Hunting Activity Among Naturalistically Housed Chimpanzees (Pan troglodytes) at the Fundació Mona (Girona, Spain). Predation, Occasional Consumption and Strategies in Rehabilitated Animals


Abstract
:Simple Summary:
Abstract:
1. Introduction
2. Material and Methods
2.1. Study Subjects and Facilities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year arrived at Mona Foundation | Name | Sex | Birth date | Birth place | Background | Group | Participation in hunting |
|---|---|---|---|---|---|---|---|
| 2009 | África | Female | 2000 | Wild | Pet | B | YES |
| 2002 | Bongo | Male | 2000 | Captivity | Circus | B/A | YES |
| 2001 | Charly | Male | 1989 | Captivity | Circus, TV | A | YES |
| 2003 | Juanito | Male | 2003 | Captivity | Pet | B | YES |
| 2001 | Marco | Male | 1984 | Captivity | Circus, TV | A | YES |
| 2004 | Nico | Male | 2001 | Captivity | Pet | B | YES |
| 2001 | Romie | Female | 1979 | Wild | Breeding, Circus | B | YES |
| 2004 | Sara | Female | 1998 | Captivity | Pet, TV | B | YES |
| 2005 | Tico | Male | 1985 | Captivity | Pet, Zoo | B | NO |
| 2001 | Toni | Male | 1983 | Wild | TV, Zoo | A | NO |
| 2003 | Toto | Male | 1956 | Wild | Pet, Zoo | B | YES |
| 2006 | Víctor | Male | 1982 | Wild | Pet, Zoo | B | NO |
| 2002 | Waty | Female | 1996 | Captivity | Circus | B | YES |
2.2. Behavioral Observations
3. Results
| ID | Date | Species | Social/Individual | N° individuals involved | Predatory strategy | Prey consumption | System record | Chimpanzee group |
|---|---|---|---|---|---|---|---|---|
| H1 | 2007-01-19 | Oryctolagus cuniculus | Social | 4 | Coordinated | Try | Video | B |
| H2 | 2008-05-24 | Turdus merula | Individual | 1 | Individual | Yes | Photo | B |
| H3 | 2008-08-02 | Larus michaellis | Social | 3 | Individual | Try | Video and photo | A |
| H4 | 2010-10-29 | Oryctolagus cuniculus | Individual | 2 | Individual | Yes | Video and photo | A |
| H5 | 2011-09-17 | Oryctolagus cuniculus | Individual | 1 | Individual | Try | Photo | B |
| Species | Common name | Evidence of hunting | Chimpanzee group |
|---|---|---|---|
| Passer domesticus | House sparrow | Post-kill | A/B |
| Luscinia megarhynchos | Nightingale | No | |
