
A Mineralized Alga and Acritarch Dominated Microbiota from the Tully Formation (Givetian) of Pennsylvania, USA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
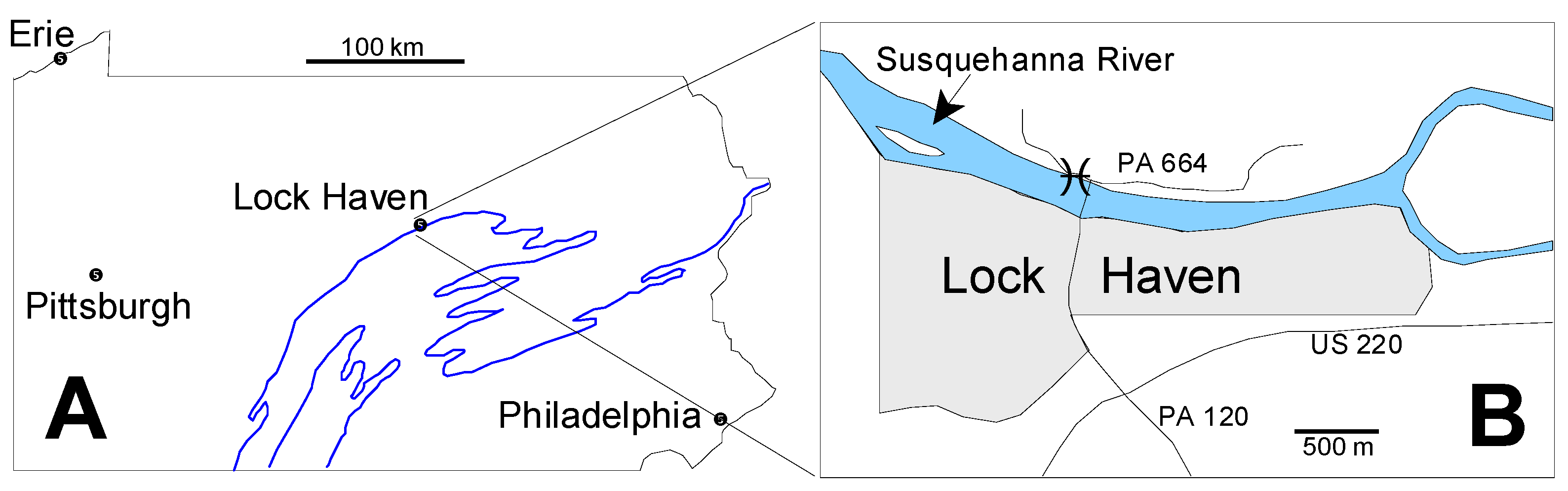
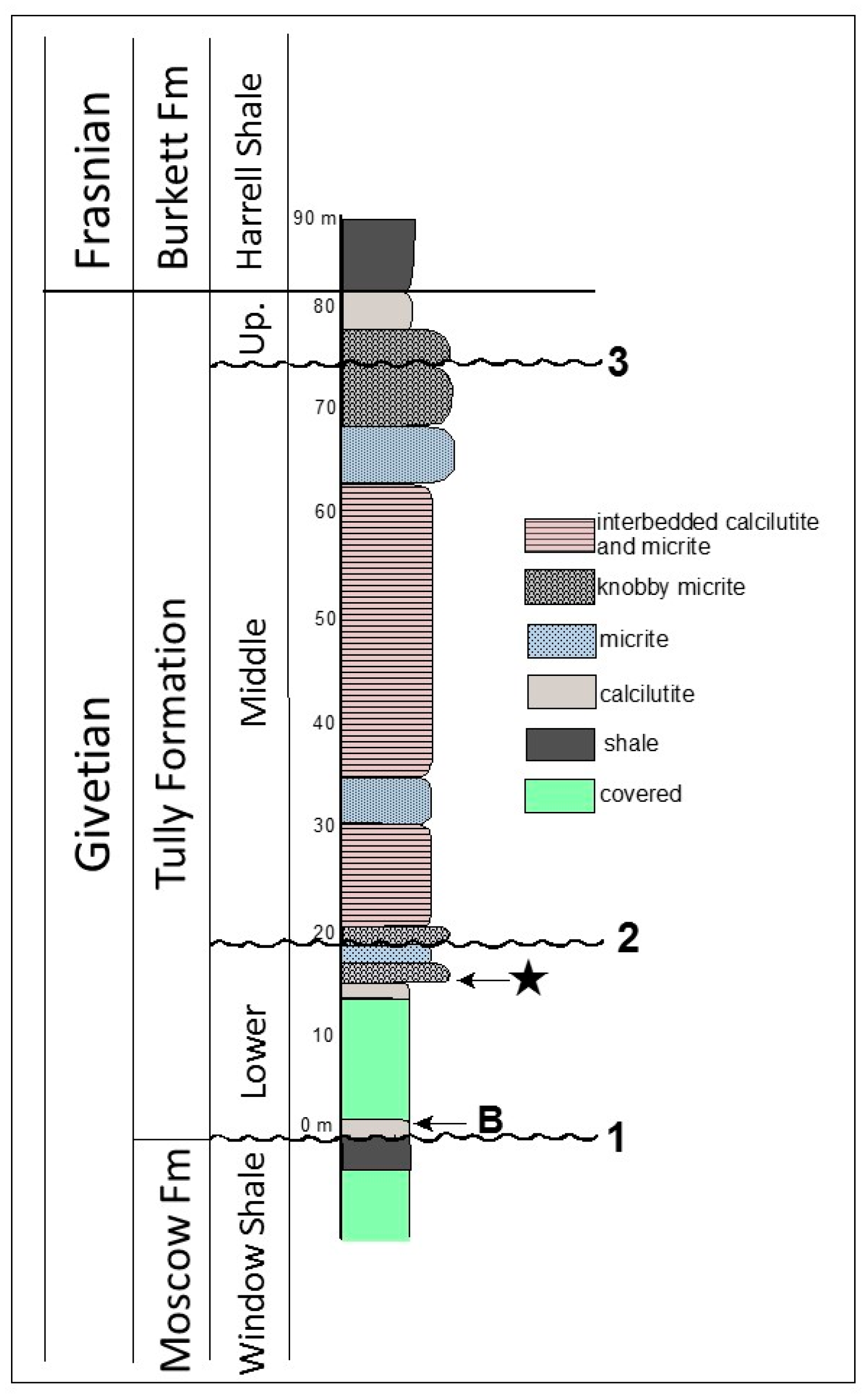
