1. Introduction
Phenology is the study of recurring life cycle stages, and their timing and relationship with environmental factors [
1,
2]. Phenology controls the seasonality of ecosystem functions and plant feedbacks to climate through diverse processes, such as changes in the surface albedo and the exchange of CO
2 between atmosphere and biosphere [
3,
4,
5,
6]. Despite its importance, phenology is not always well described in Earth system models [
7,
8,
9], in particular, the environmental factors controlling phenology are still uncertain [
6,
10]. Therefore, additional efforts to monitor and model plant phenology are needed to improve the representation of phenology in Earth system models [
6,
11].
Conventional visual monitoring of phenology dates back to 705 CE [
12], and still plays an important role in evaluating the impacts of climate change on ecosystems [
13,
14,
15,
16]. However, conventional monitoring requires substantial field work, which limits spatial and temporal representativeness [
17].
Near-surface remote sensing is becoming a more frequently used tool to monitor vegetation phenology at the ecosystem scale. In recent years, installation of commercial digital cameras for phenology monitoring (i.e., PhenoCam) has proliferated throughout diverse biomes and continents [
18,
19,
20,
21,
22,
23], which has led to the consolidation of national and continental monitoring networks [
24,
25,
26,
27,
28]. The use of PhenoCam consistently reduces manual labor, guarantees time series of high temporal resolution, and creates a permanent data record from which visual interpretation and qualification can be made at any later point in time [
29]. The proximity to the target ecosystem allows the cameras to track phenological transition dates (PTDs), such as leaf emergence, leaf discoloration, senescence, and green up and senescence of vegetation with high temporal resolution [
23,
30], as well as monitor the different plant types within the camera’s field of view (FOV; [
24]). Nowadays, the increasing number of sites with digital cameras co-located with ecosystem-atmosphere CO
2 flux measurements collected using the eddy covariance (EC) technique are contributing to understanding the relationship between phenology of structure and function of ecosystems [
28,
31,
32].
Green chromatic coordinates (GCC) is the most commonly used vegetation index (VI) extracted from PhenoCam, due to the requirement of only three visible spectral bands for computation, and it is used to represent plant development throughout the season [
33]. PhenoCam, with an additional near-infrared (NIR) band which is more sensitive to vegetation structural change than visible bands, has increased the use of the PhenoCam-based normalized difference vegetation index (CamNDVI) for the same purpose [
29,
34,
35]. Both GCC and CamNDVI are considered plausible indexes to bridge satellite and ground-based observations of phenology [
24]. For instance, GCC and CamNDVI have been shown to be effective (though not always consistent) tools for describing greenness variation of individual plant species and ecosystems in a variety of plant functional types [
29,
36], and for evaluating and linking remote sensing phenology products [
18,
37,
38,
39] with ground observations [
40].
Recently, Badgley et al. [
41] introduced a new vegetation index called near-infrared reflectance of vegetation index (NIRv), designed to mitigate the mixed pixel problem (determining the fraction of vegetated land surface and reconstructing the signal attributable to vegetation) to better represent photosynthesis of ecosystems. A strong correlation between satellite-based NIRv and gross primary productivity (GPP) at global scale was observed, which outperforms the correlation between NDVI and GPP [
40]. As such, it would be interesting to know if this new index could provide an advantage in tracking seasonal GPP and phenology compared to the widely used CamNDVI and GCC at ecosystem-scale. The computation of NIRv, as well as other VIs, such as the ratio vegetation index (RVI [
42]), is possible with NIR-enabled PhenoCam by following the approach proposed by Petach et al. [
29] and Filippa et al. [
34] for the computation of CamNDVI. However, up to now, we are not aware of studies evaluating the differences between PTDs derived from multiple PhenoCam-based VIs from PhenoCam (GCC, CamNDVI, CamNIRv, and CamRVI).
Past studies related to derived PTDs from PhenoCam mainly focus on temperate/boreal forests and grassland (e.g., [
32]), and only a few recent studies have focused on seasonally dry tree–grass ecosystems [
43,
44]. Considering that tree–grass ecosystems are a widely distributed land cover type, which occupies 16–35% of the Earth’s land surface [
45,
46,
47], it is necessary to further investigate the methods to extract PTDs for these ecosystems.
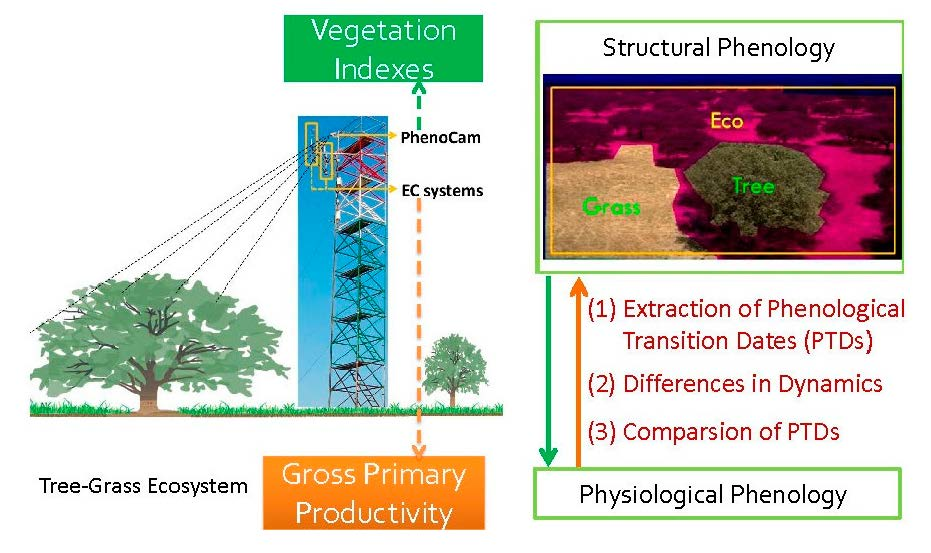
Moreover, the increasing number of sites with PhenoCam associated with EC flux measurements open interesting perspectives to evaluate: first, the consistency between PTDs derived from PhenoCam-based VIs and PTDs of ecosystem functioning (physiological phenology, i.e., [
48]); second, the direct relationship between PhenoCam-based VIs and GPP. However, to our knowledge, only a few studies pay special attention to the differences between phenology of ecosystem structure and of ecosystem functioning and carbon fluxes [
28,
32].
In this study, our main objective is to evaluate the potential of PhenoCam to monitor phenology of seasonally dry Mediterranean tree–grass ecosystems. Specifically, the objectives are (1) to characterize structural and physiological phenology of tree–grass ecosystems and their main climatic drivers using PhenoCam and GPP derived from EC measurements; (2) to compare the PTDs and growing season length (GSL) derived from different PhenoCam-based VIs, and to evaluate their performance in tracking the PTDs and GSL derived from GPP.