| Ardea cinerea | Grey Heron | No | |
| Erithacus rubecula | European robin | No | |
| Larus michahellis | Yellow-legged Gull | Kill | A |
| Milvus migrans | Black kite | No | |
| Upupa epops | Hoopoe | No | |
| Turdus merula | Common Blackbird | Kill | B |
| Pica pica | European magpie | No | |
| Falco tinnunculus | Common Kestrel | No | |
| Erinaceus europaeus | European hedgehog | Post-kill | A/B |
| Oryctolagus cuniculus | European rabbit | Kill | A/B |
| Apodemus sylvaticus | Wood mouse | Post-kill | A/B |
| Rattus norvegicus | Brown rat | No | |
| Talpa europaea | European mole | Post-kill | A/B |
| Natrix natrix | Grass snake | Post-kill | A |
| Rhinechis scalaris | Ladder snake | Post-kill | A |
| Hyla meridionalis | Mediterranean tree frog | Post-kill | A/B |
| Rana perezi | Pere's frog | Post-kill | A/B |
| Bufo bufo | Common toad | Post-kill | A/B |
| Podarcis hispanica | Iberian wall lizard | Post-kill | A/B |
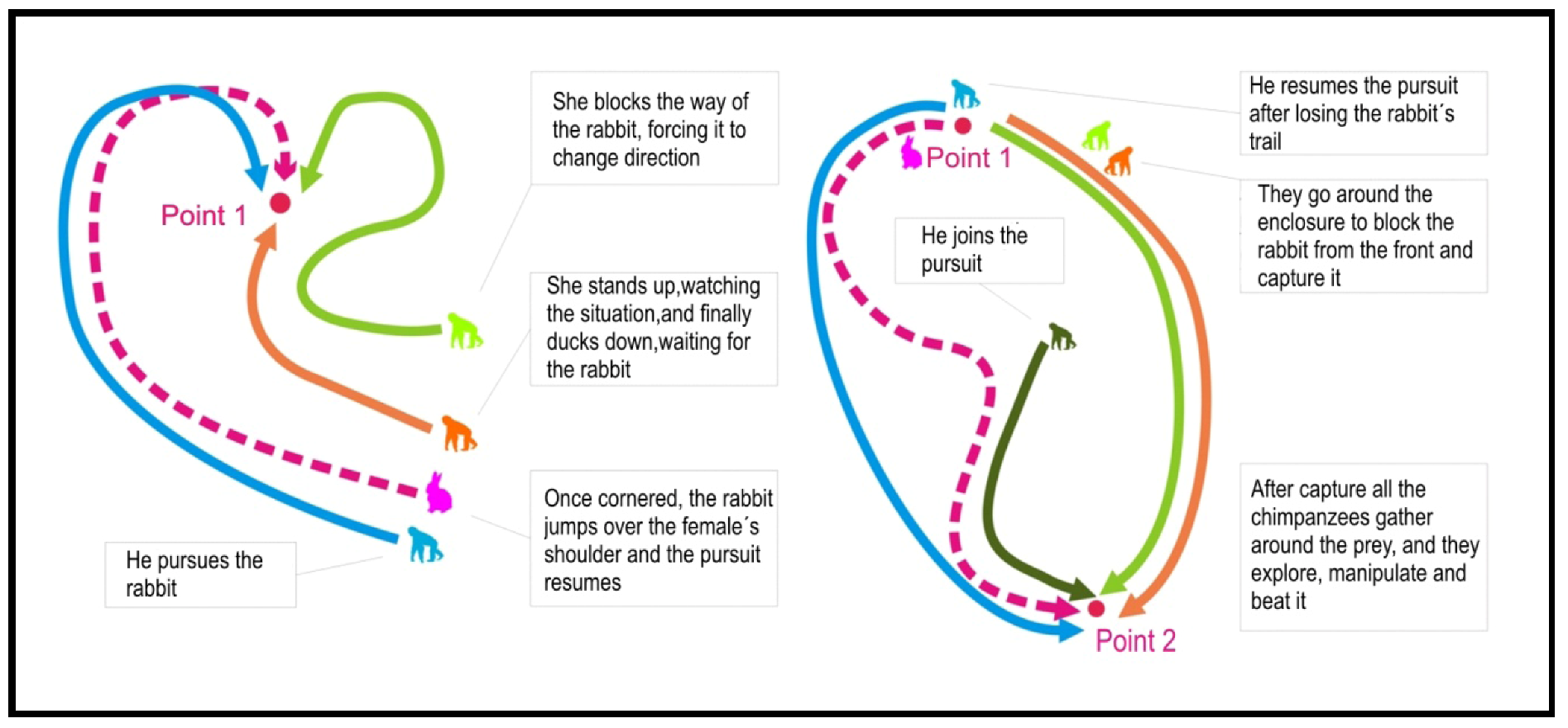
3.1. Case 1 (H1)—19 January 2007. Social Predation of a Rabbit (Oryctolagus cuniculus)
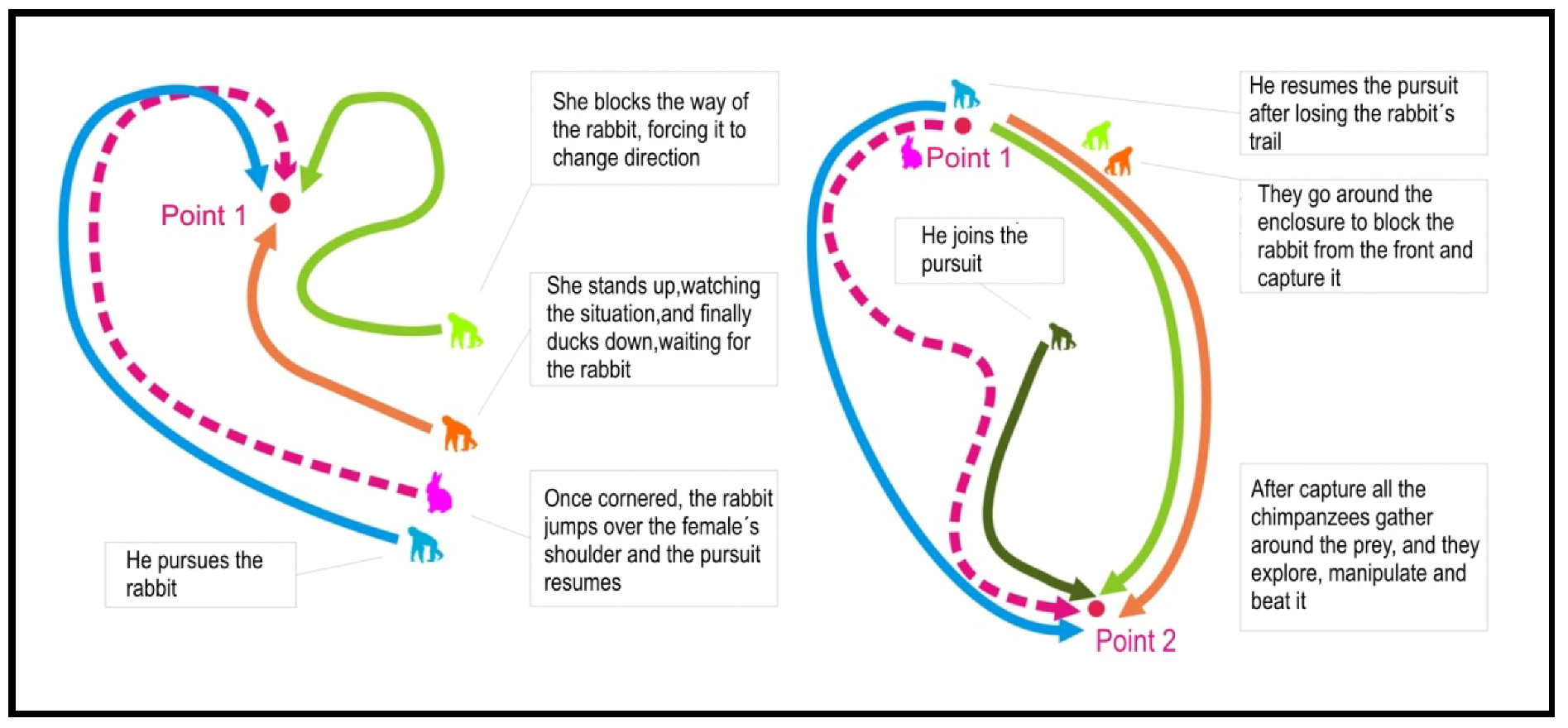

3.2. Case 2 (H2)—24 May 2008. Individual Predation of a Common Blackbird (Turdus merula)

3.3. Case 3 (H3)—2 August 2008. Individual Predation of a Yellow-Legged Gull (Larus michahellis)

3.4. Case 4 (H4)—29 October 2010. Individual Predation of a Rabbit (Oryctolagus cuniculus)

3.5. Case 5 (H5)—17 September 2011. Individual Predation of a Rabbit (Oryctolagus cuniculus)


4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Stanford, C.B. The Hunting Apes; Princeton University Press: Princeton, NJ, USA, 1999; p. 262. [Google Scholar]
- Stanford, C.B. A comparison of social meat-foraging by chimpanzees and human foragers. In Meat-Eating & Human Evolution; Stanford, C.B., Bunn, H.T., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 122–140. [Google Scholar]
- Boesch, C.; Boesch, H. Hunting behavior of wild chimpanzees in the Taï National Park. Am. J. Phys. Anthropol. 1989, 78, 547–573. [Google Scholar] [CrossRef]
- Boesch, C. Cooperative hunting in wild chimpanzees. Anim. Behav. 1994, 48, 653–667. [Google Scholar] [CrossRef]