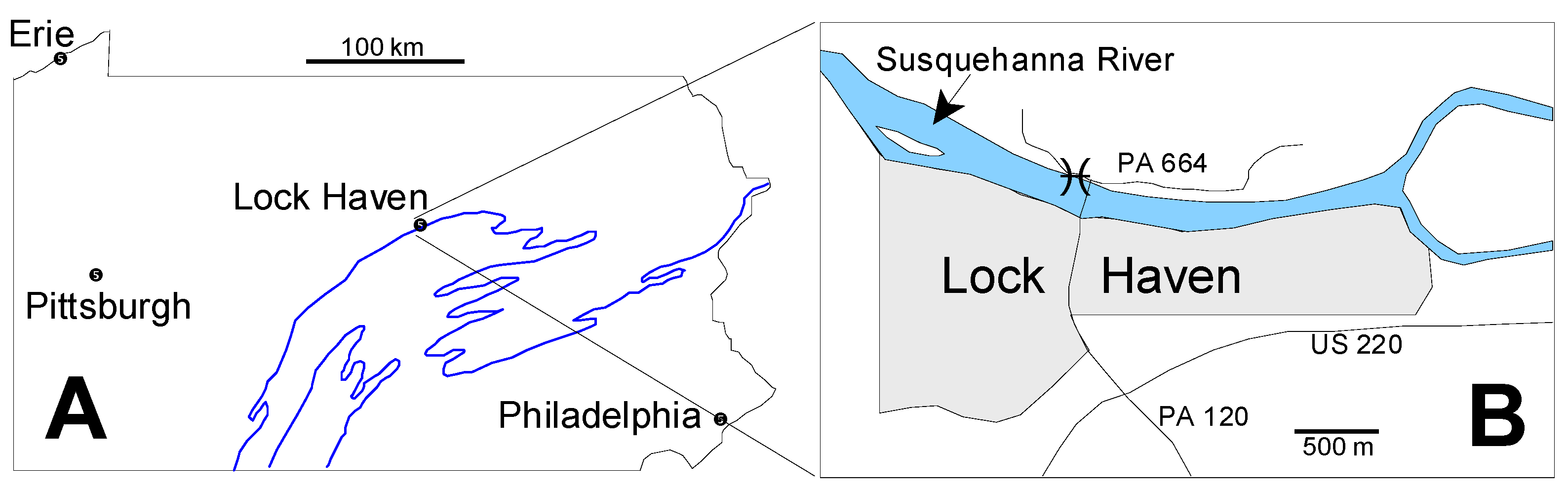
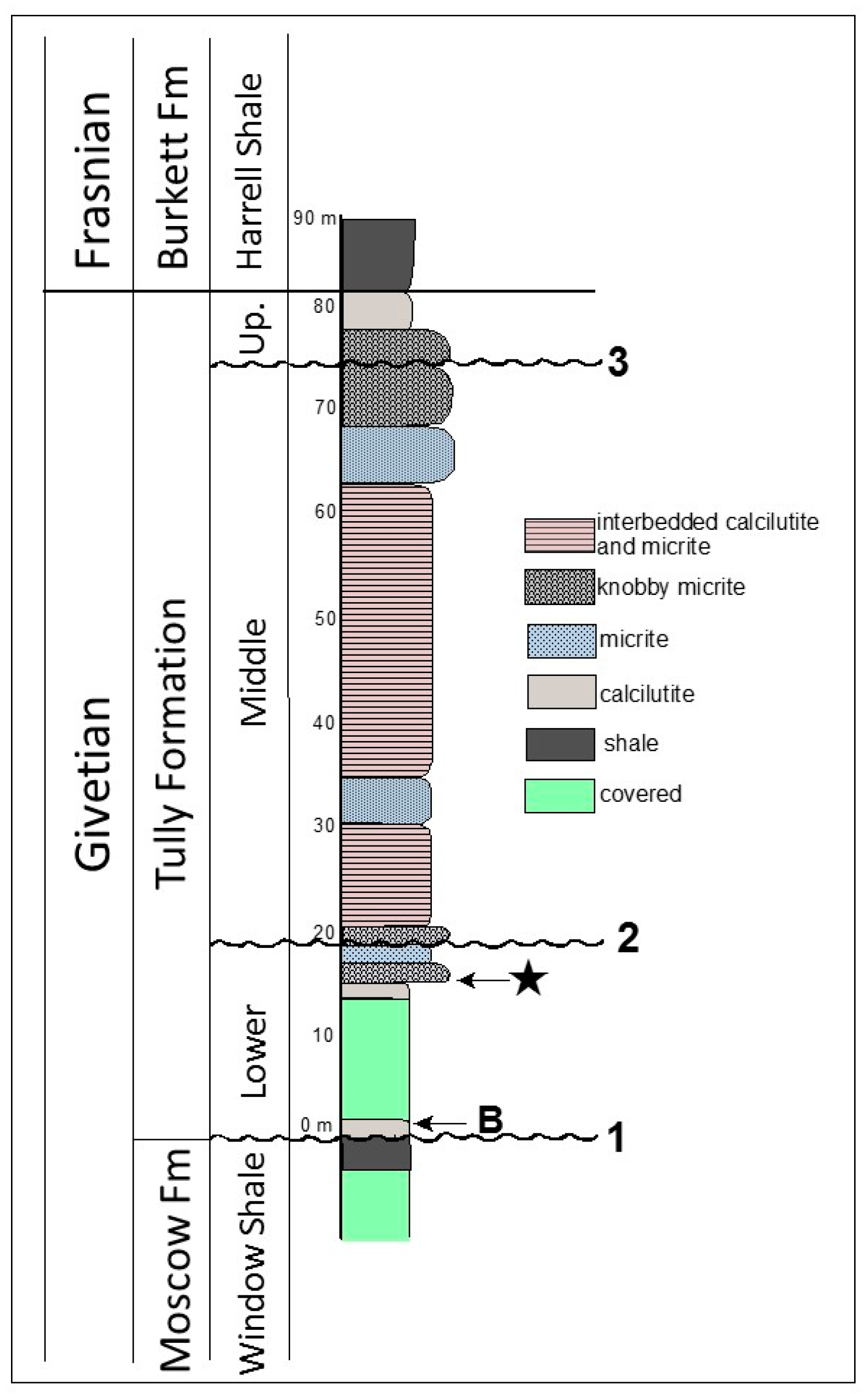
2.1. Geologic Setting
2.2. Microfossil Occurrence
2.3. Preparation Methods
2.4. SEM Methods
2.5. Thin Section Methods
2.6. Specimen Repository
3. Results
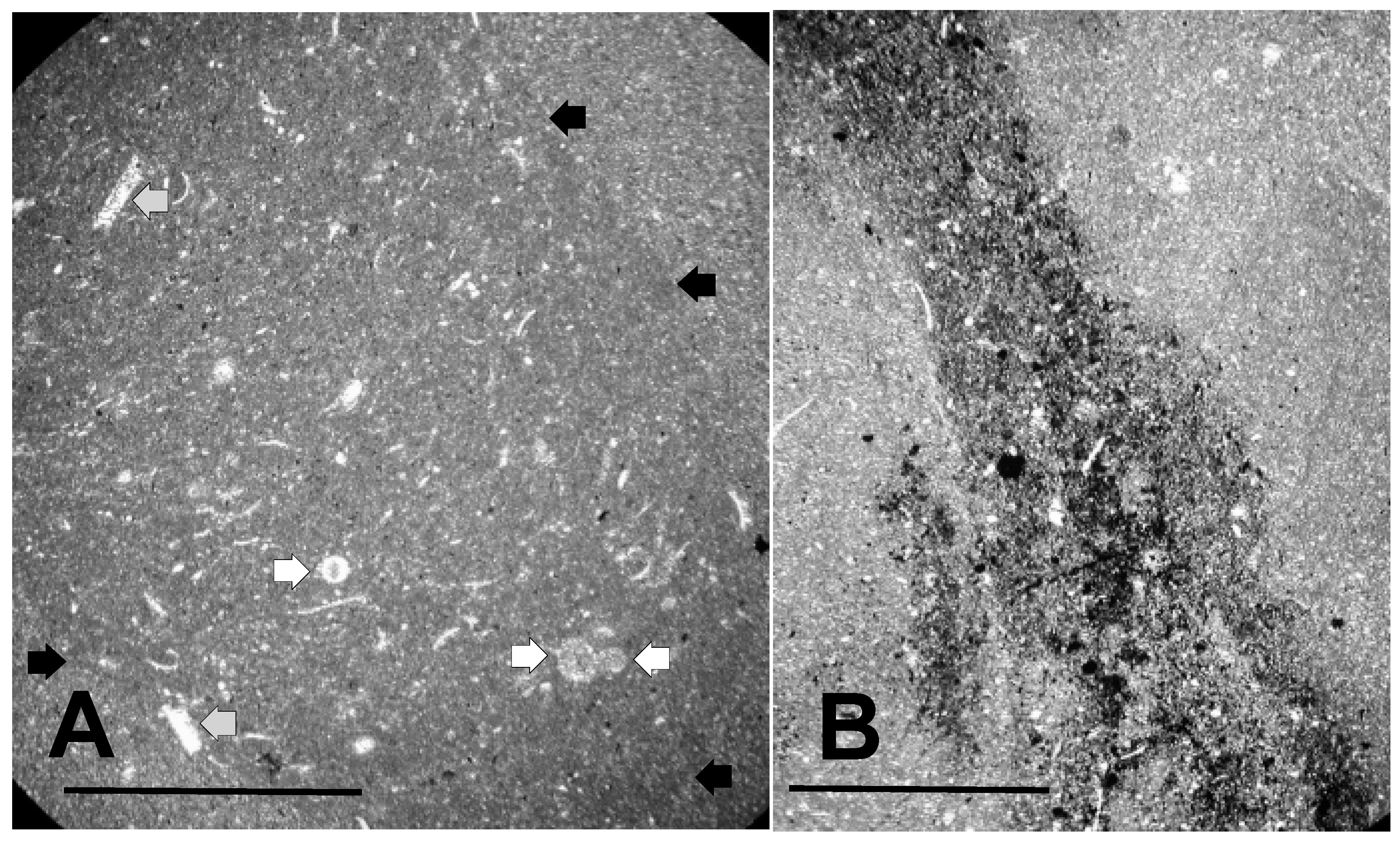
