Potential Species Distribution of Balsam Fir Based on the Integration of Biophysical Variables Derived with Remote Sensing and Process-Based Methods


Abstract
:
1. Introduction
2. Methods
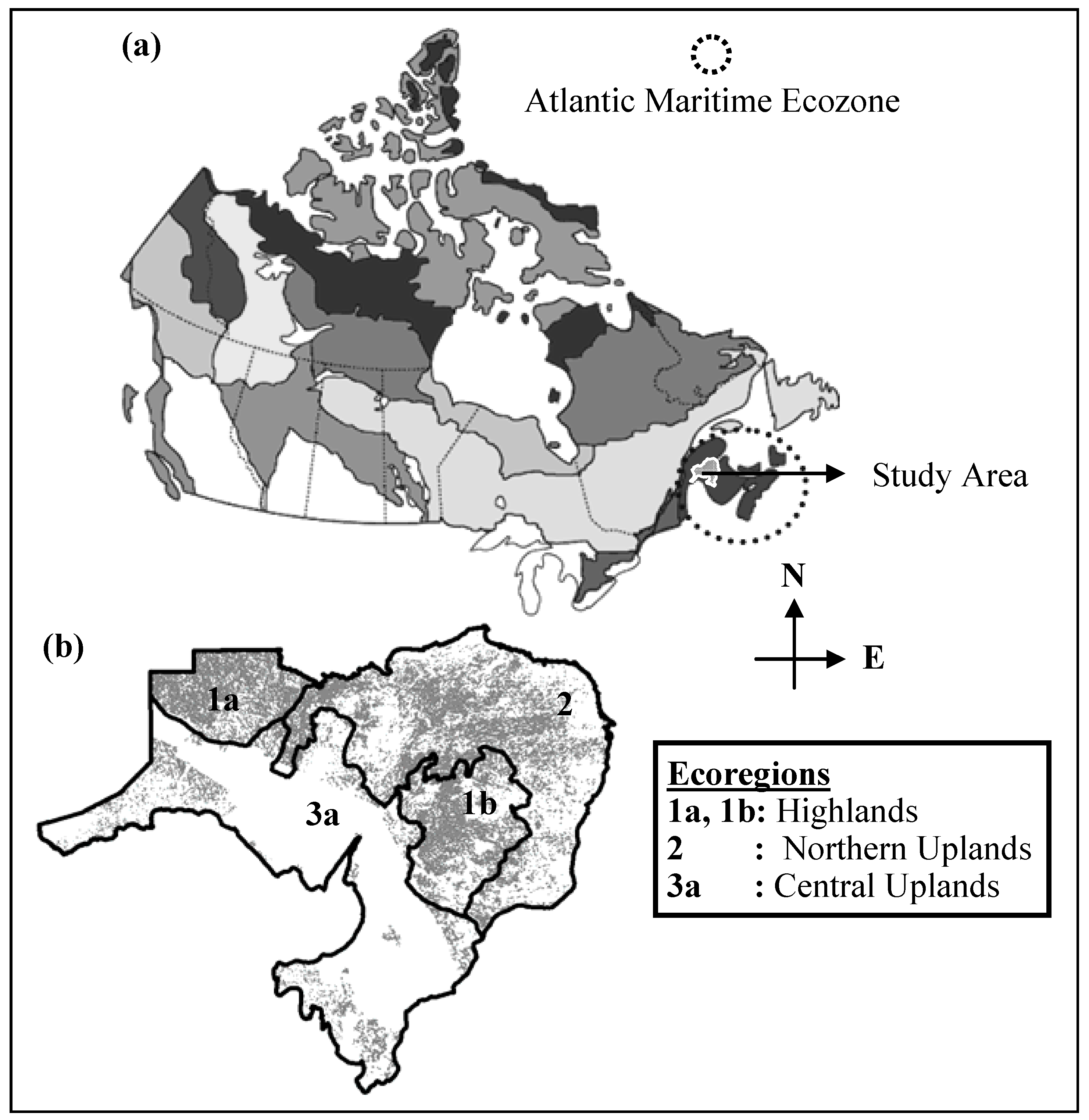