2. Materials and Methods
2.1. Sites Description, Instrument Set-Up, and Data Sources
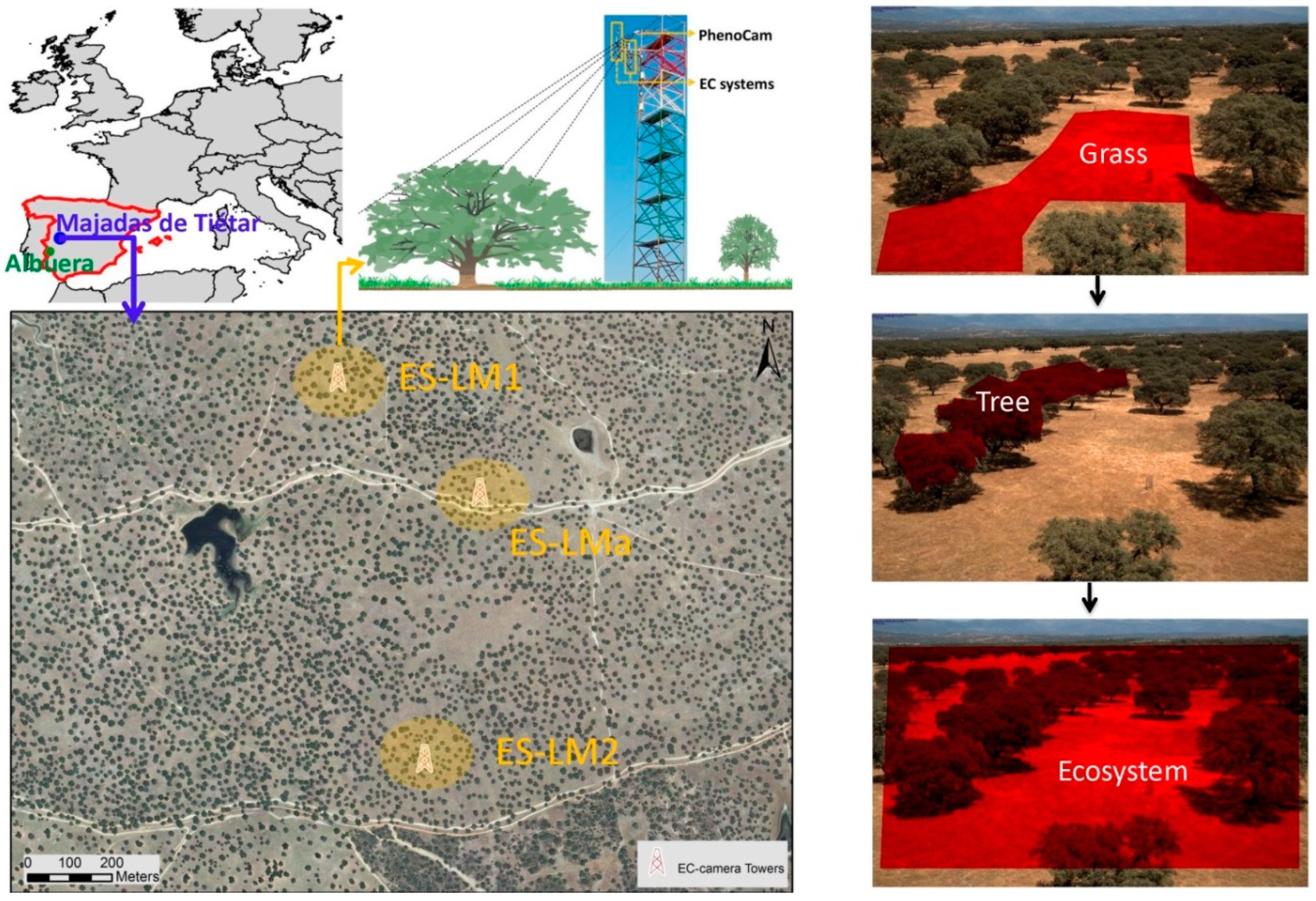
The sites used in this study are Mediterranean tree–grass ecosystems, composed predominantly of an herbaceous layer and low-density evergreen broadleaf oak trees (Quercus ilex; ~20 tree ha
−1;
Figure 1). Three sites are located approximately 500 m apart from each other in Majadas de Tiétar, Cáceres, Spain (39°56′24.68″N, 5°46′28.70″W), while one site is located in La Albuera, Spain (38°42′6.48″N, 6°47′9.24″W). The experimental sites in Majadas de Tiétar belong to a large-scale manipulation experiment, where the three areas of approximately 20 ha were manipulated with addition of nitrogen (FLUXNET ID ES-LM1), nitrogen and phosphorous (FLUXNET ID ES-LM2), and the last was kept as control (FLUXNET ID ES-LMa). The experimental site in the La Albuera (FLUXNET ID ES-Abr) is a natural ecosystem with no manipulation. In this study, we did not focus on the fertilization, but only on the evaluation of the effectiveness of different vegetation indexes to represent the ecosystem functions. The Majadas de Tiétar and La Albuera are characterized by a long-term annual mean air temperature of 16.7 ± 0.2 °C and 18.3 ± 1.5 °C, respectively; while mean annual rainfall is ca. 650 mm and 400 mm, respectively. The rain falls typically from November to May with a very dry summer [
49].
In each site, an EC system was installed at 15 m of height to measure the carbon, water and energy fluxes (
Section 2.2 for more details). The fluxes data are available from March 2014 in ES-LM1, ES-LM2, and ES-LMa; and from October 2015 for ES-Abr. Two broadband Decagon SRS (spectral reflectance sensor) sensors with a FOV of 36 degrees were installed on a rotating arm in each tower area. Downwelling irradiance and upwelling radiance at 650 nm (red spectral band) and 810 nm (near-infrared spectral band) were measured every 5 min for tree and grasses from 30 October 2015.
A NIR-enabled digital camera (Stardot NetCam 5MP), was mounted at the top of the EC tower (facing north) at each site. Images were collected every 30 min (from 10:00 to 14:30 UTC) as JPEG format. The camera settings were defined according to the “PhenoCam” protocol (
https://phenocam.sr.unh.edu/webcam/tools/). Sequential red, blue, green (RGB) and RGB + NIR images were collected by the Stardot camera according to Petach et al., [
29]. FOVs of cameras in ES-LM1 and ES-LM2 were stable during the study period (from 1 August 2014 to 31 July 2017), whereas the FOV of ES-LMa was not constant and the camera experienced a white balance problem until 3 December 2015. At the ES-Abr site, images were available from 1 January 2016. Hence, RGB and RGB+NIR images were available for the analysis from 1 August 2014 to 31 July 2017 for ES-LM1 and ES-LM2; from 3 December 2015 to 31 July 2017 for the ES-LMa site; and from 1 January 2016 to 31 July 2017 for the ES-Abr site. This guarantees a total of 9 site-years for the following analysis.
2.2. EC Data Processing and Flux Partitioning to GPP
Each EC system consists of a three-dimensional sonic anemometer (R3-50, Gill LTD, Lymington UK) and an infrared gas analyzer (LI-7200, Licor Bioscience, Lincoln, NE, USA) to measure mixing ratios of CO2 and H2O. Additional vertical CO2 and H2O concentration profiles were measured at seven levels between the surface and the measurement height in the EC tower (0.1, 0.5, 1.0, 2.0, 5.0, 9.0, and 15 m above ground with a LI840, Licor Bioscience, Lincoln, NE, USA). Meteorological variables such as air temperature (Ta), wind speed (WS), relative humidity (RH), incoming global radiation (Rg), photosynthetically active radiation (PAR), and precipitation (Prec) were also measured at each site.