- Teleki, G. The Predatory Behavior of Wild Chimpanzees; Bucknell University Press: Lewisburg, PA, USA, 1973; p. 232. [Google Scholar]
- Kuroda, S.; Suzuki, S.; Nishihara, T. Preliminary report on predatory behavior and meat sharing in Tschego chimpanzees (Pan troglodytes troglodytes) in the Ndoki Forest, Northern Congo. Primates 1996, 37, 253–259. [Google Scholar] [CrossRef]
- Gilby, I.C.; Wrangham, R.W. Risk-prone hunting by chimpanzees (Pan troglodytes schweinfurthii) increases during periods of high diet quality. Behav. Ecol. Sociobiol. 2007, 61, 1771–1779. [Google Scholar] [CrossRef]
- McGrew, W.C. Chimpanzee Material Culture. Implications for Human Evolution; Cambridge University Press: Cambridge, UK, 1992; p. 294. [Google Scholar]
- Stanford, C.B.; Wallis, J.; Matama, H.; Goodall, J. Patterns of predation by chimpanzees on red colobus monkeys in Gombe National Park, 1982–1991. Am. J. Phys. Anthropol. 1994, 94, 213–228. [Google Scholar] [CrossRef]
- Uehara, S. Predation on mammals by chimpanzee (Pan troglodytes). Primates 1997, 38, 193–214. [Google Scholar] [CrossRef]
- Teelen, S. Influence of chimpanzee predation on the red colobus population at Ngogo, Kibale National Park, Uganda. Primates 2008, 49, 41–49. [Google Scholar] [CrossRef]
- Boesch, C. Comparative hunting roles among Taï chimpanzees. Hum. Nature 2002, 13, 27–46. [Google Scholar] [CrossRef]
- Stanford, C.B. Chimpanzee and the Red Colobus. The Ecology of Predator and Prey; Harvard University Press: Cambridge, MA, USA, 1998; p. 336. [Google Scholar]
- O’Malley, R.C. Two observations of Galago predation by the Kasakela Chimpanzees of Gombe Stream National Park, Tanzania. Pan-Africa News 2010, 17, 17–19. [Google Scholar]
- Nishie, H. Increased hunting of Yellow Baboons (Papio cynocephalus) by M Group Chimpanzees at Mahale. Pan-Africa News 2004, 11, 10–12. [Google Scholar]
- Pruetz, J.D.; Marshack, J.L. Savanna chimpanzees (Pan troglodytes verus) prey on patas monkeys (Erythrocebus patas) at Fongoli, Senegal. Pan-Africa News 2009, 16, 15–17. [Google Scholar]
- Pruetz, J.D.; Bertolani, P. Savanna chimpanzees, Pan troglodytes verus, hunt with tools. Curr. Biol. 2007, 17, 412–417. [Google Scholar] [CrossRef]
