. 3–28. [Google Scholar]
- Park, J.-Y.; Kim, D.-K.; Lee, J.-P.; Park, S.-C.; Kim, Y.-J.; Lee, J.-S. Blending effects of biodiesels on oxidation stability and low temperature flow properties. Bioresour. Technol. 2008, 99, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Somersalo, S.; Karunen, P.; Aro, E.-M. The acyl lipid composition of wheat leaves and moss protonemata using a new, non-carcinogenic extraction solvent system. Physiol. Plant. 1986, 68, 467–470. [Google Scholar] [CrossRef]
- Gosch, B.J.; Magnusson, M.; Paul, N.A.; Nys, R. Total lipid and fatty acid composition of seaweeds for the selection of species for oil-based biofuel and bioproducts. GCB Bioenergy 2012, 4, 919–930. [Google Scholar] [CrossRef]
- David, F.; Sandra, P.; Wylie, P.L. Improving the Analysis of Fatty Acid Methyl Esters Using Retention Time Locked Methods and Retention Time Databases. In Agilent Technologies—Application; Agilent Technologies: Palo Alto, CA, USA, 2002. [Google Scholar]
- Klopfenstein, W. Estimation of cetane index for esters of fatty acids. J. Am. Oil Chem. Soc. 1982, 59, 531–533. [Google Scholar] [CrossRef]
- Ramírez-Verduzco, L.F.; Rodríguez-Rodríguez, J.E.; Jaramillo-Jacob, A.R. Predicting cetane number, kinematic viscosity, density and higher heating value of biodiesel from its fatty acid methyl ester composition. Fuel 2012, 91, 102–111. [Google Scholar] [CrossRef]
- Kalayasiri, P.; Jeyashoke, N.; Krisnangkura, K. Survey of seed oils for use as diesel fuels. J. Am. Oil Chem.Soc. 1996, 73, 471–474. [Google Scholar] [CrossRef]
- Guitouni, A.; Martel, J.-M. Tentative guidelines to help choosing an appropriate MCDA method. Eur. J. Oper. Res. 1998, 109, 501–521. [Google Scholar] [CrossRef]
- Brans, J.P.; Mareschal, B. The PROMCALC & GAIA decision support system for multicriteria decision aid. Decis. Support Syst. 1994, 12, 297–310. [Google Scholar] [CrossRef]
- Olmstead, I.L.D.; Hill, D.R.A.; Dias, D.A.; Jayasinghe, N.S.; Callahan, D.L.; Kentish, S.E.; Scales, P.J.; Martin, G.J.O. A quantitative analysis of microalgal lipids for optimization of biodiesel and omega-3 production. Biotechnol. Bioeng. 2013, 110, 2096–2104. [Google Scholar] [CrossRef] [PubMed]
- Von Alvensleben, N.; Stookey, K.; Magnusson, M.; Heimann, K. Salinity tolerance of Picochlorum atomus and the use of salinity for contamination control by the freshwater cyanobacterium Pseudanabaena limnetica. PLoS One 2013, 8. [Google Scholar] [CrossRef]
- Ben-Amotz, A. Bioactive Compounds: Glycerol Production, Carotenoid Production, Fatty Acids Production. In The Alga Dunaliella, Biodiversity, Physiology, Genomics and Biotechnology; Science Publishers: Enfield, NH, USA, 2009; pp. 189–207. [Google Scholar]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar] [CrossRef]
- Knothe, G. Improving biodiesel fuel properties by modifying fatty ester composition. Energy Environ. Sci. 2009, 2, 759–766. [Google Scholar] [CrossRef]
- Brown, M.R. Nutritional Value and Use of Microalgae in Aquaculture. In Avances en Nutrición Acuícola VI. Memorias del VI Simposium Internacional de Nutrición Acuícola (in Spanish); Cruz-Suárez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Gaxiola-Cortés, M.G., Simoes, N., Eds.; Cancún: Quintana Roo, México, 2002; pp. 281–292. [Google Scholar]