Raw EC data were collected at 20 Hz, and were then processed using EddyPro 6.2. The main processing procedures for CO
2 fluxes included (1) coordinate rotation using planar fit method [
50]; (2) CO
2 time lag adjustments by covariance maximization in predefined windows; (3) spectral corrections performed for low and high pass-filtering effects according to Moncrieff et al. [
51] and Moncrieff et al. [
52]. The calculated flux for CO
2 was then quality checked [
53,
54]. The net ecosystem exchange (NEE) flux was corrected by adding storage fluxes (integrated CO
2 fluxes using seven levels of CO
2 profiles when possible, otherwise using 1-point storage) to CO
2 flux.
The u*-threshold, which was used as a criterion to discriminate low- and well-mixed eddies in the nighttime, was estimated for each year and tower individually (the median of u*-threshold ranges from 0.11 to 0.18 for ES-LM1, ES-LM2, and ES-LMa while 0.20–0.24 for ES-Abr) following Papale et al. [
55].
The time series of NEE were gap filled using the marginal distribution sampling (MDS) method [
56] which is based on lookup tables of temperature, global radiation, and water vapor pressure deficit (VPD) classes for short temporal windows i.e., 14 days. The gap-filled time series of NEE were then partitioned into GPP as described in Reichstein et al. [
56]. In brief, the nighttime flux (Rg < 10 W/m
2) of NEE (i.e., only respiration) is extrapolated from nighttime to daytime through a temperature response function, which is based on short term temperature sensitivities (for details see [
56]). The u*-threshold, gap-filling, and partitioning was performed with the R package REddyproc [
57].
2.3. Calculation of Vegetation Indexes from PhenoCam
Digital numbers (DNs) of each individual channel (R
DN, G
DN, B
DN and NIR
DN) were extracted from each photograph, and averaged over the different regions of interest (ROIs) (
Figure 1). The overall brightness of each ROI (RGB
DN) and the relative brightness of green channel, known also as green chromatic coordinates—GCC, were computed with Equations (1) and (2):
CamNDVI was also computed in the different ROIs according to Petach et al. [
29], using the algorithm implemented in the “phenopix” R package [
34,
36]:
where NIR
DN‘ and R
DN’ are the adjusted exposure values of NIR
DN and R
DN, respectively. For a detailed calculation and the exposure adjustment formula, please refer to Petach et al. [
29]. As the R
DN’ and NIR
DN’ are not direct measurements of reflectance, the CamNDVI values are not directly comparable to the NDVI from other data sources. Petach et al. [
29] found a linear relationship between CamNDVI and the NDVI derived from the radiometric sensor (ASD FieldSpec 3) using the bands of 750 nm for the NIR and 605 nm for the red. They suggested using the linear regression coefficients to adjust the CamNDVI values for comparability with NDVI from radiometers [
29,
34].
Therefore, we applied the method suggested by Petach et al. [
29] and Filippa et al. [
34] to rescale CamNDVI using the NDVI derived from the Decagon SRS (VIs was calculated and averaged over a 30 min period to be consistent with VIs from PhenoCam). The coefficients and statistics of the linear regression used to compute the scaling factors are shown in
Table 1. In the following only the rescaled CamNDVI values are used and presented.
Likewise, the CamNIRv and CamRVI were also calculated with adopting Equations (4) and (5) which refer to Badgley et al. [
41] and Chen [
42], respectively.
where NIR
DN′ and R