- Nakamura, M.; Itoh, N. Hunting with tools by Mahale chimpanzees. Pan-Africa News 2008, 15, 3–6. [Google Scholar] [Green Version]
- Clarke, S.; Mitchell, G. Primate predation report: Survey results. Int. Zoo News 1983, 30, 4–8. [Google Scholar]
- Caine, N.; Vanovitz, C.; VanTassell, J.; Yee, K.; Mitchell, G. Predatory behavior in a captive rhesus monkey. Lab. Primate Newslett. 1979, 18, 25–26. [Google Scholar]
- Videan, E.N.; Fritz, J.; Murphy, J. Hunting and occasional consumption of prey items by chimpanzees at the Primate Foundation of Arizona. Int. J. Primatol. 2007, 28, 477–481. [Google Scholar] [CrossRef]
- Ross, S.R.; Lonsdorf, E.V. Interactions between zoo-housed great apes and local wildlife. Am. J. Primatol. 2009, 71, 458–465. [Google Scholar] [CrossRef]
- Llorente, M.; Mosquera, M.; Fabré, M. Manual laterality for simple reaching and bimanual coordinated task in naturalistic housed chimpanzees (Pan troglodytes). Int. J. Primatol. 2009, 30, 183–197. [Google Scholar] [CrossRef]
- Llorente, M.; Mosquera, M.; Fabré, M. Handedness for simple reaching and bimanual coordinated tasks in naturalistic housed chimpanzees (Pan troglodytes). Folia Primatol. 2008, 79, 284. [Google Scholar]
- Mosquera, M.; Llorente, M.; Riba, D.; Estebaranz, F.; González, M.; Lorenzo, C.; Sanmartí, N.; Toll, M.; Carbonell, E.; Feliu, O. Ethological study of manual laterality in naturalistic housed chimpanzees (Pan troglodytes) from the Mona Foundation Sanctuary (Girona, Spain). Laterality 2007, 12, 19–30. [Google Scholar]
- Armelles, A.; Llorente, M.; Feliu, O. Social interaction between three chimpanzee groups (Pan troglodytes) in Mona Foundation: Assessment of the unification probabilities. Folia Primatol. 2008, 79, 296. [Google Scholar]
- Rocha, S.; Gaspar, A.; Llorente, M. Asessment of two enrichment strategies in the rehabilitation of three rescued chimpanzees at the Mona Foundation Sanctuary. Folia Primatol. 2008, 79, 304. [Google Scholar]
- Carvalho, S.; Yamanashi, Y.; Yamakoshi, G.; Matsuzawa, T. Bird in the hand: Bossou chimpanzees (Pan troglodytes) capture West African wood-owls (Ciccaba woodfordi) but not to eat. Pan-Africa News 2010, 17, 6–9. [Google Scholar]