2.1. General Description of Study Area
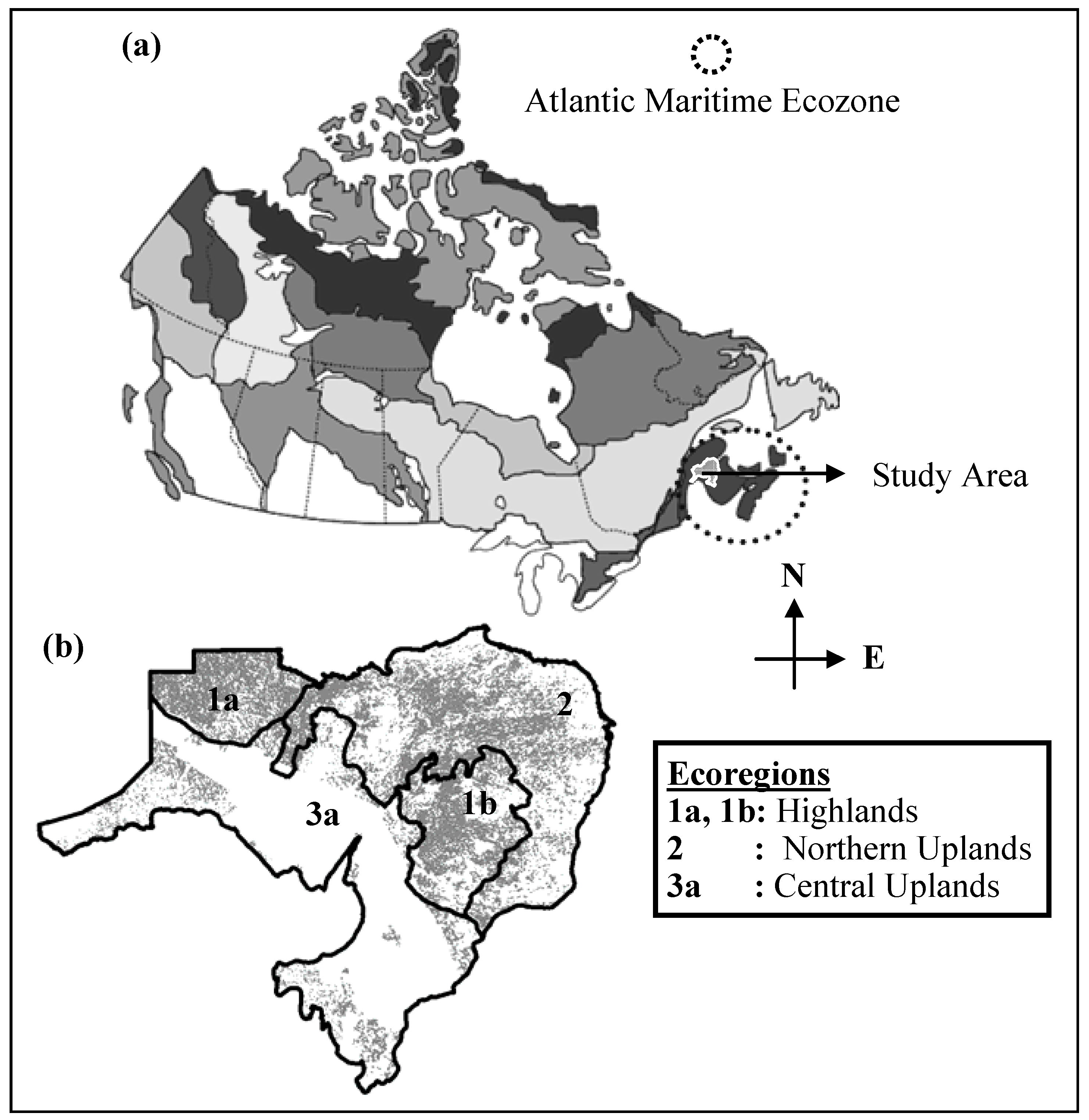
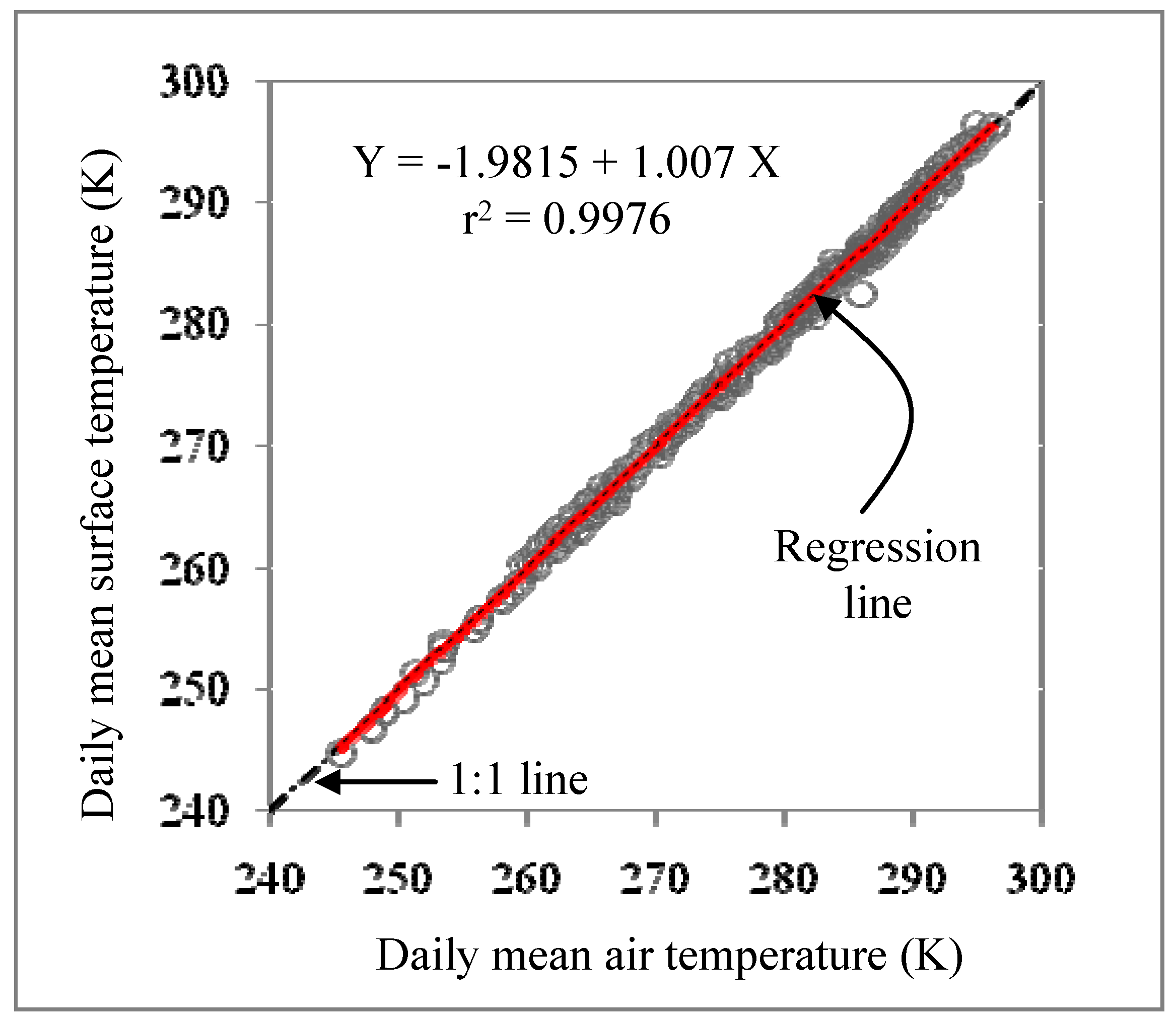
2.2. Data Requirements and Processing
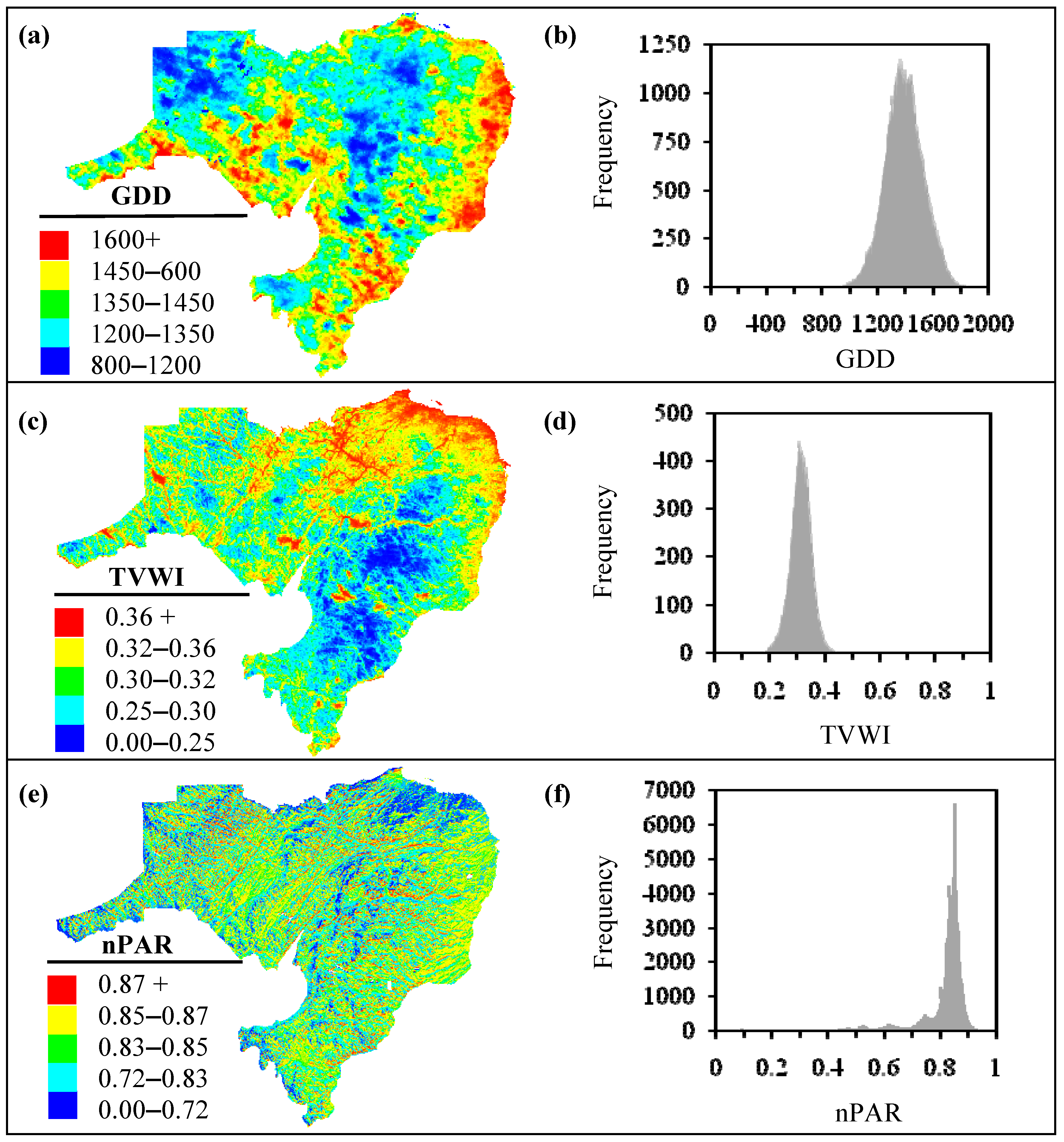
2.2.1. Growing Degree Days (GDD)
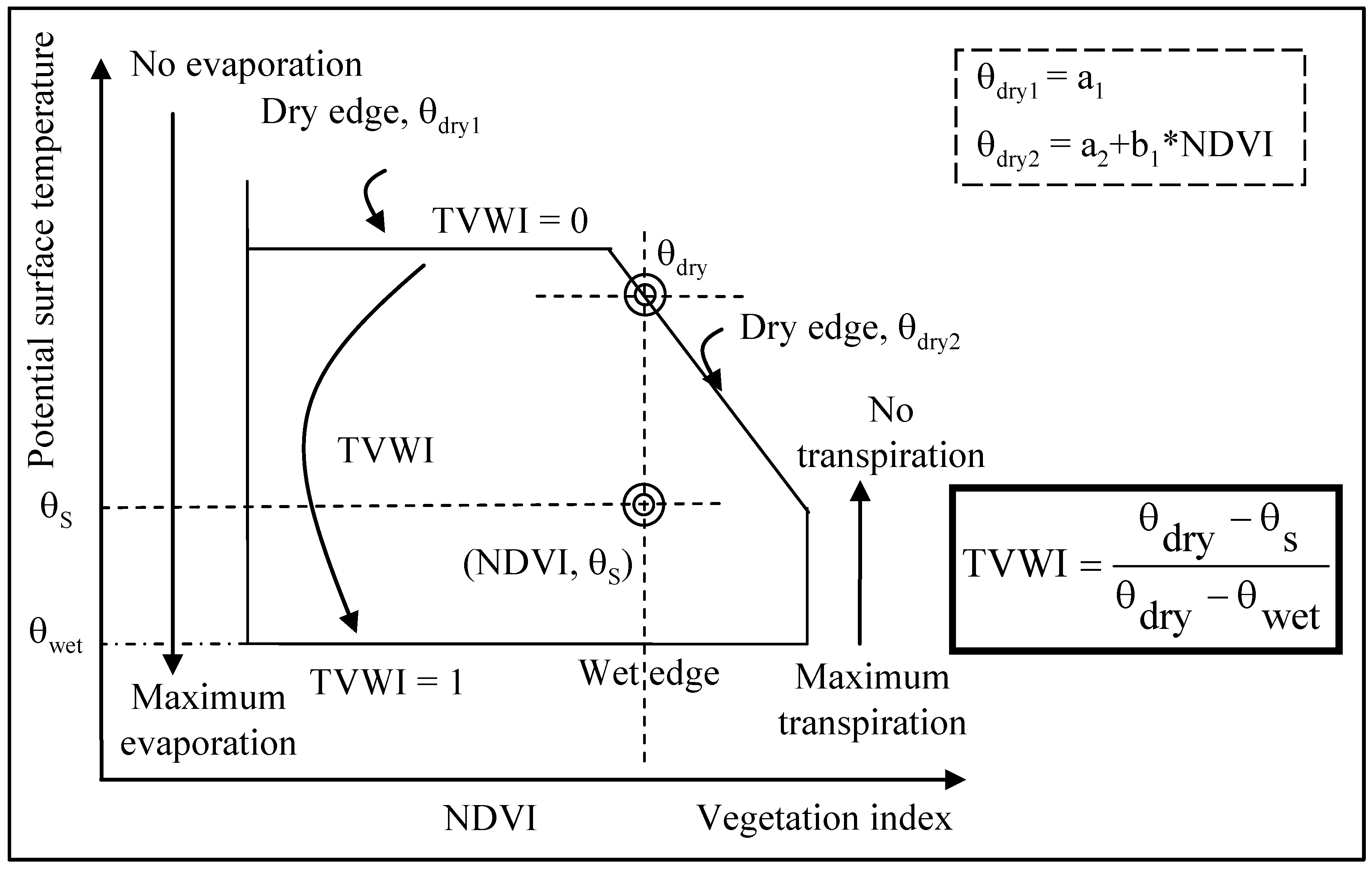