DN′ are the adjusted exposure values like Equation (3). A similar approach used for the CamNDVI was used to compute CamNIRv and CamRVI: SRS-based NIRv and RVI were used to adjust the CamNIRv and CamRVI in order to make them comparable with the data derived from other sources (
Table 1).
The analysis was conducted on various ROIs as depicted in
Figure 1: we selected ROIs with only trees, grass, and both (hereafter referred as Tree, Grass, and Eco ROI, respectively). The different sites have different tree/grass proportions in the camera FOVs, as only one direction of ecosystems could be captured from the PhenoCam. However, the fractional tree canopy covers were consistent (~0.20) in the four sites by referring to field surveys and the classification analysis using airborne hyperspectral imagery [
58,
59]. The analysis of each footprint for each EC tower also indicates GPP is contributed from ~20% tree canopy and ~80% grasses [
59]. In order to reduce the bias introduced by the different ratios of tree/grass in the images and camera FOV, which has to do with logistical constrains during the camera installation, the ecosystem VIs (GCC, CamNDVI, CamNIRv, CamRVI) were computed by using the weighted average of the VIs derived from Grass ROIs of 0.8 and Tree ROIs of 0.2.
2.4. Data Filtering and to Compute Daily VIs and GPP
After computing the half-hourly VIs (GCC, CamNDVI, CamNIRv, CamRVI) at ecosystem scale, we applied a series of steps to derive robust time series of daily VIs:
2.5. Phenological Transition Dates (PTDs) Extraction
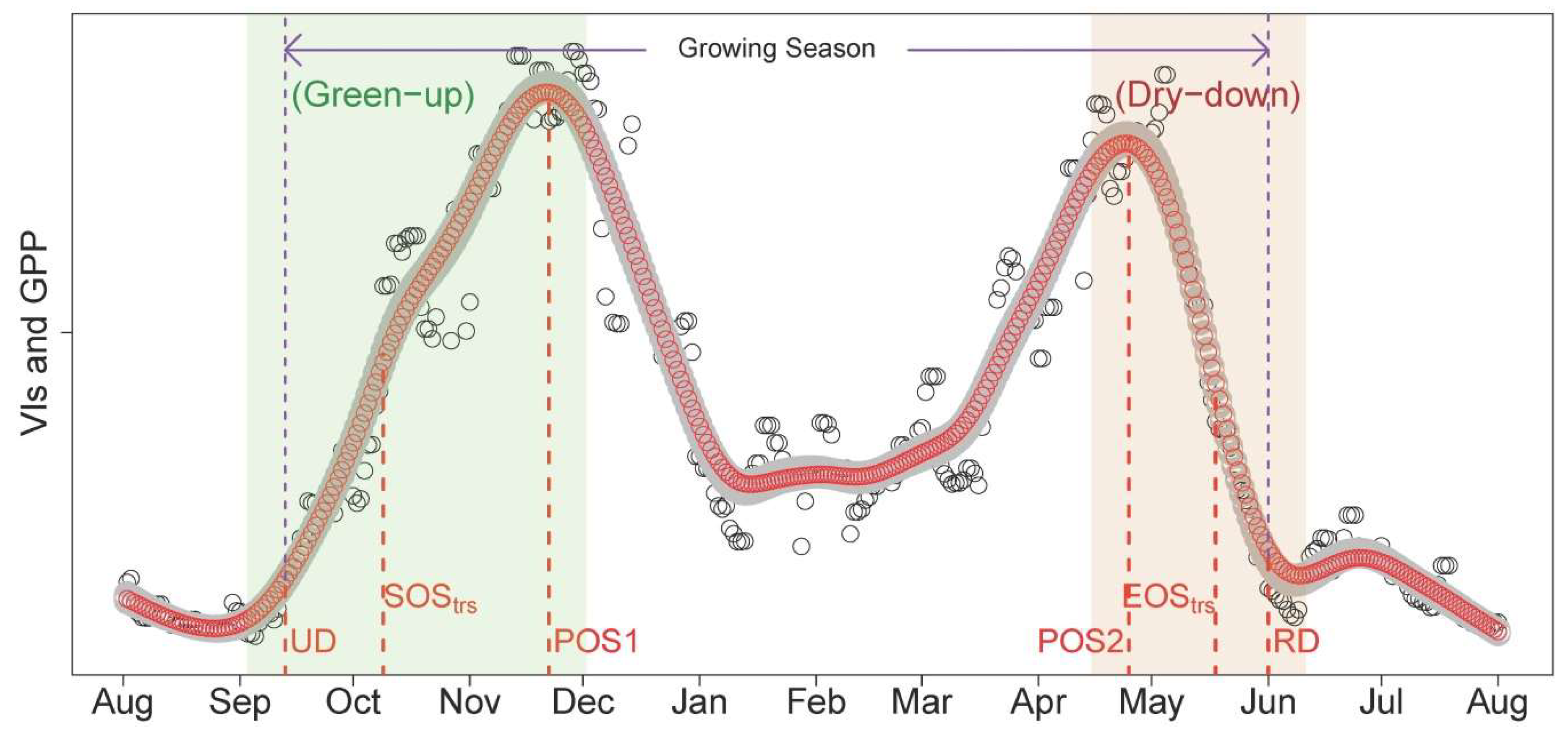
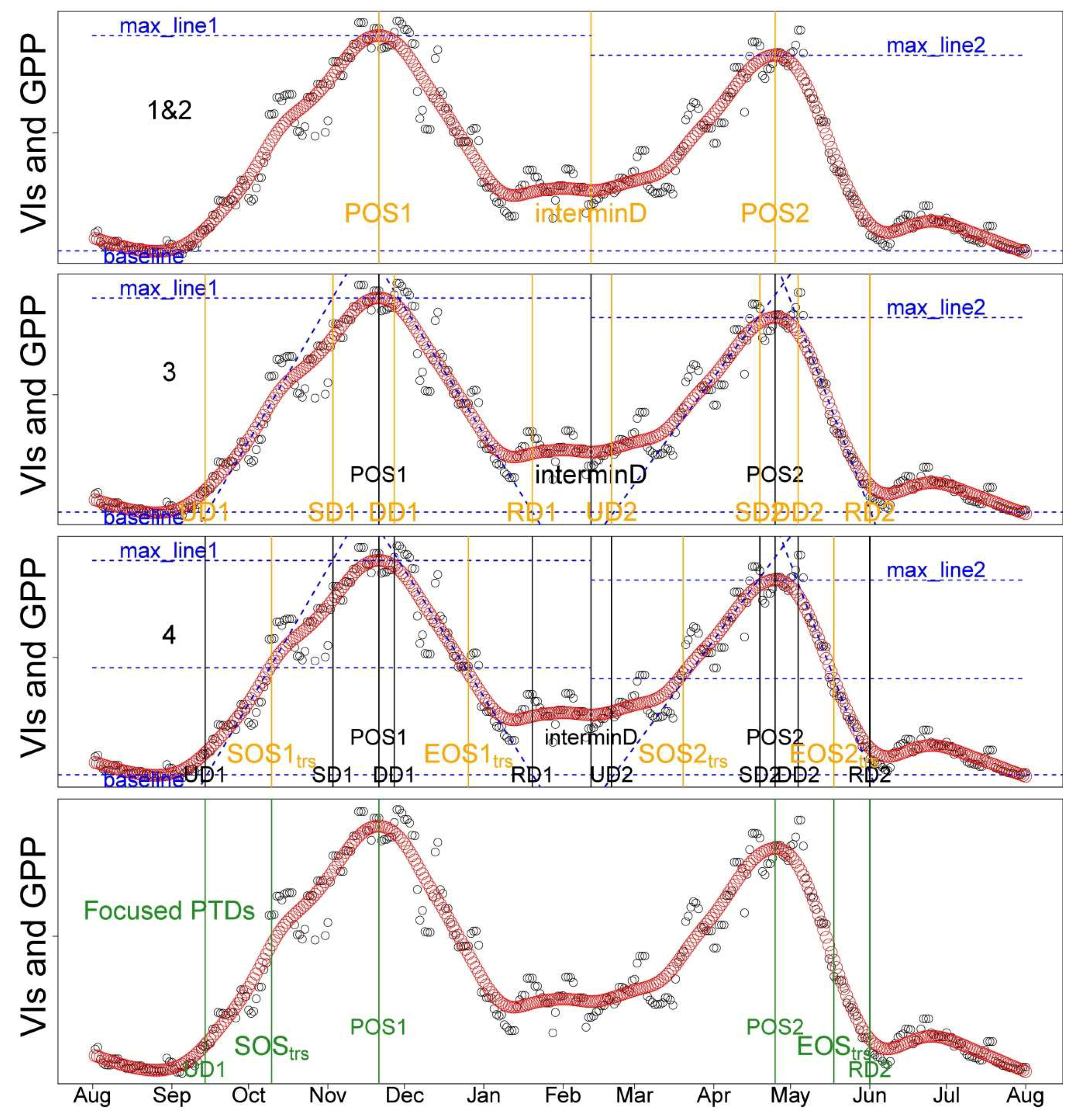
The Mediterranean climate is characterized by rainy late autumn–winters, and warm dry summers. Typically, the studied tree–grass ecosystems are dry and covered with senescent grasses in summer, while they increase in greenness in the late autumn (after the onset of the rainy season) and spring. Considering the characteristics of the phenological cycle described above, we decided to conduct the analysis using the concept of “hydrological years”, which is, here, defined from 1 August to 31 July (
Figure 2).
In this study, we developed a PTD extraction method for PhenoCam-based VIs in seasonally dry tree–grass ecosystems. The methodology of PTDs extraction is composed by the following steps:
Data were smoothed using the spline method [
20,
36]; PTDs were extracted using the derivatives of smoothed seasonal cycle [
61] and applying thresholds (i.e., 50%) of amplitude of VIs [
62]. As the start and end of the season are extremely important to characterize the phenology, we defined two sets of PTDs in the start (UD, SOS
trs;
Table 2) and end of season (RD, EOS
trs;
Table 2) for intercomparison and better characterizing the phenology. These two sets of PTDs are derived based on different perception and methodology. UD and RD are retrieved as the intersection dates between steepest slope and minimum value in the Green-up and Dry-down periods, respectively [
61]. In contrast, SOS
trs and EOS
trs are retrieved by using the thresholds of 50% amplitude [
62]; i.e., they are defined when 50% of amplitudes are reached in the Green-up and Dry-down periods, respectively. Other extracted PTDs and the phenological periods analyzed in this study were summarized in
Figure 2 and
Table 2. The detailed procedures and corresponding code related to the extraction of PTDs are provided in
Appendix B.
Uncertainty of extracted PTDs was assessed by extracting PTDs repeatedly (100 times) from an ensemble of time series constructed by summing original data and random noise as described by Filippa et al. [
36].