- Hirata, S.; Yamakoshi, G.; Fujta, S.; Ohasi, G.; Matsuzawa, T. Capturing and toying with hyraxes (Dendrohyrax dorsalis) by wild chimpanzees (Pan troglodytes) at Bossou, Guinea. Am. J. Primatol. 2001, 53, 93–97. [Google Scholar] [CrossRef]
- Sabater Pi, J.; Bermejo, M.; Illera, G.; Veà, J.J. Behavior of bonobos (Pan paniscus) following their capture of monkeys in Zaire. Int. J. Primatol. 1993, 14, 797–804. [Google Scholar] [CrossRef]
- Fujimoto, M.; Shimada, M. Newly observed predation of wild birds by M-group chimpanzees (Pan troglodytes schweinfurthii) at Mahale, Tanzania. Pan-Africa News 2008, 15, 23–26. [Google Scholar]
- Campi, A.; Llorente, M.; Riba, D.; Ferretti, J.; Yajeya, E. Enrichment, tool use and influences on behavioural pattern: chimpanzees (Pan troglodytes) vs. Macaques (Macaca sylvanus). Folia Primatol. 2011, 82, 321–402. [Google Scholar] [CrossRef]
- Duke, C.A.; Llorente, M. Use of inanimate novel enrichment with familiar/non-familar & conspecifics/humans stimulation among a group of captive chimpanzees. Folia Primatol. 2011, 82, 321–402. [Google Scholar] [CrossRef]
- Sepúlveda, Z.; Llorente, M.; Riba, D.; Feliu, O.; Gomara, A.; Sandoval, A.; Valsera, C. Tool use and tool making in rehabilitated and resocialized chimpanzees: Patterns of standardization in two artificial termite fishing activities. Folia Primatol. 2009, 80, 423. [Google Scholar]
- Sepúlveda, Z.; Llorente, M.; Riba, D.; Feliu, O.; Gomara, A.; Sandoval, A.; Valsera, C. Behavioural effects of two enrichment activities: The influence of tool use and making. Folia Primatol. 2009, 80, 424. [Google Scholar]
- Llorente, M.; Riba, D.; Campi, A.; Ferretti, J.; Armelles, A.; Ribas, M.; Rostán, C.; Feliu, O. Is it possible to re-socialize abused primates? A 5-year longitudinal study on the social behaviour of chimpanzees from the Mona Foundation. Folia Primatol. 2011, 82, 321–402. [Google Scholar] [CrossRef]
- Hickey, T.E.; Kelly, W.A.; Sitzman, J.E. Demodectic mange in a tamarin (Saguinus geoffroyi). Lab. Anim. Sci. 1983, 33, 192–193. [Google Scholar]
- Calle, P.P.; Bowerman, D.L.; Pape, W.J. Nonhuman primate tularemia (Francisella tularensis) epizootic in a zoological park. J. Zoo Wildlife Med. 1993, 24, 459–468. [Google Scholar]