2.2.2. Temperature-Vegetation Wetness Index (TVWI)
2.2.3. Photosynthetically Active Radiation (PAR)
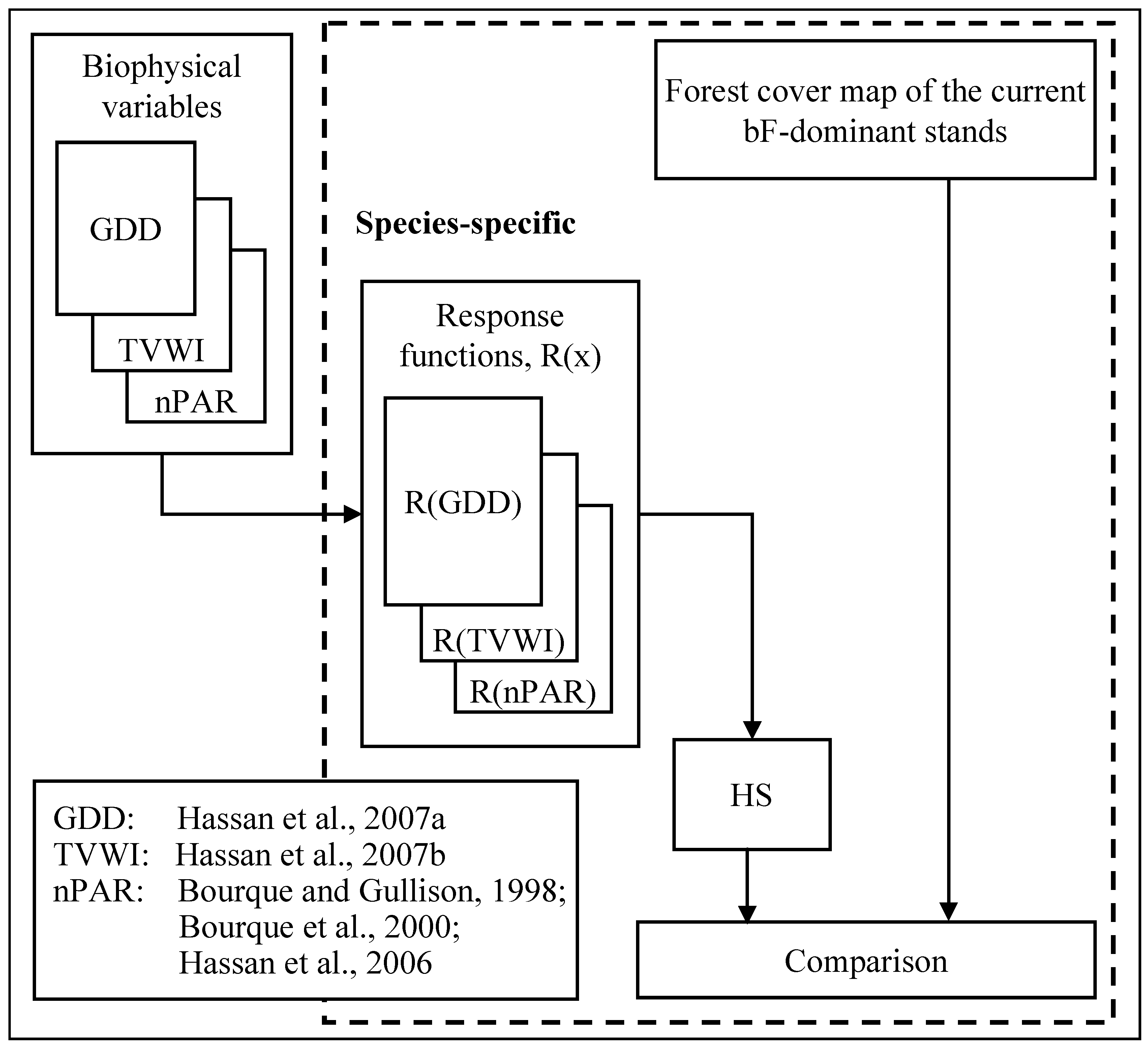
2.3. Modelling of PSD

3. Results and Discussion
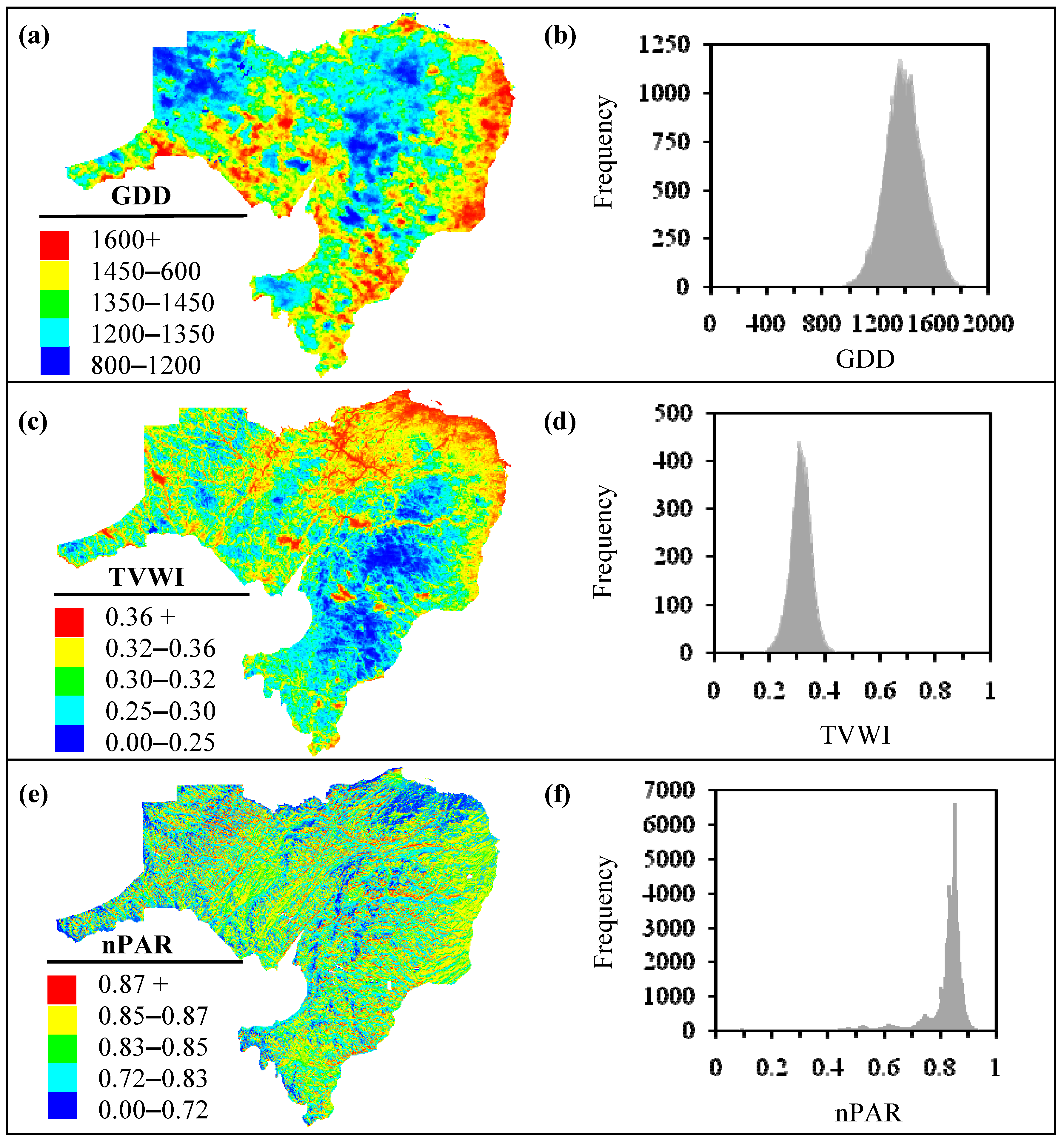