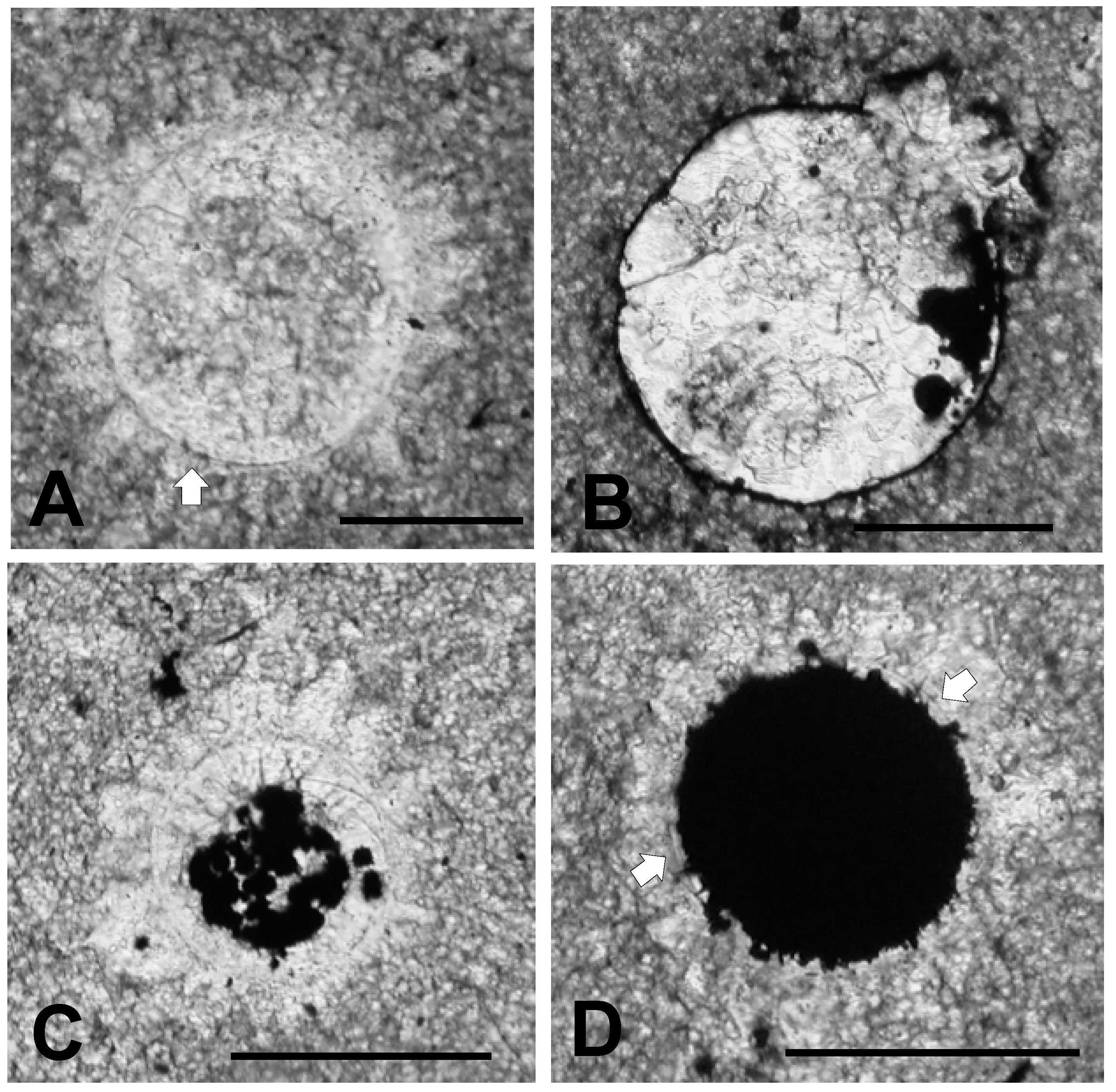
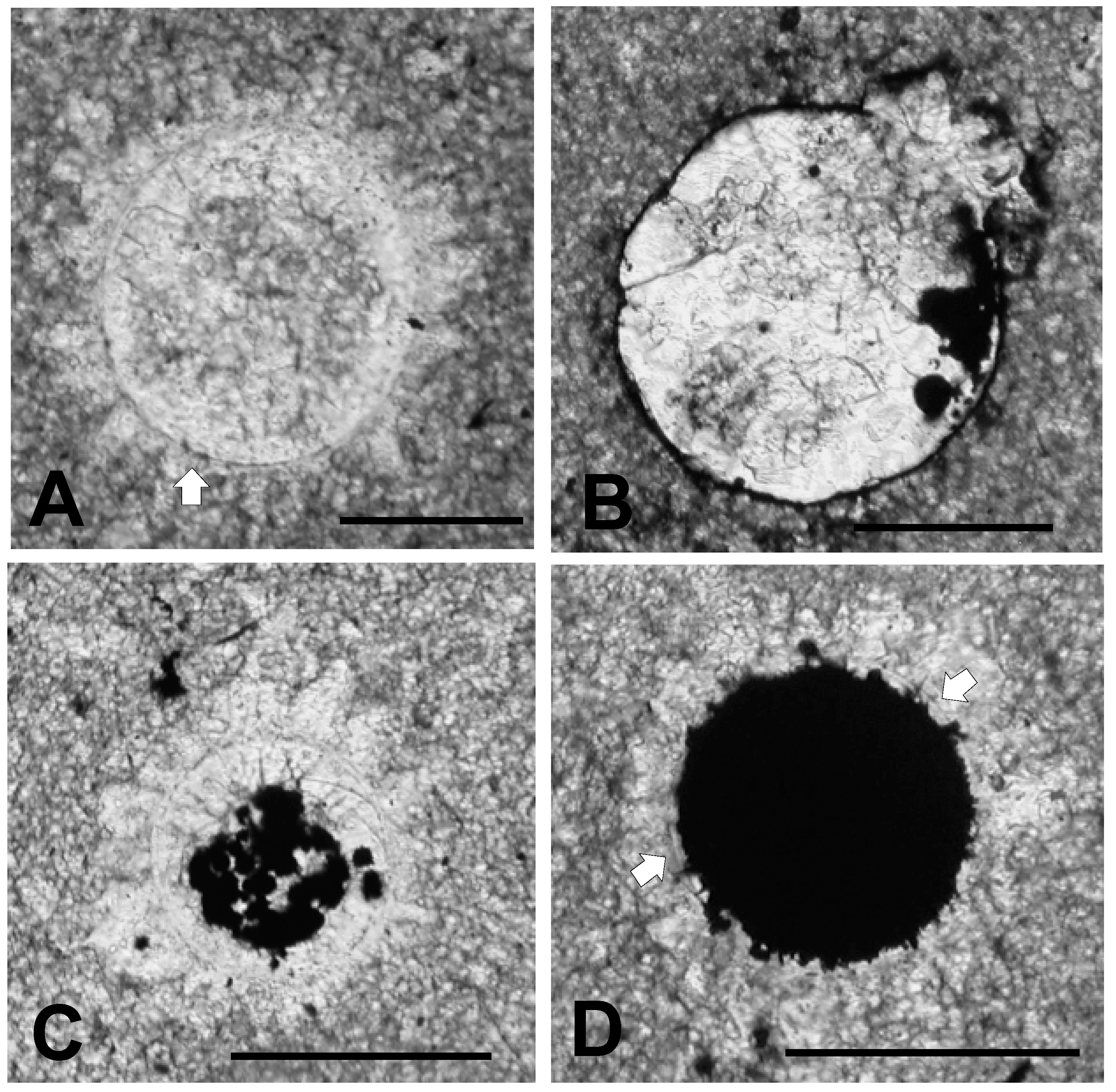
3.1. Thin Section Results
3.1.1. Rock Texture
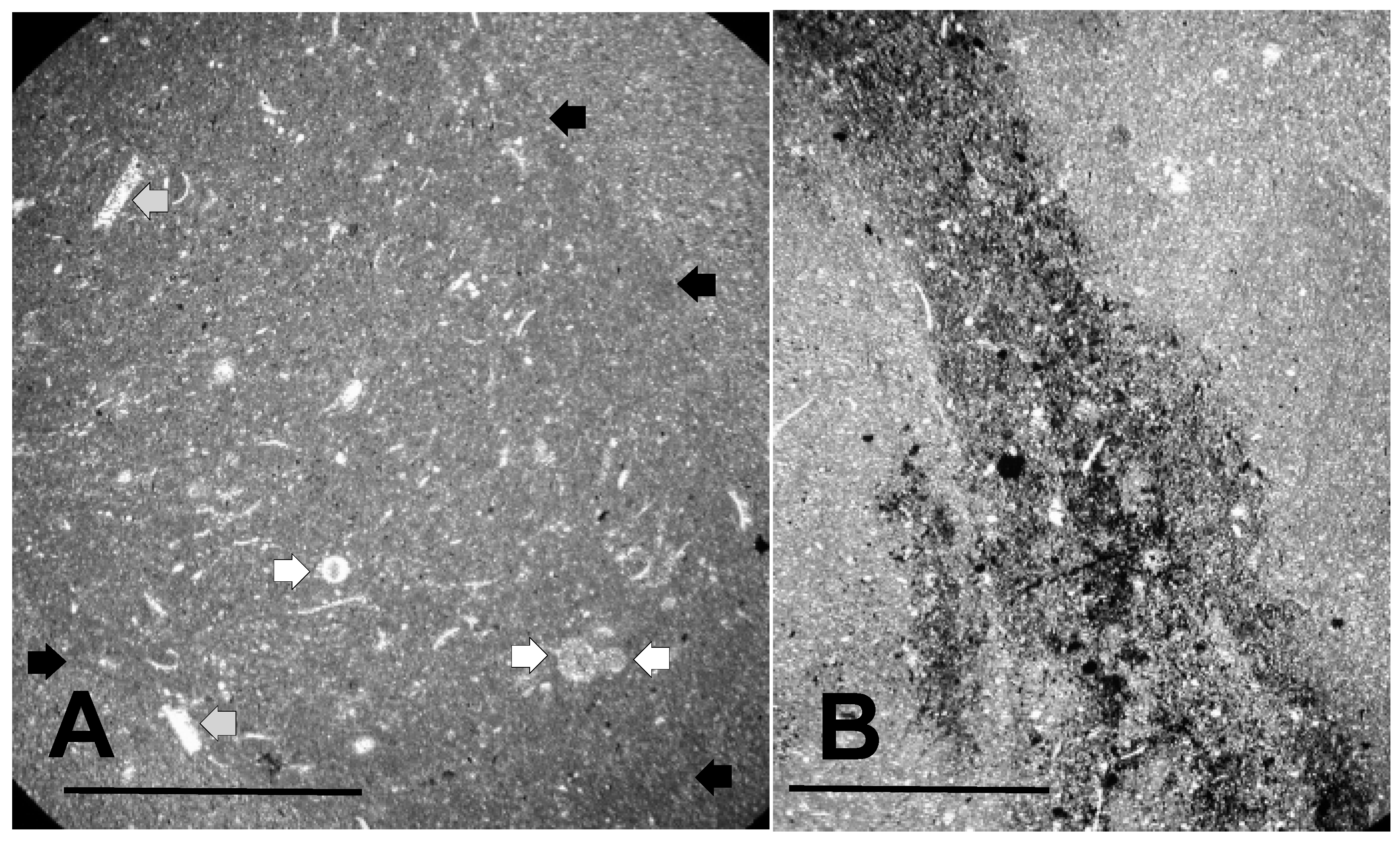
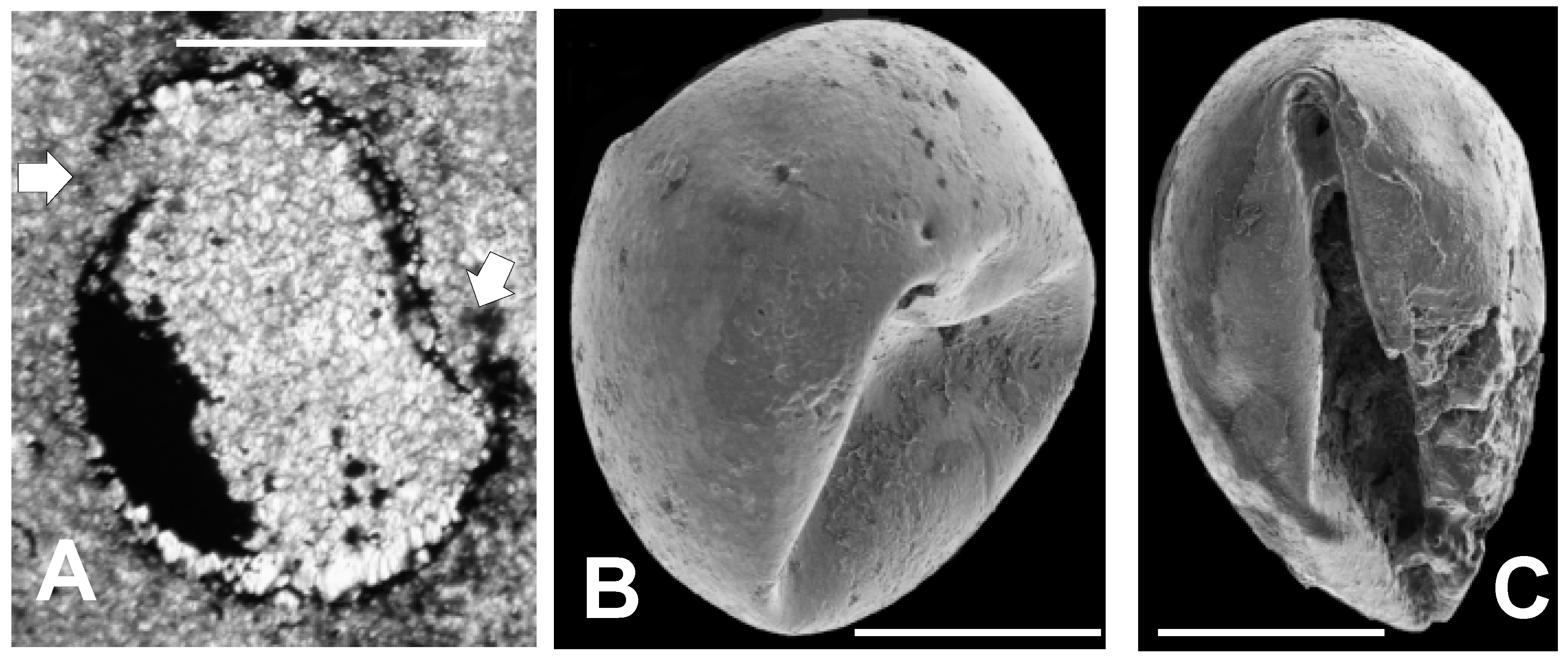
3.1.2. Shell Fragments