2.6. Statistical Analysis
All the statistical analyses were conducted with the R 3.4.3 programming language [
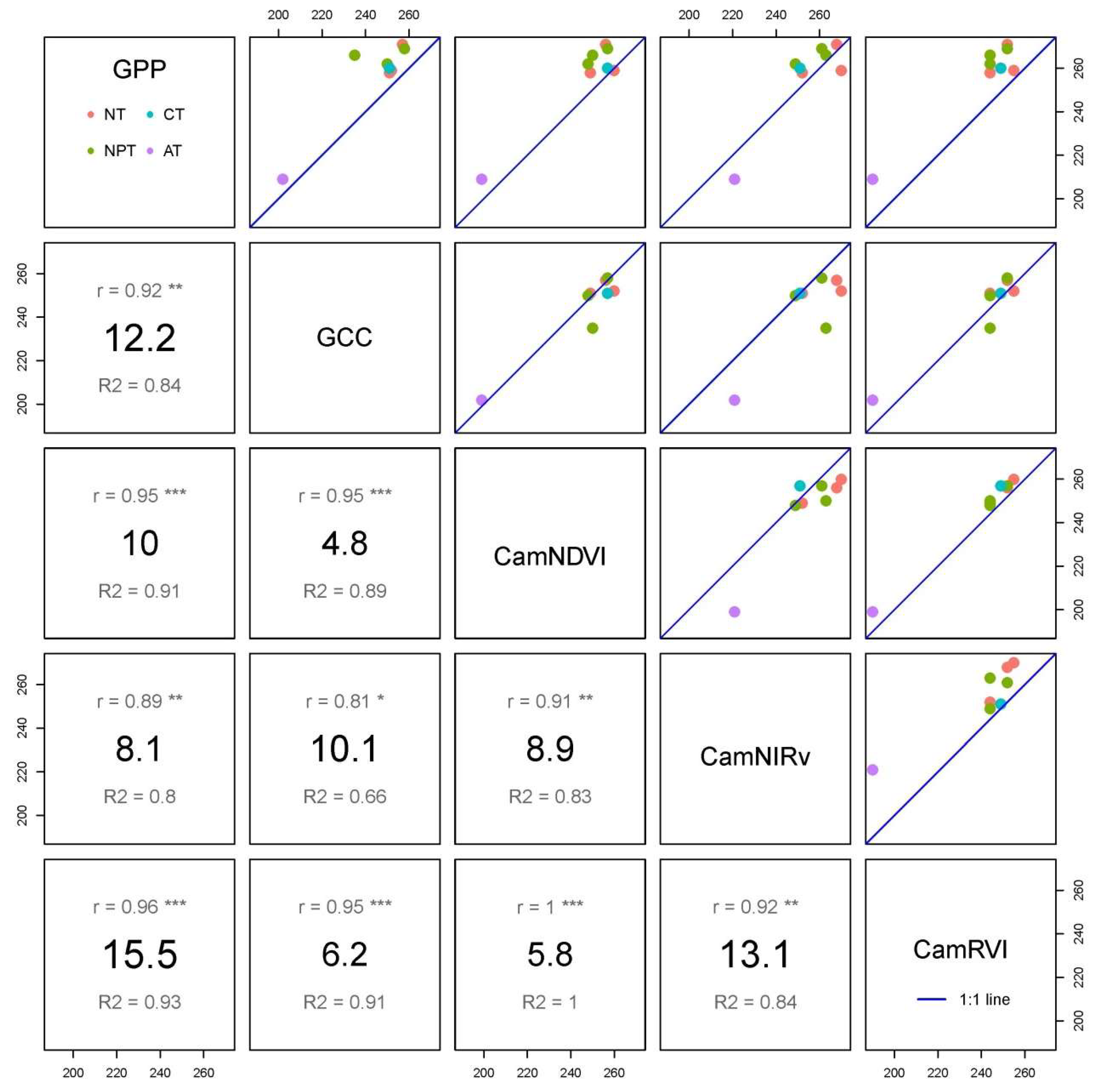
63]. The differences among PTDs extracted from the different datasets (GCC, CamNDVI, CamNIRv, CamRVI, and GPP) were evaluated using the mean absolute error (MAE) and root mean squared error (RMSE) (Equations (6) and (7)):
where y
i’ and y
i were the PTD dates extracted from two different datasets. Wilcoxon signed-rank tests were used to test for statistically significant differences between each paired PTDs from the different datasets given that were not normally distributed, while paired Student’s
t tests were used when PTDs were normally distributed.
The linear regressions were conducted between time series of VIs and GPP, or between meteorological variables and GPP using ordinary least squares regression (OLS). However, the linear regression between PTDs and GSL extracted from different VIs and GPP was conducted using major axis regression (R package “lmodel2”) to account for errors of similar magnitude in the y and x axis.