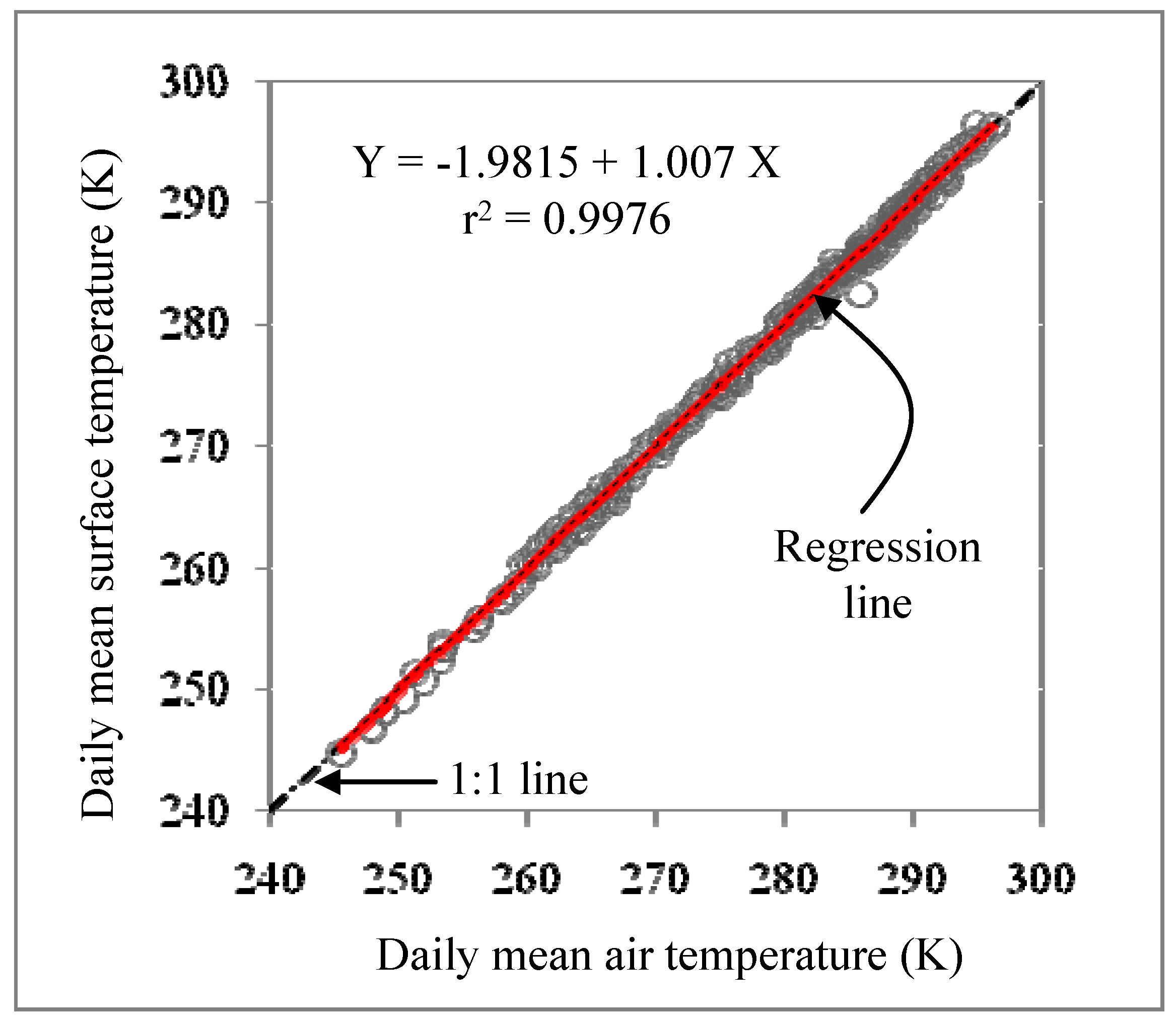
3.1. Biophysical Variables
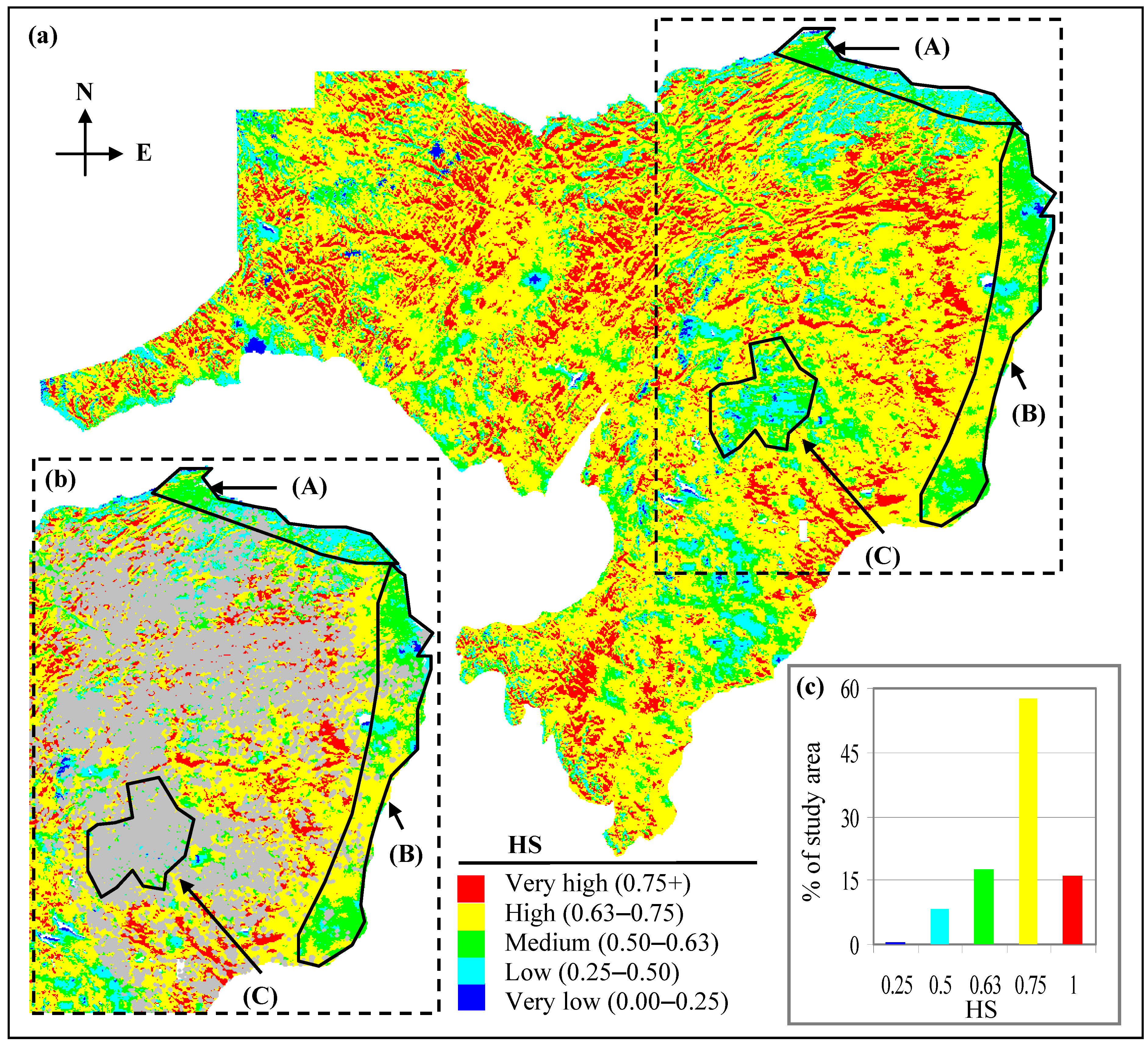
3.2. Spatial Distribution of PSD in Comparison to Existing Forest-cover
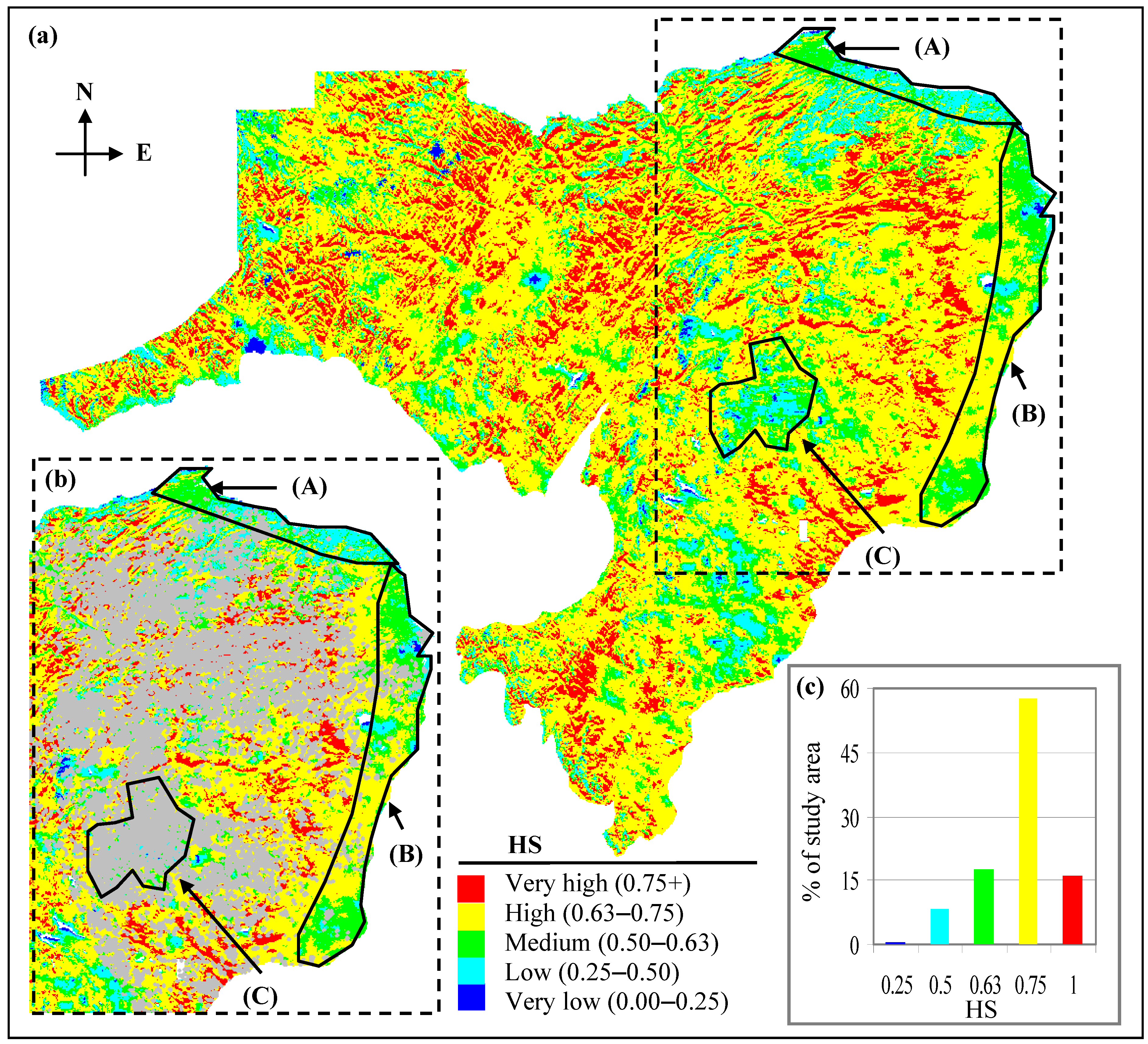
- Polygon A shows an area with low-to-medium HS values (i.e., in between 0.25–0.63; Figure 6a,b). This area is prone to lower PAR, increased TVWI due to physiographic position, and relatively high GDD as a result of the area’s proximity to the Bay of Chaleur to the north. This area most likely provides a better growing environment for temperate, hardwood species. This is consistent with species distribution maps produced by Godin and Roberts [23]. The few bF stands in the area (Figure 6b) support this interpretation.