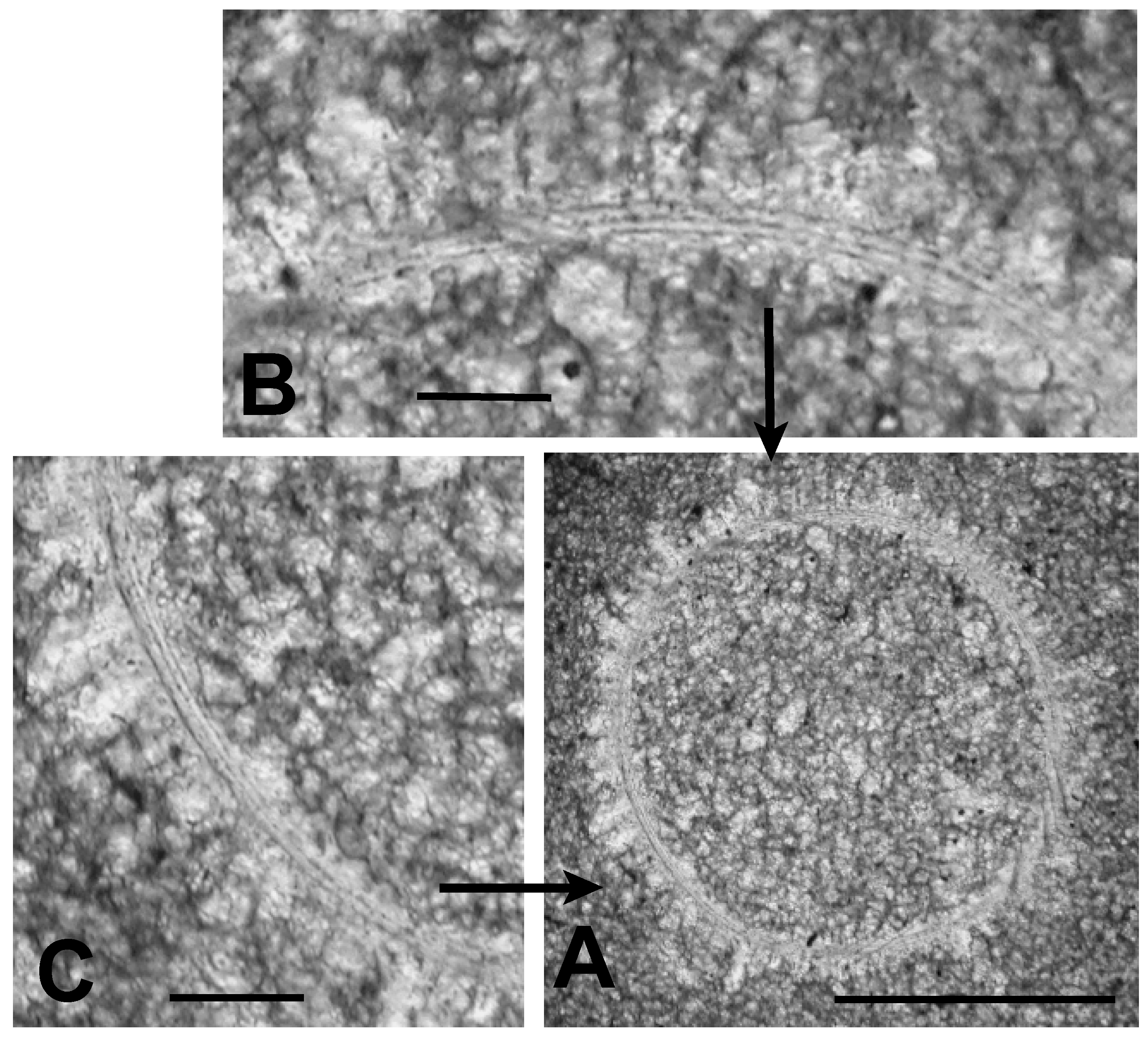
3.1.3. Microalga and Acritarch Remains
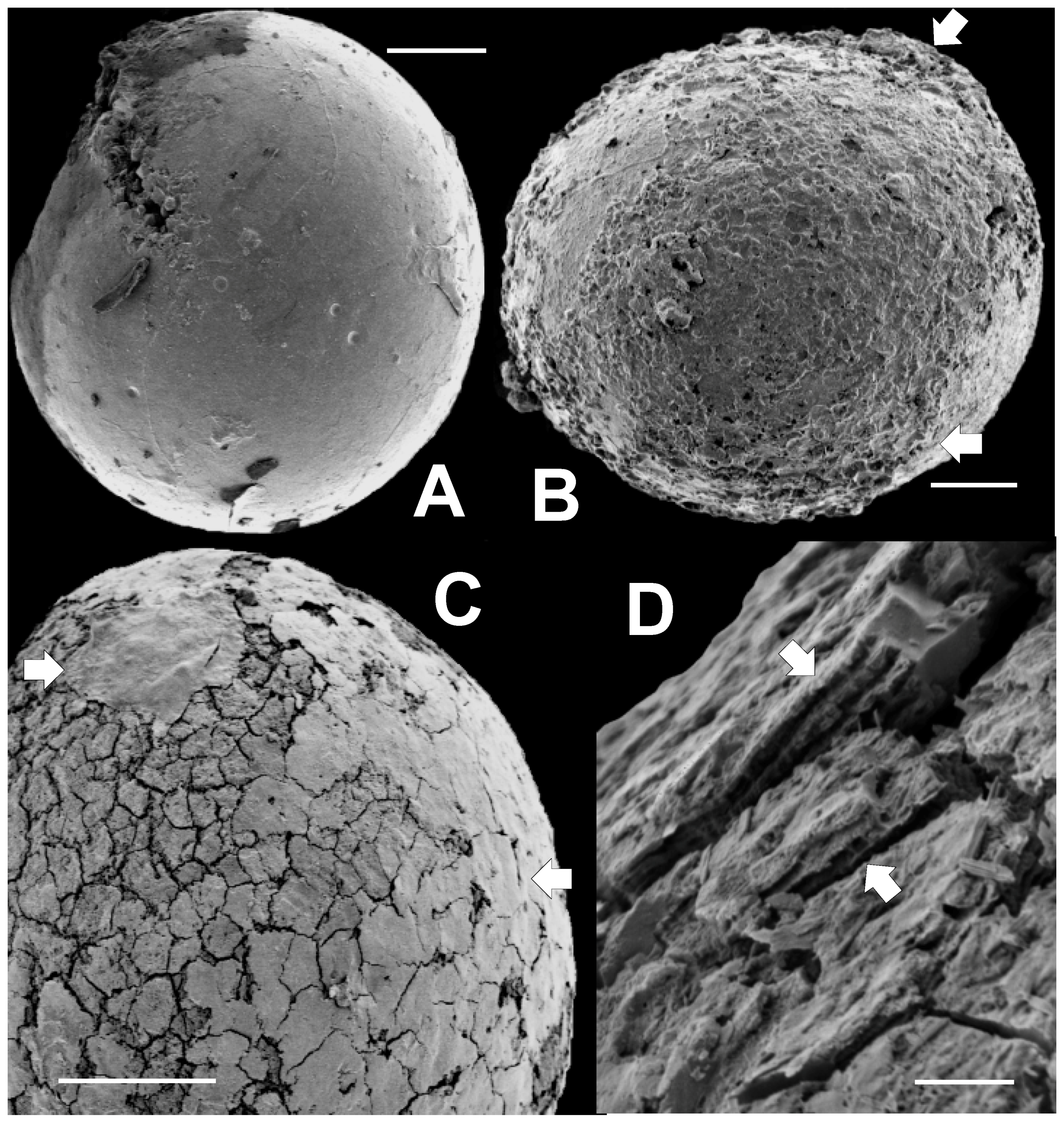
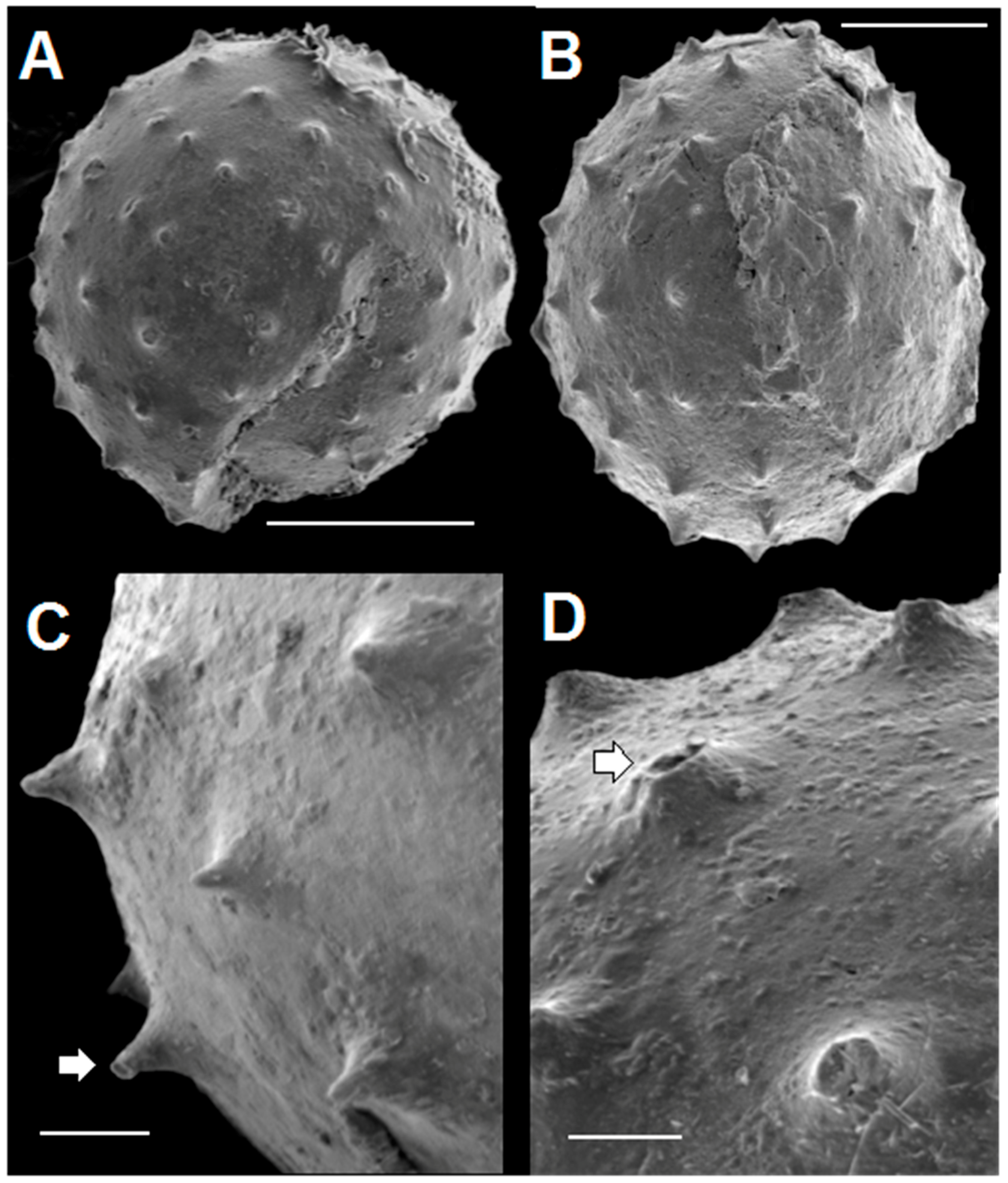
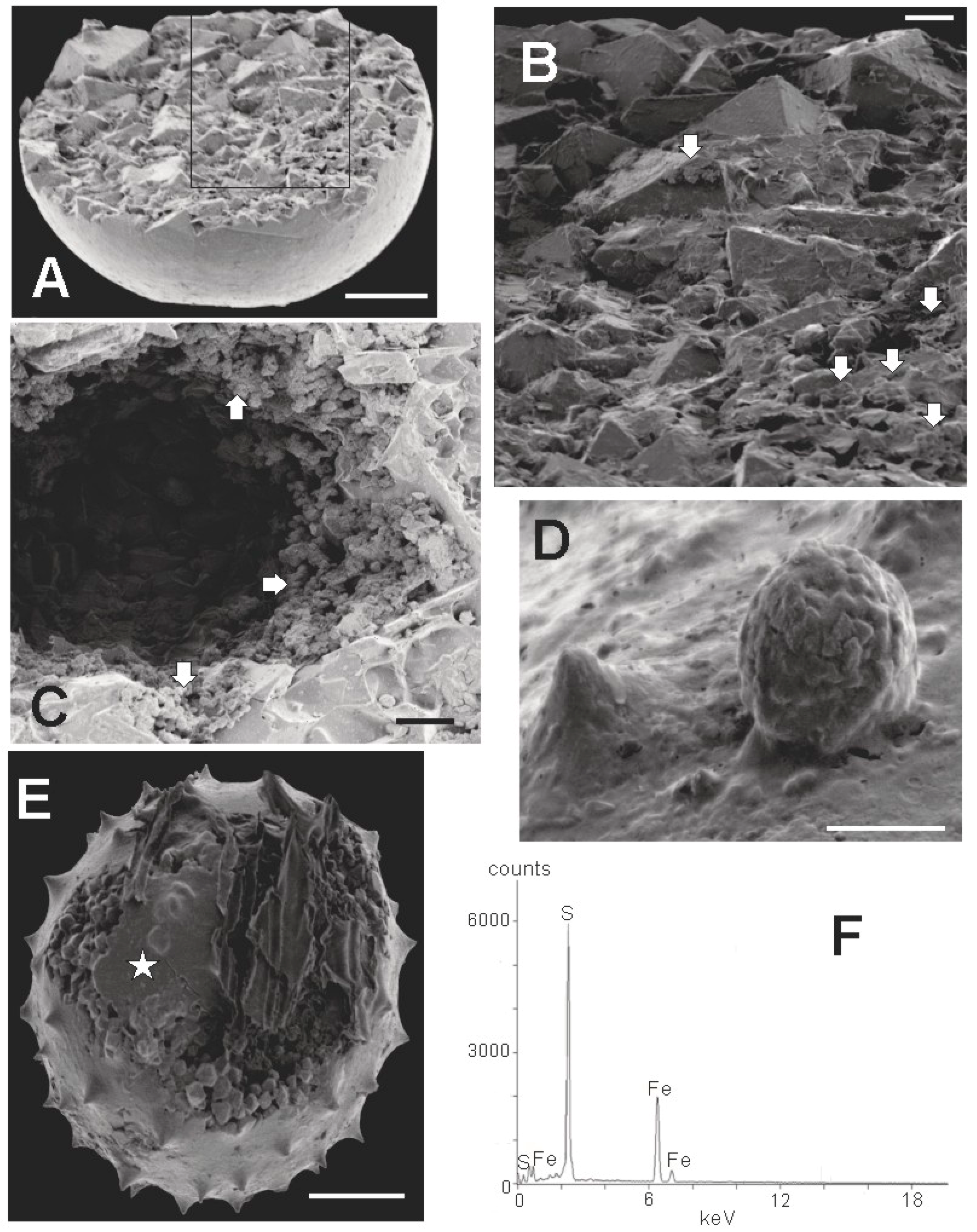
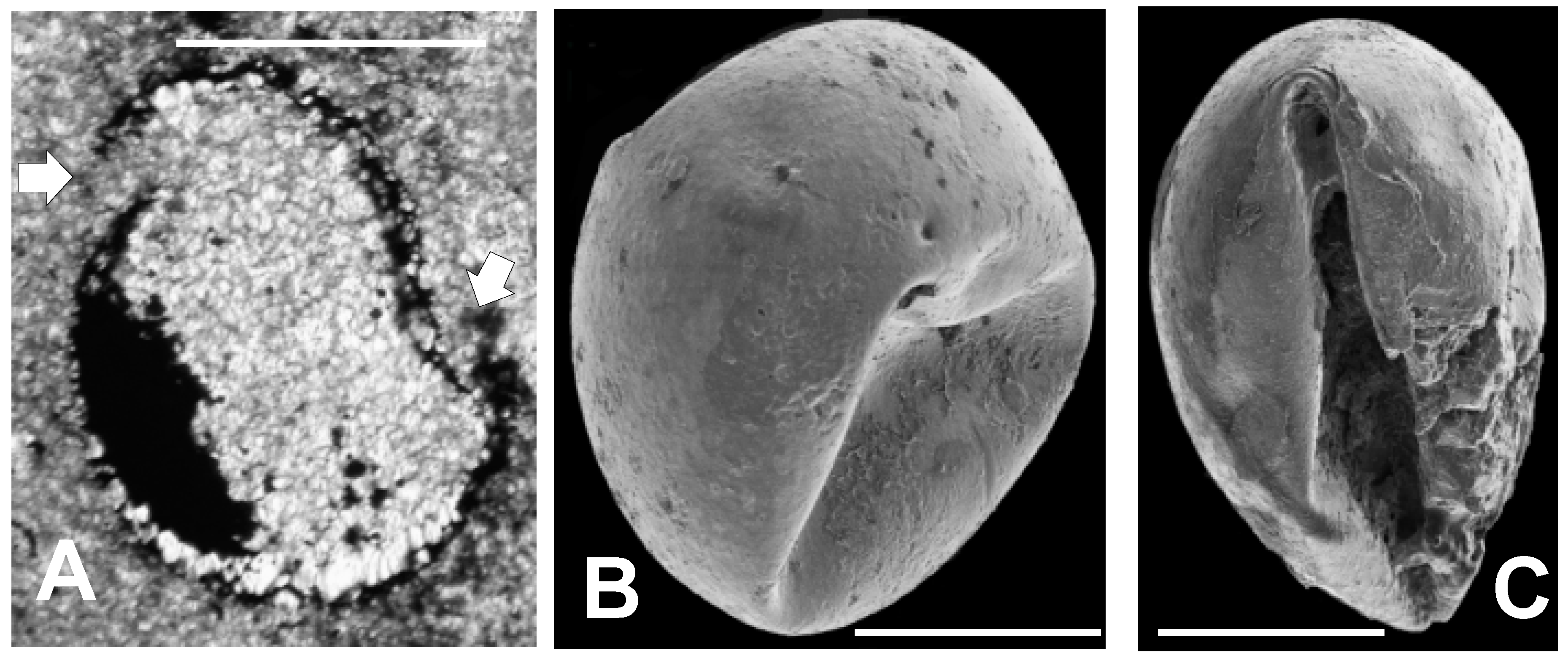
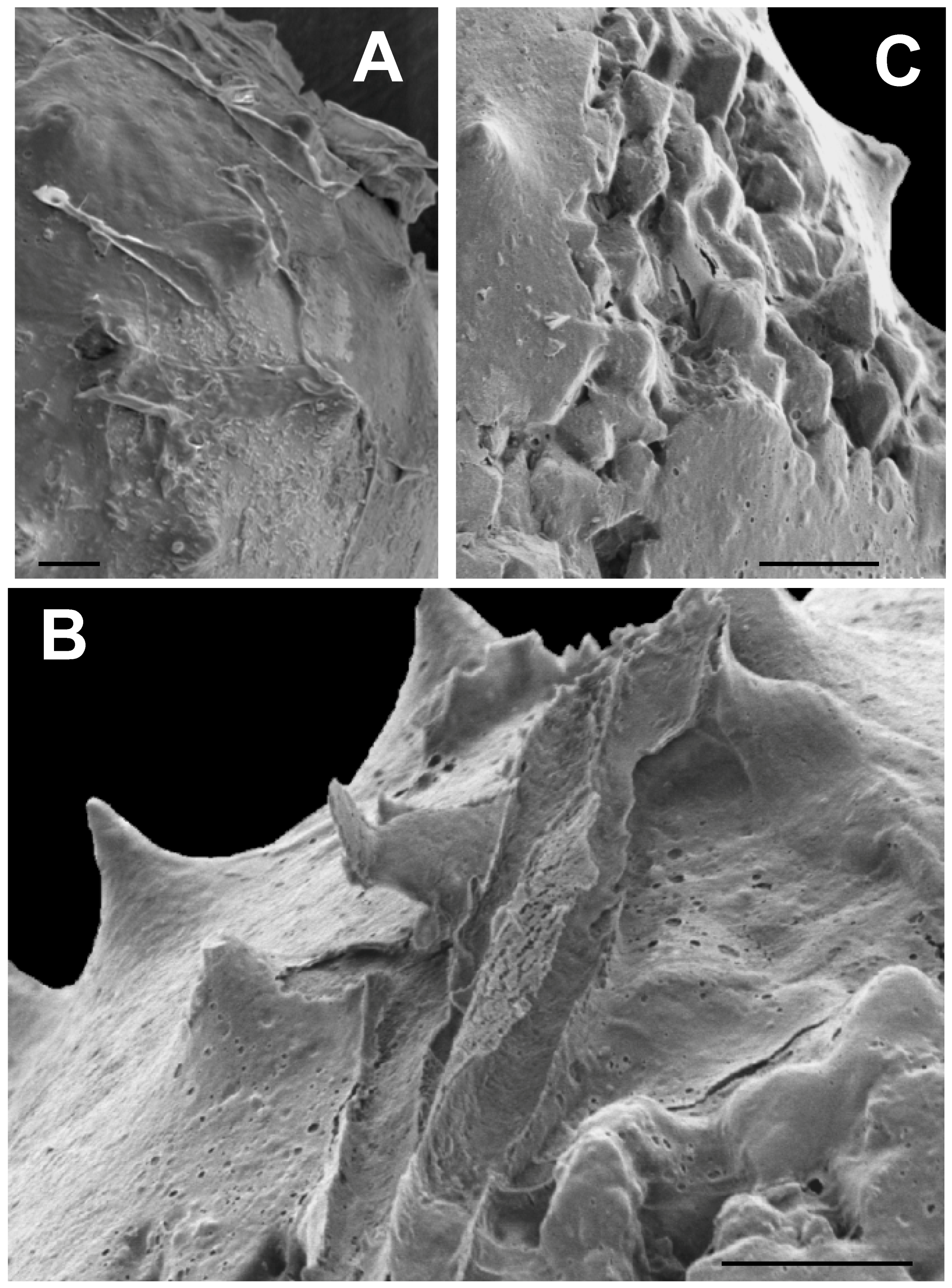