5. Conclusions
In this study, we assessed the potential to jointly use near-infrared-enabled digital repeat photography and eddy covariance data for monitoring structural and physiological phenology in seasonally dry Mediterranean tree–grass ecosystems. We analyzed 9 site-years using four PhenoCam-based vegetation indices (GCC, CamNDVI, CamNIRv, and CamRVI) and GPP, and we compared the phenological transition dates (PTDs) and growing season length (GSL) derived from the different data streams.
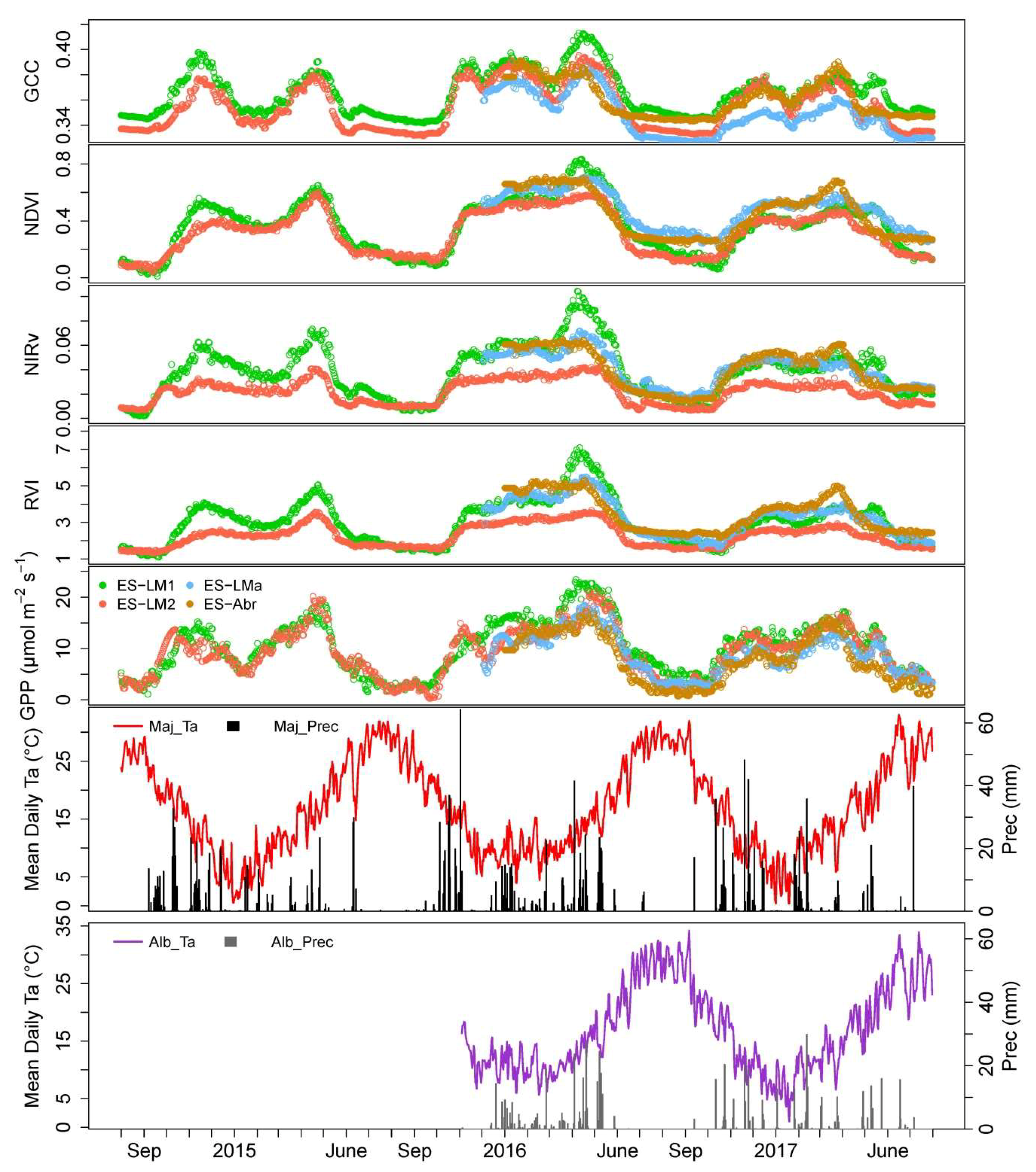
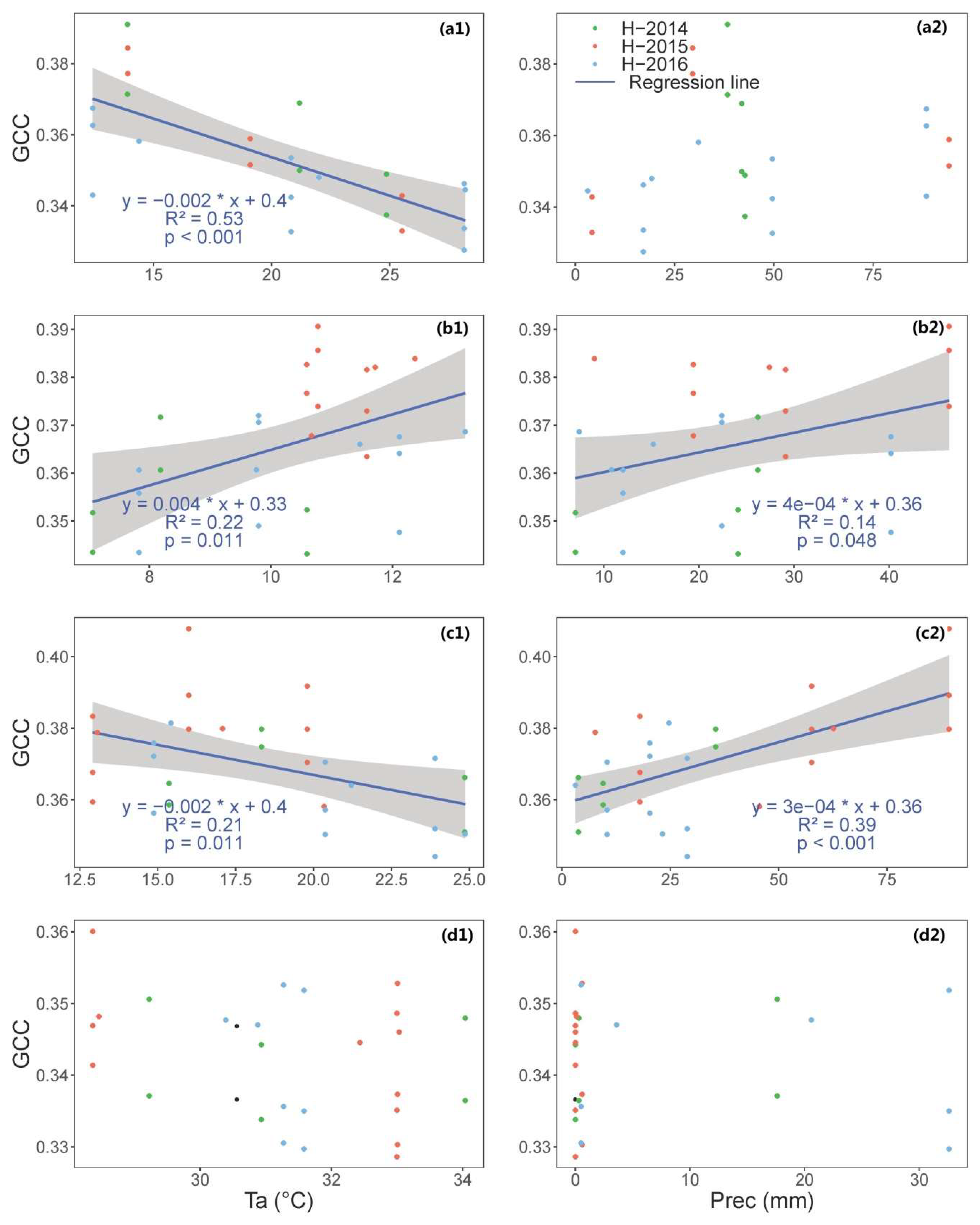
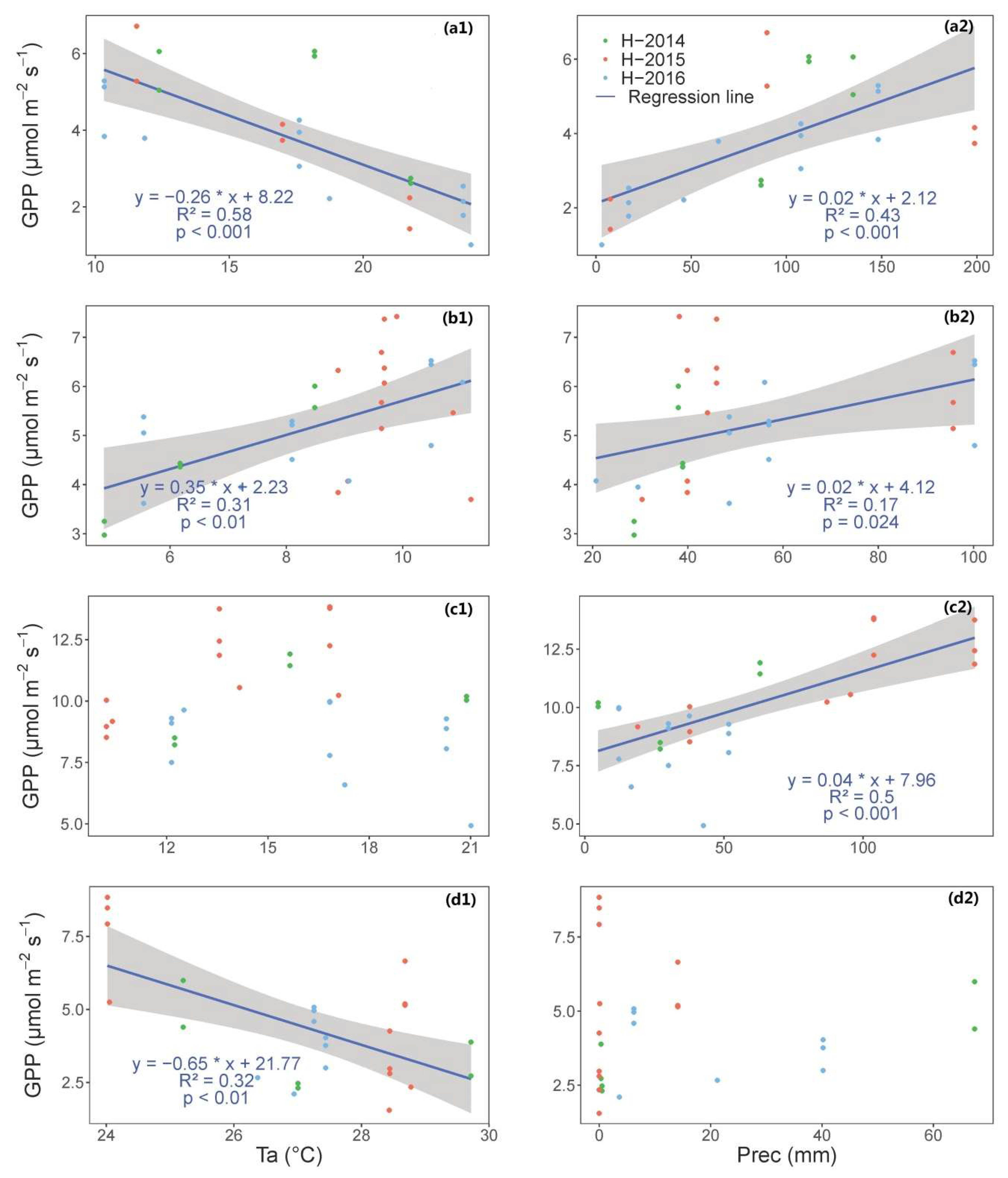
We show that, in Mediterranean tree–grass ecosystem, meteorology plays an important role in governing seasonal variation of vegetation indices and GPP, though the importance of water availability and temperature vary across seasons.