- 2.
- Polygon B shows an area that is partially affected by a rain shadow produced by the Central Uplands of NB (producing lower TVWI); dominant HS values in the area are in the medium-to-high value range (i.e., 0.50–0.75; Figure 6a,b). However, the lack of bF stands in the area (Figure 6b) contradicts what we would expect with high HS. This may be related to the fact that bF in most of these stands were affected by spruce budworm attack during 1970–1990 [32] and past management activities (D.E. Swift, personal communication, 2009). Currently, bF resides in the understory and is anticipated to become prominent with the release and growth of bF in the understory as the stand nears maturity (D.E. Edwin Swift, personal communication, 2009). Also, high GDDs in the area potentially favour growth of hardwood species, which would tend to out-compete bF, potentially relegating bF (a shade-tolerant species) to the understory.
- 3.
- Polygon C shows areas with HS mostly in the low-to-medium value range (i.e., 0.25–0.63; Figure 6a,b). The current forest-cover map indicates many bF stands in the area (Figure 6b). This profusion of bF, mostly at elevations >450 m amsl, is indicative of reduced species competition as a result of lower GDD, which typically favour the growth of bF (D.E. Swift, personal communication, 2009), albeit at reduced rates.
- 4.
- Medium-to-very high HS (0.50–1; Figure 6a) predominate the study area (~91% of the landbase; Figure 6c). Moderate-to-low HS (0.25–0.50; Figure 6a,c) are found on about 8% of the area. Poor bF habitat (with HS < 0.25; Figure 6a,c) constitutes <1% of the total land area, and is found mostly in the warmer and wetter areas of the landscape.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand bF content (“%”) | Number of stands (n) | “%” of stands by HS range | |
|---|---|---|---|
| <0.5 (Very low-to-low) | >0.5 (Medium-to-very high) | ||
| 50–60% | 11035 | 6.7% | 93.3% |
| 60–70% | 9470 | 7.3% | 92.7% |
| 70–80% | 4927 | 9.1% | 90.9% |
| 80–90% | 1771 | 11.8% | 88.2% |
| 90–100% | 886 | 14.1% | 85.9% |
| 100% | 31 | 16.1% | 83.9% |
| 50–100% | 28120 | 7.9% | 92.1% |
3.3. Future Considerations
- Ecoregion 3a (Figure 1b) has limited forest cover indicated compared to the other regions as most of the land in that region is owned by private companies, limiting the evaluation of HS and PSD for that area.
- Implementation of the framework to address other species might require investigating environmental responses for those species (e.g., [5,33]). Although it is reasonable to assume, as a first approximation, that the effect of temperature and SWC can be reasonably represented as parabolic functions; it is likely, however, that the relationships are asymmetrical and vary according to tree species (e.g., temperature optimum), soil texture, and available soil water [34]. Over periods of 30 days or more, the assumption that photosynthesis (and growth) is a linear function of absorbed PAR is realistic, as long as stomatal conductance is not limiting and leaves are present. However, this assumption may be less realistic at shorter time intervals.
- The process of normalization, as applied in northwest NB, should be applied to other biomes in a similar manner (i.e., normalization is biome specific).
3.4. Comparison of PSD Methods
- Currently, many PSD studies use interpolation to derive input surfaces of biophysical variables in the development of PSD maps. In our work, we employ RS data and process-based modelling techniques to eliminate the inherent problems associated with interpolation of sparse data.
- The PSD work of Saatchi et al. explicitly uses RS data [18]. However, predicting the future state of the landscape under scenarios of climate change would be impossible because their methods lack the capability to predict future states. In contrast, our method can be used to predict future PSD of selected tree species (a) by directly incorporating Global Circulation Model (GCM)-projected increases in temperature in the GDD map, and (b) by replacing estimates of TVWI with SWC derived with the soil-hydrology component of LanDSET. Procedures to carry out such work are addressed in a recent paper [36] discussing the effects of climate change on species distribution in the Acadian forest region of eastern Nova Scotia.
- Most studies in PSD modelling are applied at spatial resolutions of 1–10 km [e.g., 12,14,15,35]. As a result, the utility of the information to forest management and wood supply analysis and planning professionals is limited. Our method can generate realistic maps of PSD at 250-m resolution with existing MODIS data and landscape-process modelling techniques.
4. Concluding Remarks
Acknowledgements
References and Notes
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology, 9th ed.; John Willey & Sons: New York, NY, USA, 1997. [Google Scholar]
- Austin, M.P. Spatial prediction of species distribution: an interface between ecological theory and statistical modeling. Ecol. Model. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Tsoar, A.; Alloche, O.; Steinitz, O.; Rotem, D.; Kadmon, R. A comparative evaluation of presence-only methods for modelling species distribution. Diversity Distrib. 2007, 13, 397–405. [Google Scholar] [CrossRef]
- Aussenac, G. Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Ann. Forest Sci. 2000, 57, 287–301. [Google Scholar] [CrossRef]
- Bourque, C.P.-A.; Meng, F.-R.; Gullison, J.J.; Bridgland, J. Biophysical and potential vegetation growth surfaces for a small watershed in northern Cape Breton Island, Nova Scotia, Canada. Can. J. Forest Res. 2000, 30, 1179–1195. [Google Scholar] [CrossRef]
- Ung, C.-H.; Bernier, P.Y.; Raulier, F.; Fournier, R.A.; Lambert, M.-C.; Regniere, J. Biophysical site indices for shade tolerant and intolerant boreal species. Forest Sci. 2001, 47, 83–95. [Google Scholar]
- Gustafson, E.J.; Lietz, S.M.; Wright, J.L. Predicting the spatial distribution of aspen growth potential in the upper Great Lakes region. Forest Sci. 2003, 49, 499–508. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predicting habitat distribution in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Bourque, C.P.-A.; Cox, R.M.; Allen, D.J.; Arp, P.A.; Meng, F.-R. Spatial extent of winter thaw events in eastern North America: Historical weather records in relation to yellow birch decline. Glob. Change Biol. 2005, 11, 1477–1492. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Foody, G.M.; Rocchini, D.; Giorgi, A.P.; Saatchi, S. Measuring and modelling biodiversity from space. Prog. Phys. Geog. 2008, 32, 203–221. [Google Scholar] [CrossRef]