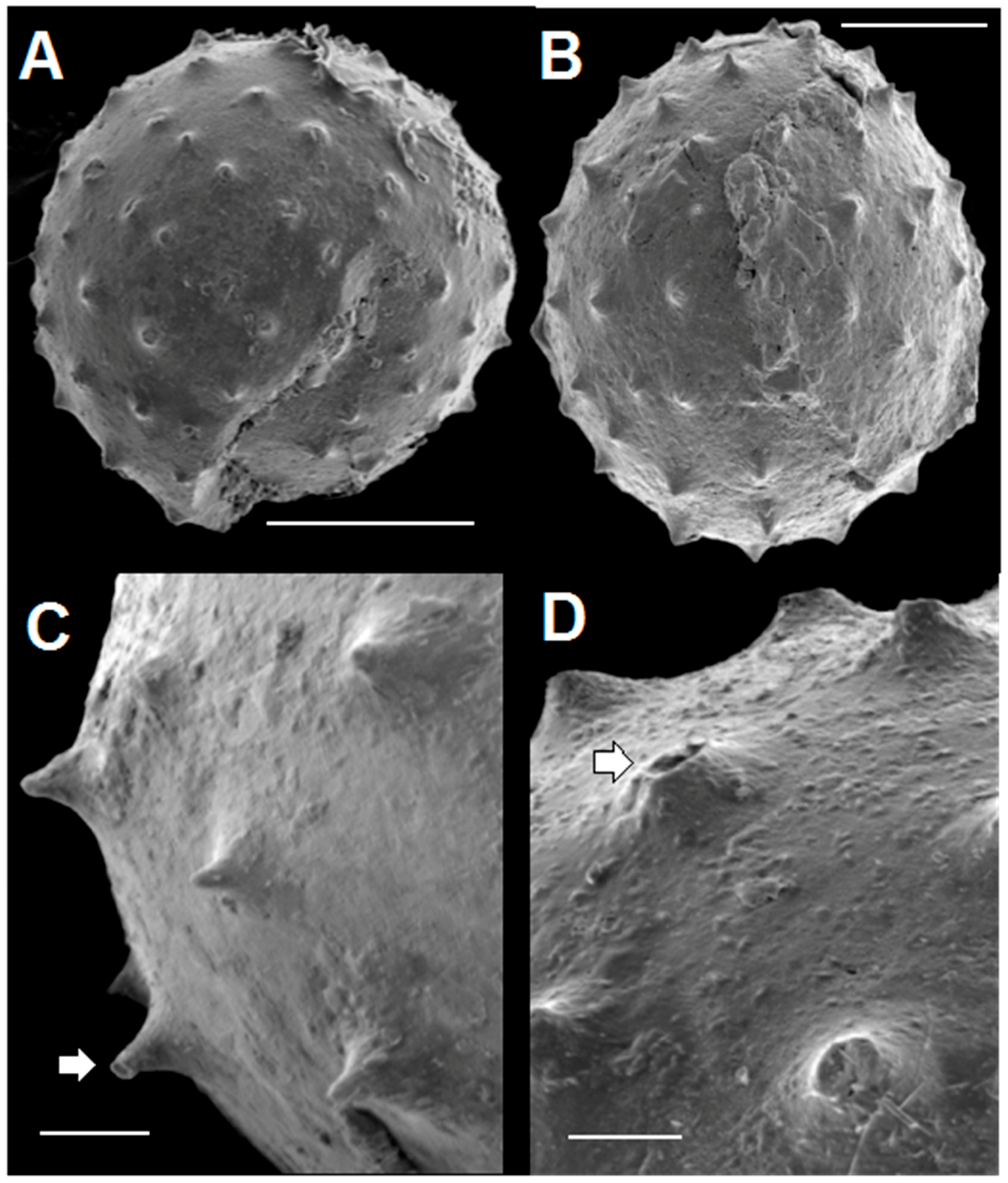
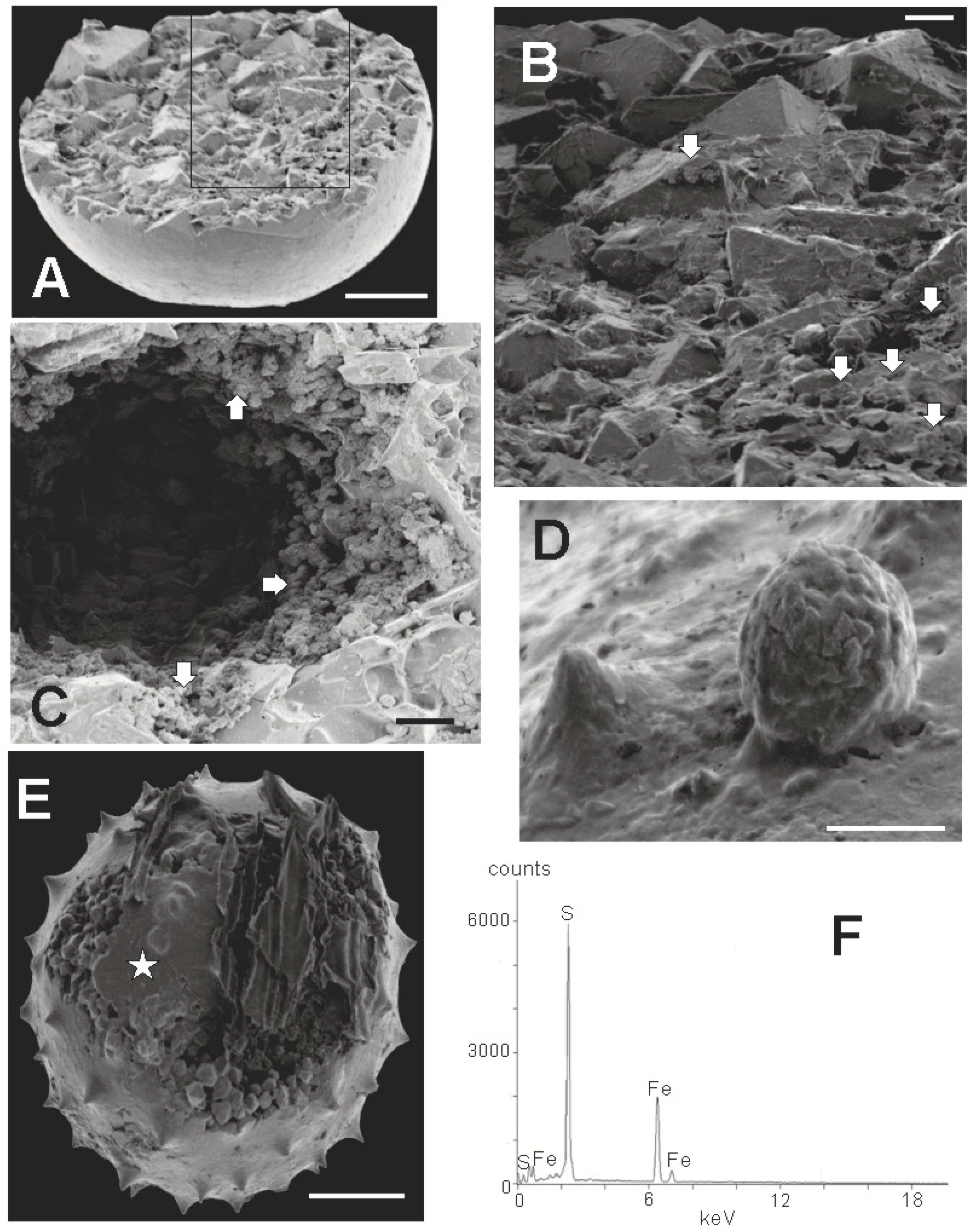
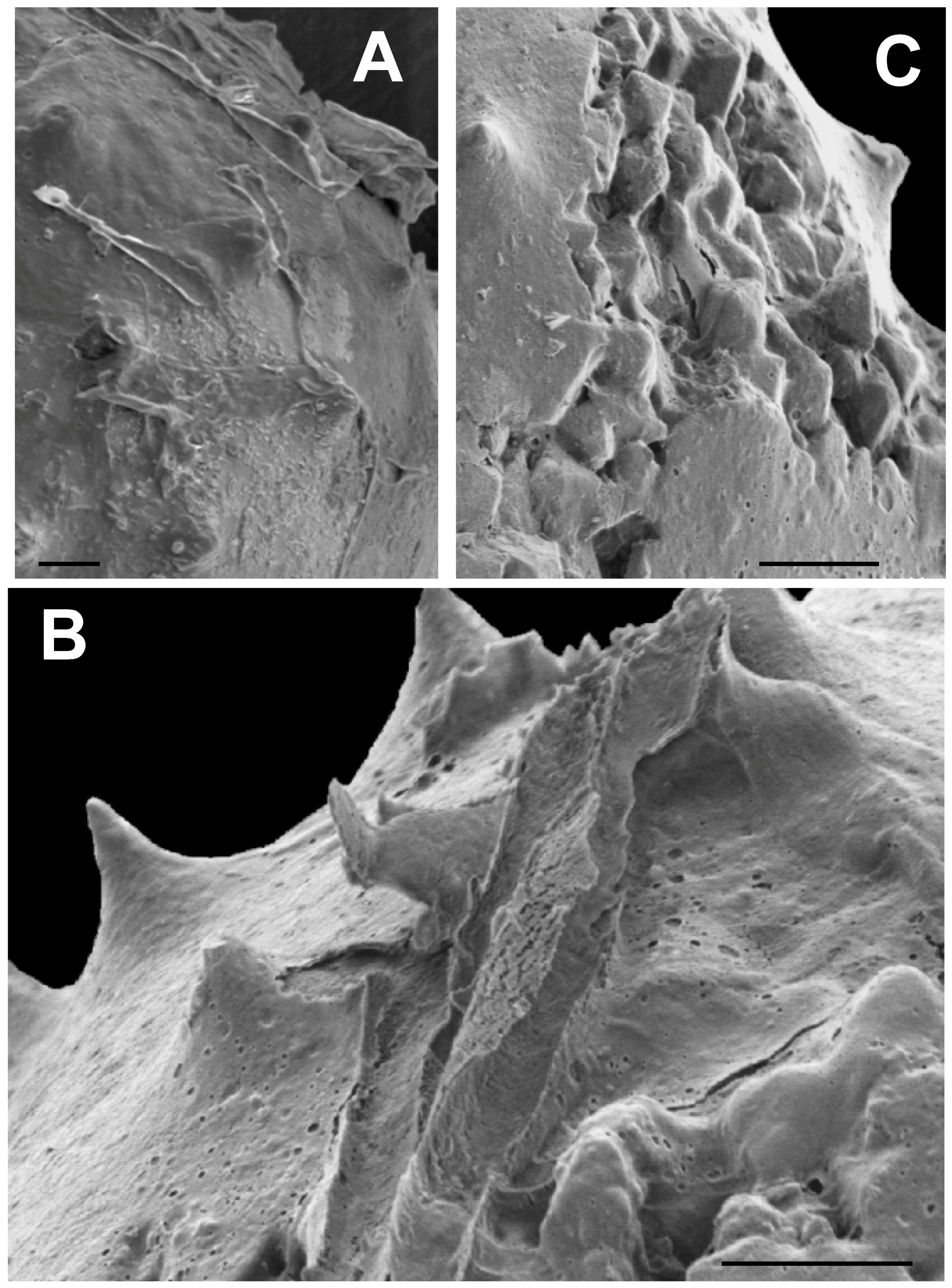
3.2. SEM Results
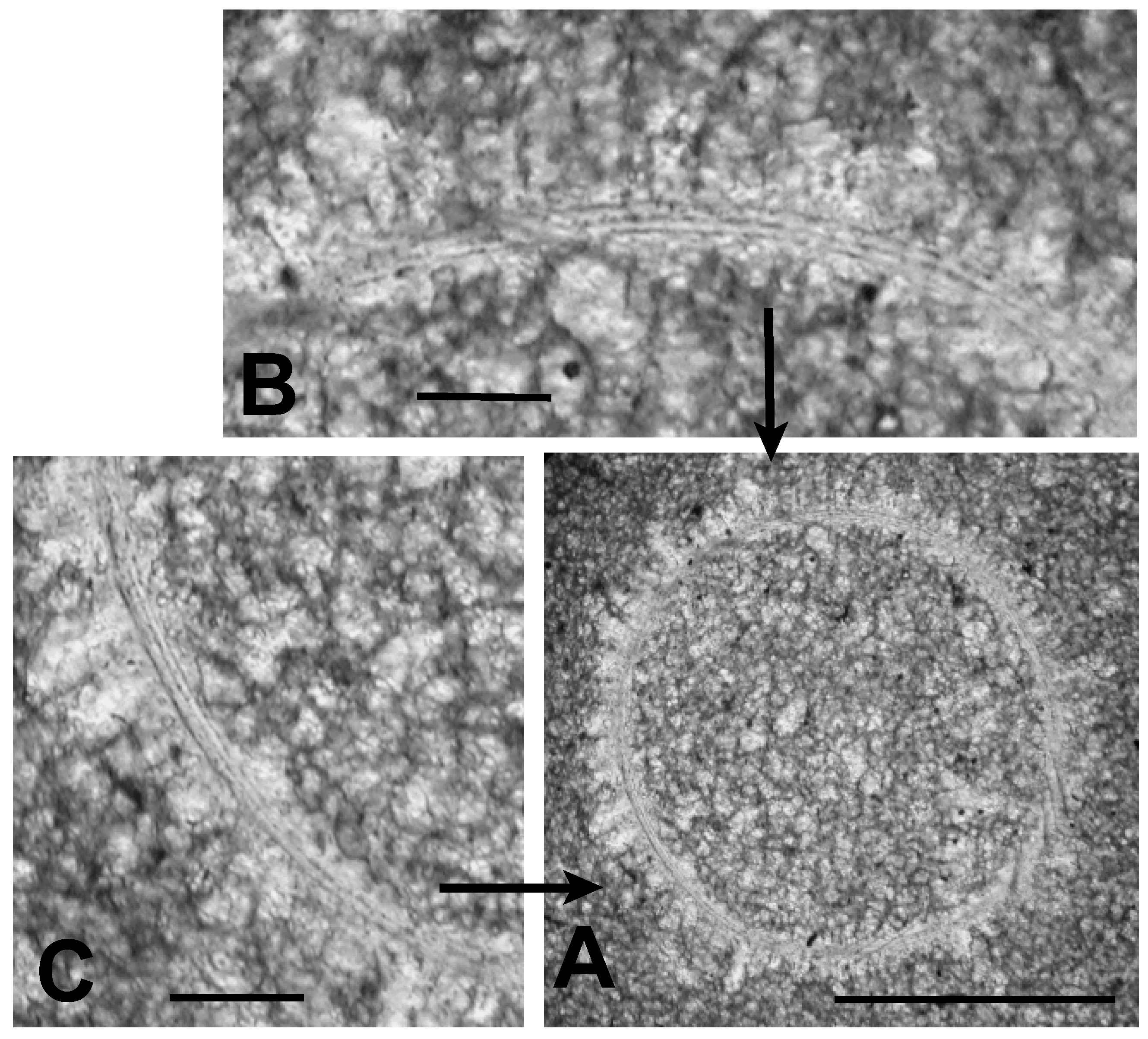
3.2.1. Sphaeromorphic Specimens
3.2.2. Acanthomorphic Specimens