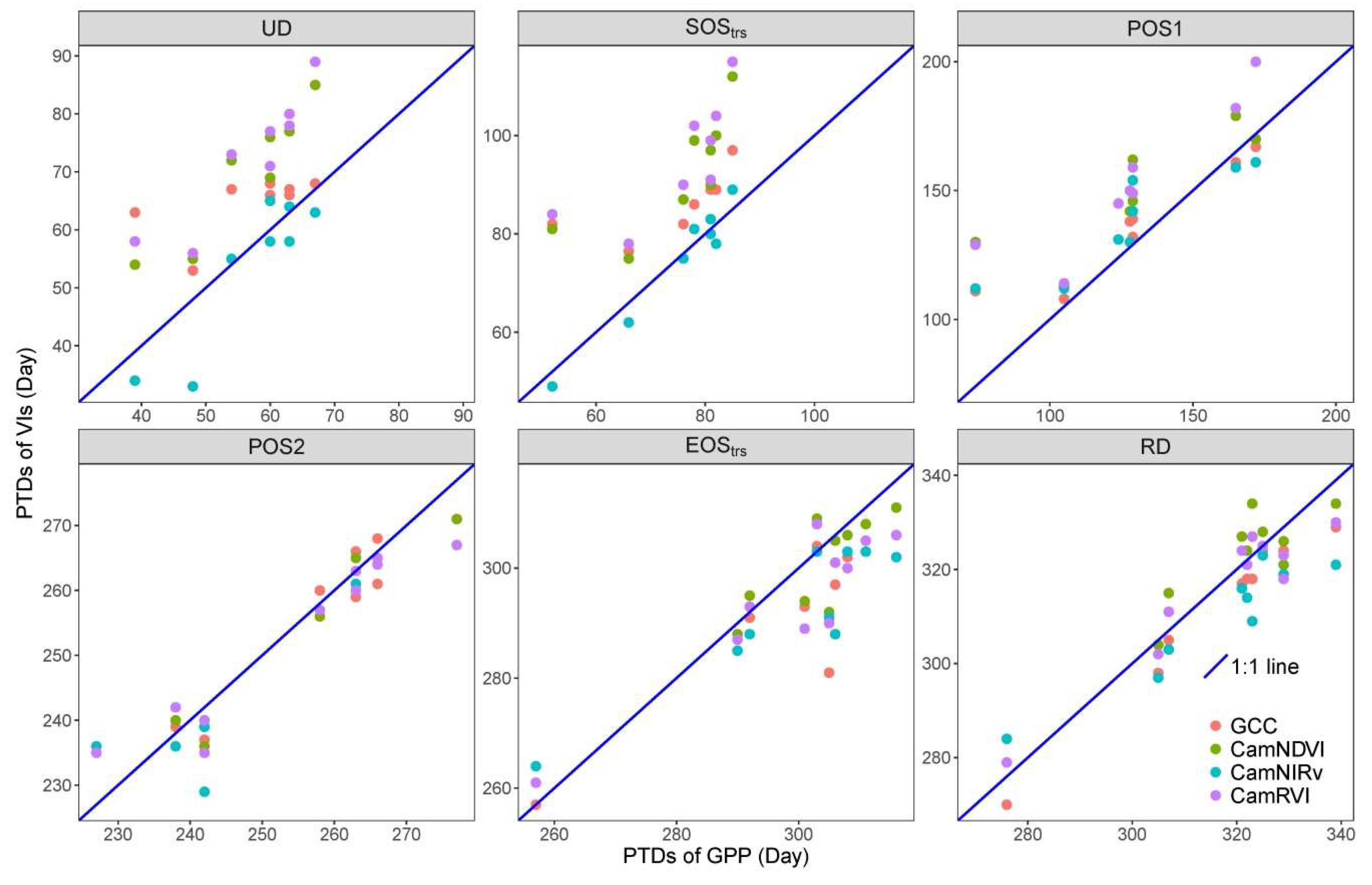
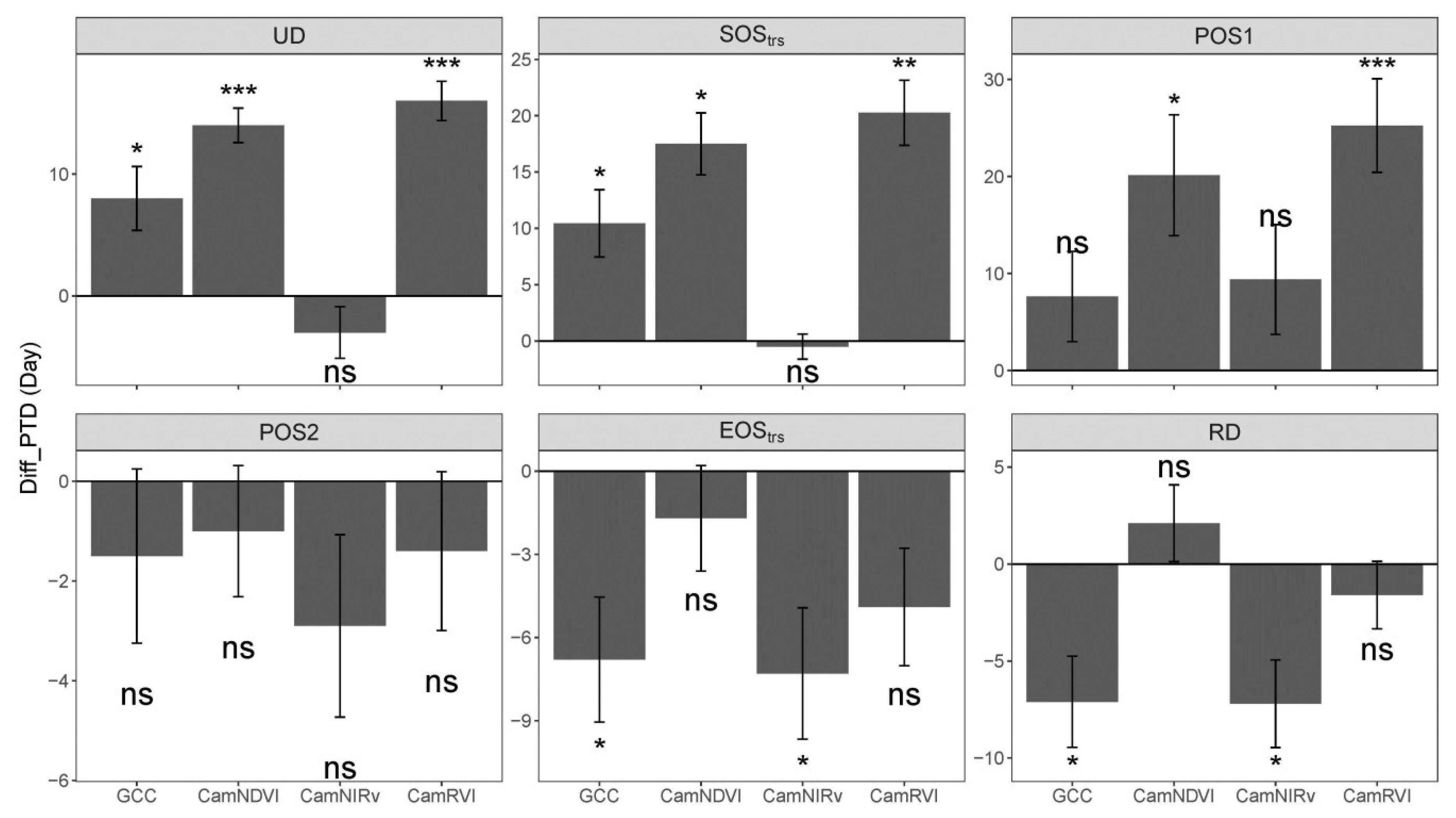
We show the PTDs derived from VIs differ from each other. For the widely used GCC and CamNDVI, the PTDs extracted from CamNDVI are delayed compared to the ones derived from the GCC, which is likely attributed to GCC, and is more sensitive to color changes, while CamNDVI is more sensitive to LAI and biomass.
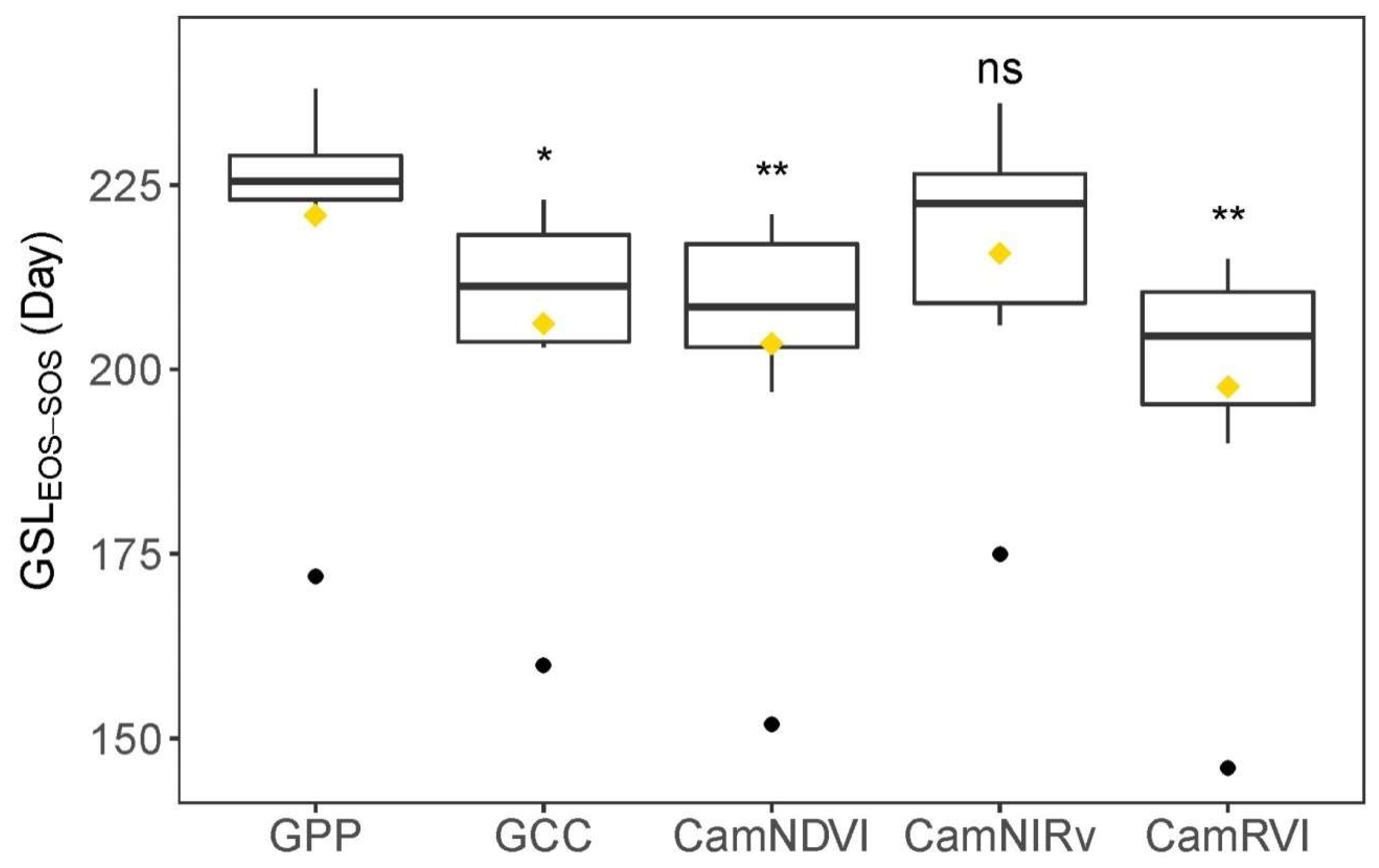
CamNIRv is best at representing the PTDs of GPP at the Green-up period, while CamNDVI is the best proxy to represent the PTDs of GPP at the Dry-down period. CamNIRv performs best regarding the representation of the GSL of GPP.
In summary, we show that it is possible to determine crucial PTDs of structural and physiological phenology through using near-infrared-enabled digital cameras. GPP could be well represented when combining the use of different VIs for this purpose.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}