- Maetz-Rensing, K.; Floto, A.; Schrod, A.; Becker, T.; Finke, E.J.; Seibold, E.; Splettstoesser, W.D.; Kaup, F.J. Epizootic of tularemia in an outdoor housed group of cynomolgus monkeys (Macaca fascicularis). Vet. Pathol. 2007, 44, 327–334. [Google Scholar] [CrossRef]
- Speck, S.; Pauly, A.; Stark, R. Fatal case of Salmonella typhimurium infection in a cherry crowned mangabey (Cercocebus torquatus). Verh. ber. Erkrg. Zootiere 2007, 43, 321–324. [Google Scholar]
- Knöbl, T.; Rocha, L.T.; Menão, M.C.; Igayara, C.A.S.; Paixão, R.; Moreno, A.M. Salmonella Yoruba infection in white-tufted-ear marmoset (Callithrix jacchus). Pesquisa Vet. Brasil. 2011, 31, 707–710. [Google Scholar] [CrossRef]
- Ulrich, R.; Boer, M.; Herder, V.; Spitzbarth, I.; Hewicker-Trautwein, M.; Baumgartner, W.; Wohlsein, P. Epizootic fatal amebiasis in an outdoor group of Old World monkeys. J. Med. Primatol. 2010, 39, 160–165. [Google Scholar] [CrossRef]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Swaisgood, R.R. Current status and future directions of applied behavioral research for animal welfare and conservation. Appl. Anim. Behav. Sci. 2007, 102, 139–162. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Llorente, M.; Riba, D.; Mosquera, M.; Ventura, M.; Feliu, O. Hunting Activity Among Naturalistically Housed Chimpanzees (Pan troglodytes) at the Fundació Mona (Girona, Spain). Predation, Occasional Consumption and Strategies in Rehabilitated Animals. Animals 2012, 2, 363-376. https://doi.org/10.3390/ani2030363
Llorente M, Riba D, Mosquera M, Ventura M, Feliu O. Hunting Activity Among Naturalistically Housed Chimpanzees (Pan troglodytes) at the Fundació Mona (Girona, Spain). Predation, Occasional Consumption and Strategies in Rehabilitated Animals. Animals. 2012; 2(3):363-376. https://doi.org/10.3390/ani2030363
Chicago/Turabian StyleLlorente, Miquel, David Riba, Marina Mosquera, Mei Ventura, and Olga Feliu. 2012. "Hunting Activity Among Naturalistically Housed Chimpanzees (Pan troglodytes) at the Fundació Mona (Girona, Spain). Predation, Occasional Consumption and Strategies in Rehabilitated Animals" Animals 2, no. 3: 363-376. https://doi.org/10.3390/ani2030363