- McKenny, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchinson, M.F. Potential impacts of climate change on the distribution of North American trees. BioScience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- McKenny, D.W.; Price, D.; Papadapol, P.; Siltanen, M.; Lawrence, K. High-Resolution Climate Change Scenarios for North America. Sault Ste. Marie (Canada): Natural Resources Canada, Front Line Technical Note No. 107; 2006. [Google Scholar]
- Latta, G.; Temesgen, H.; Barrett, T.M. Mapping and imputing potential productivity of Pacific Northwest forests using climate variables. Can. J. Forest Res. 2009, 39, 1197–1207. [Google Scholar] [CrossRef]
- Monserud, R.A.; Yang, Y.; Huang, S.; Tchebakova, N. Potential change in lodgepole pine site index and distribution under climatic change in Alberta. Can. J. Forest Res. 2008, 38, 343–352. [Google Scholar] [CrossRef]
- Peng, G.; Li, J.; Chen, Y.; Norizan, A.P.; Tay, L. High-resolution surface relative humidity computation using MODIS image in Peninsular Malaysia. Chin. Geog. Sci. 2006, 16, 260–264. [Google Scholar] [CrossRef]
- Bourque, C.P.-A.; Gullison, J.J. A technique to predict hourly potential solar radiation and temperature for a mostly unmonitored area in the Cape Breton Highlands. Can. J. Soil Sci. 1998, 78, 409–420. [Google Scholar] [CrossRef]
- Saatchi, S.; Buermann, W.; ter Steege, H.; Mori, S.; Smith, T.B. Modeling distribution of Amazonian tree species and diversity using remote sensing measurements. Remote Sens. Environ. 2008, 112, 2000–2017. [Google Scholar] [CrossRef]
- Zimmermann, N.E.; Moisen, G.G.; Edwards Jr., T.C.; Frescino, T.S.; Blackard, J.A. Remote sensing-based predictors improve distribution models of rare, early successional and broadleaf tree species in Utah. J. Appl. Ecol. 2007, 44, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.; Spector, S.; Gardiner, N.; Fladeland, M.; Sterling, E.; Steininger, M. Remote sensing for biodiversity science and conservation. Trends Ecol. Evol. 2003, 18, 306–314. [Google Scholar] [CrossRef]
- Hassan, Q.K.; Bourque, C.P.-A.; Meng, F.-R.; Richards, W. Spatial mapping of growing degree days: An application of MODIS-based surface temperatures and enhanced vegetation index. J. Appl. Remote Sens. 2007a, 1, 013511:1–013511:12. [Google Scholar] [CrossRef]
- Hassan, Q.K.; Bourque, C.P.-A.; Meng, F.-R.; Cox, R.M. A wetness index using terrain-corrected surface temperature and normalized difference vegetation index: An evaluation of its use in a humid forest-dominated region of eastern Canada. Sensors 2007b, 7, 2028–2048. [Google Scholar] [CrossRef]
- Godin, B.; Roberts, M.R. Part I. Ecological Land Classification for New Brunswick: The Ecoprovince, Ecoregion and Ecodistrict Levels [Unpublished report], On file with Provincial Legislative Library, Fredericton, New Brunswick, Canada, Mimeographed, 1994.
- Hassan, Q.K.; Bourque, C.P.-A.; Meng, F.-R. Estimation of daytime net ecosystem CO2 exchange over balsam fir forests in eastern Canada: Combining averaged tower-based flux measurements with remotely sensed MODIS data. Can. J. Remote Sens. 2006, 32, 405–416. [Google Scholar] [CrossRef]
- Hassan, Q.K.; Bourque, C.P.-A. Estimating daily evapotranspiration for forests in Altantic Maritime Canada: application of MODIS imagery. In Proceeding ASPRS, Reno, Nevada, USA, May 1-5, 2006. (CD-ROM unpaginated).
- Sabins, F.F. Remote Sensing Principles and Interpretation, 3rd ed.; W.H. Freeman and Company: New York, NY, USA, 1997. [Google Scholar]
- Short, N.M. Remote Sensing Tutorial. Available at: http://rst.gsfc.nasa.gov/ (website last accessed August 13, 2009).
- Shugart, H.H.; Smith, T.M. A review of forest patch models and their application to global change research. Clim. Change 1996, 34, 131–153. [Google Scholar] [CrossRef]
- Barden, L.S. The linear relation between stand yield and integrated light in a shade-adapted annual grass. B. Torrey Bot. Club 1996, 123, 122–125. [Google Scholar] [CrossRef]
- Smith, M.J. An Examination of Forest Succession in the Cape Breton Highlands of Nova Scotia. M.Sc. Thesis, University of New Brunswick, Fredericton, 1998. [Google Scholar]
- Acevedo, M.F.; Urban, D.L.; Ablan, M. Transition and gap models of forest dynamics. Ecol. Appl. 1995, 5, 1040–1055. [Google Scholar] [CrossRef]
- Taylor, S.L.; MacLean, D.A. Rate and causes of decline of mature and overmature balsam fir and spruce stands in New Brunswick, Canada. Can. J. Forest Res. 2005, 35, 2479–2490. [Google Scholar] [CrossRef]
- Austin, M.P.; Nicholls, A.O.; Doherty, M.D.; Meyers, J.A. Determining species response functions to an environmental gradient by means of a β-function. J. Veg. Sci. 1994, 5, 215–228. [Google Scholar] [CrossRef]
- Landsberg, J.J.; Waring, R.H.; Coops, N.C. Performance of the forest productivity model 3-PG applied to a wide range of forest types. Forest Ecol. Manage. 2003, 172, 199–214. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A.M. Potential colonization of newly available tree-species habitat under climate change: An analysis for five eastern US species. Landscape Ecol. 2004, 19, 787–799. [Google Scholar] [CrossRef]
- Bourque, C.P.-A.; Hassan, Q.K. Projected impacts of climate change on species distribution in the Acadian forest region of eastern Nova Scotia. Forest. Chron. 2008, 84, 553–557. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hassan, Q.K.; Bourque, C.P.-A. Potential Species Distribution of Balsam Fir Based on the Integration of Biophysical Variables Derived with Remote Sensing and Process-Based Methods. Remote Sens. 2009, 1, 393-407. https://doi.org/10.3390/rs1030393
Hassan QK, Bourque CP-A. Potential Species Distribution of Balsam Fir Based on the Integration of Biophysical Variables Derived with Remote Sensing and Process-Based Methods. Remote Sensing. 2009; 1(3):393-407. https://doi.org/10.3390/rs1030393
Chicago/Turabian StyleHassan, Quazi K., and Charles P.-A. Bourque. 2009. "Potential Species Distribution of Balsam Fir Based on the Integration of Biophysical Variables Derived with Remote Sensing and Process-Based Methods" Remote Sensing 1, no. 3: 393-407. https://doi.org/10.3390/rs1030393