- Knothe, G. Dependence of biodiesel fuel properties on the structure of fatty acid alkyl esters. Fuel Process. Technol. 2005, 86, 1059–1070. [Google Scholar] [CrossRef]
- Lapuerta, M.; Rodríguez-Fernández, J.; De Mora, E.F. Correlation for the estimation of the cetane number of biodiesel fuels and implications on the iodine number. Energy Policy 2009, 37, 4337–4344. [Google Scholar] [CrossRef]
- Schenk, P.M.; Thomas-Hall, S.R.; Stephens, E.; Marx, U.C.; Mussgnug, J.H.; Posten, C.; Kruse, O.; Hankamer, B. Second generation biofuels: High-efficiency microalgae for biodiesel production. Bioenergy Res. 2008, 1, 20–43. [Google Scholar] [CrossRef]
- Pratas, M.J.; Freitas, S.; Oliveira, M.B.; Monteiro, S.C.; Lima, A.S.; Coutinho, J.A.P. Densities and viscosities of fatty acid methyl and ethyl esters. J. Chem. Eng. Data 2010, 55, 3983–3990. [Google Scholar] [CrossRef]
- Moser, B.R. Impact of fatty ester composition on low temperature properties of biodiesel-petroleum diesel blends. Fuel 2014, 115, 500–506. [Google Scholar] [CrossRef]
- Zhu, L.; Cheung, C.S.; Zhang, W.G.; Huang, Z. Combustion, performance and emission characteristics of a DI diesel engine fueled with ethanol-biodiesel blends. Fuel 2011, 90, 1743–1750. [Google Scholar] [CrossRef]
- Kumar Tiwari, A.; Kumar, A.; Raheman, H. Biodiesel production from jatropha oil (Jatropha curcas) with high free fatty acids: An optimized process. Biomass Bioenergy 2007, 31, 569–575. [Google Scholar] [CrossRef]
- Ng, J.-H.; Ng, H.K.; Gan, S. Characterisation of engine-out responses from a light-duty diesel engine fuelled with palm methyl ester (PME). Appl. Energy 2012, 90, 58–67. [Google Scholar] [CrossRef]
- Ng, J.-H.; Ng, H.K.; Gan, S. Engine-out characterisation using speed–load mapping and reduced test cycle for a light-duty diesel engine fuelled with biodiesel blends. Fuel 2011, 90, 2700–2709. [Google Scholar] [CrossRef]
- Espinasse, B.; Picolet, G.; Chouraqui, E. Negotiation support systems: A multi-criteria and multi-agent approach. Eur. J. Oper. Res. 1997, 103, 389–409. [Google Scholar] [CrossRef]
- Brans, J.P.; Mareschal, B. PROMETHEE Methods. In Multiple Criteria Decision Analysis: State of the Art Surveys; Springer: New York, NY, USA, 2005; pp. 163–186. [Google Scholar]
- Liang, Y.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Islam, M.A.; Magnusson, M.; Brown, R.J.; Ayoko, G.A.; Nabi, M.N.; Heimann, K. Microalgal Species Selection for Biodiesel Production Based on Fuel Properties Derived from Fatty Acid Profiles. Energies 2013, 6, 5676-5702. https://doi.org/10.3390/en6115676
Islam MA, Magnusson M, Brown RJ, Ayoko GA, Nabi MN, Heimann K. Microalgal Species Selection for Biodiesel Production Based on Fuel Properties Derived from Fatty Acid Profiles. Energies. 2013; 6(11):5676-5702. https://doi.org/10.3390/en6115676
Chicago/Turabian StyleIslam, Muhammad Aminul, Marie Magnusson, Richard J. Brown, Godwin A. Ayoko, Md. Nurun Nabi, and Kirsten Heimann. 2013. "Microalgal Species Selection for Biodiesel Production Based on Fuel Properties Derived from Fatty Acid Profiles" Energies 6, no. 11: 5676-5702. https://doi.org/10.3390/en6115676