3.2.3. Crystallinity of Extracted Specimens
3.2.4. Deformed Specimens
3.2.5. Surface Covering
4. Systematic Paleontology
4.1. Algae and Acritarchs
4.1.1. Descriptive Systematics: Algae
- Super-group: Archaeplastida Adl et al. [76]
- First Order Subdivision: Chloroplastida Adl et al. [76]
- Order: Pterospermatales: Tappan [84]
- Family: Tasmanitaceae: Tappan [84]
- Type Species: Tasmanites punctatus Newton [85]
- Tasmanites. cf. T. sinuosus Winslow [56]
- Tasmanites. cf. T. sommeri Winslow [56]
4.1.2. Descriptive Systematics: Acritarchs
- Group: Acritarcha Evitt [20]
- Subgroup: Acanthomorphitae Downie, Evitt and Sarjeant [93]
- emend. Moczydłowska [96]
- Type species: Solisphaeridium stimuliferum Deflandre [97]
- Solisphaeridium (?)
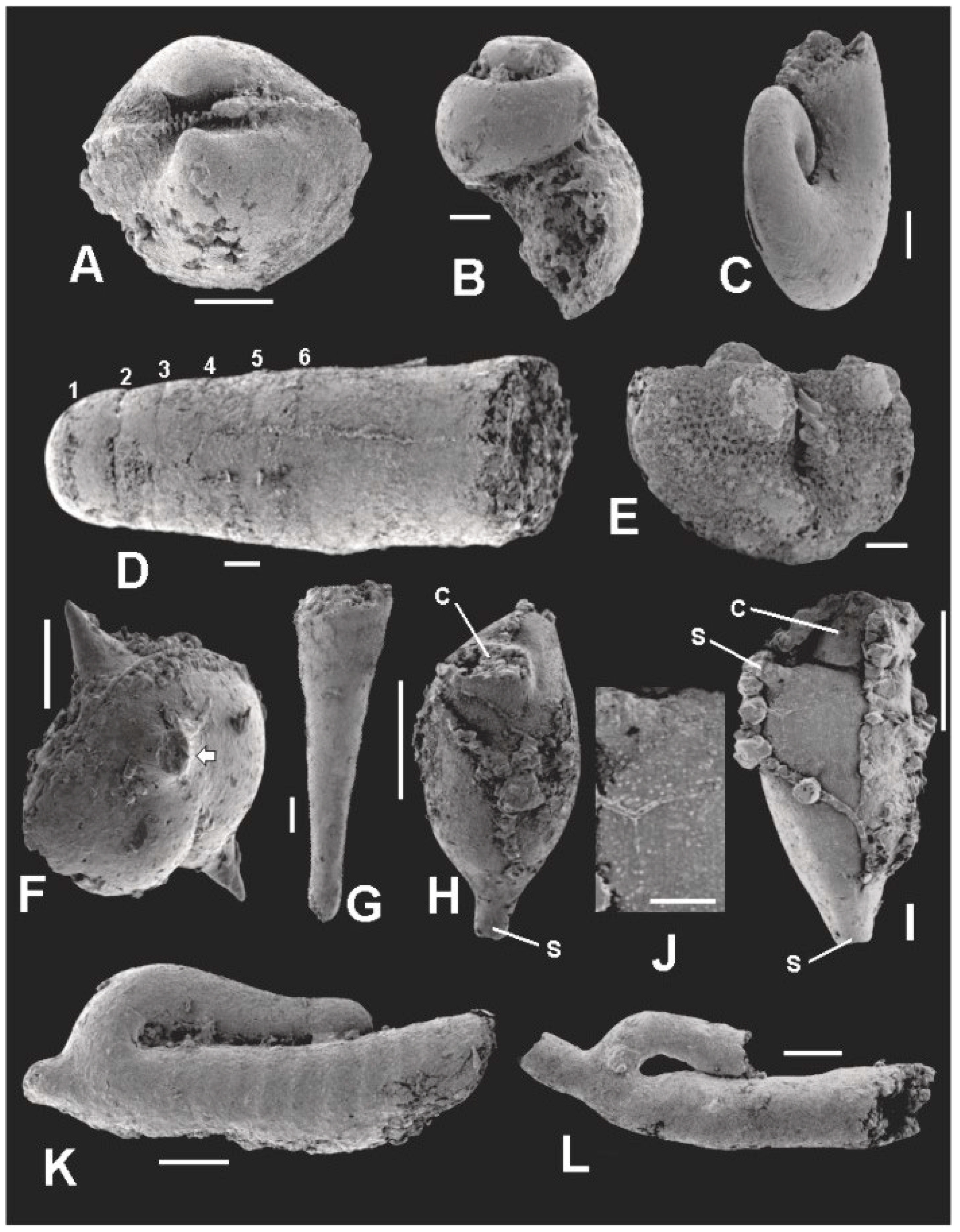
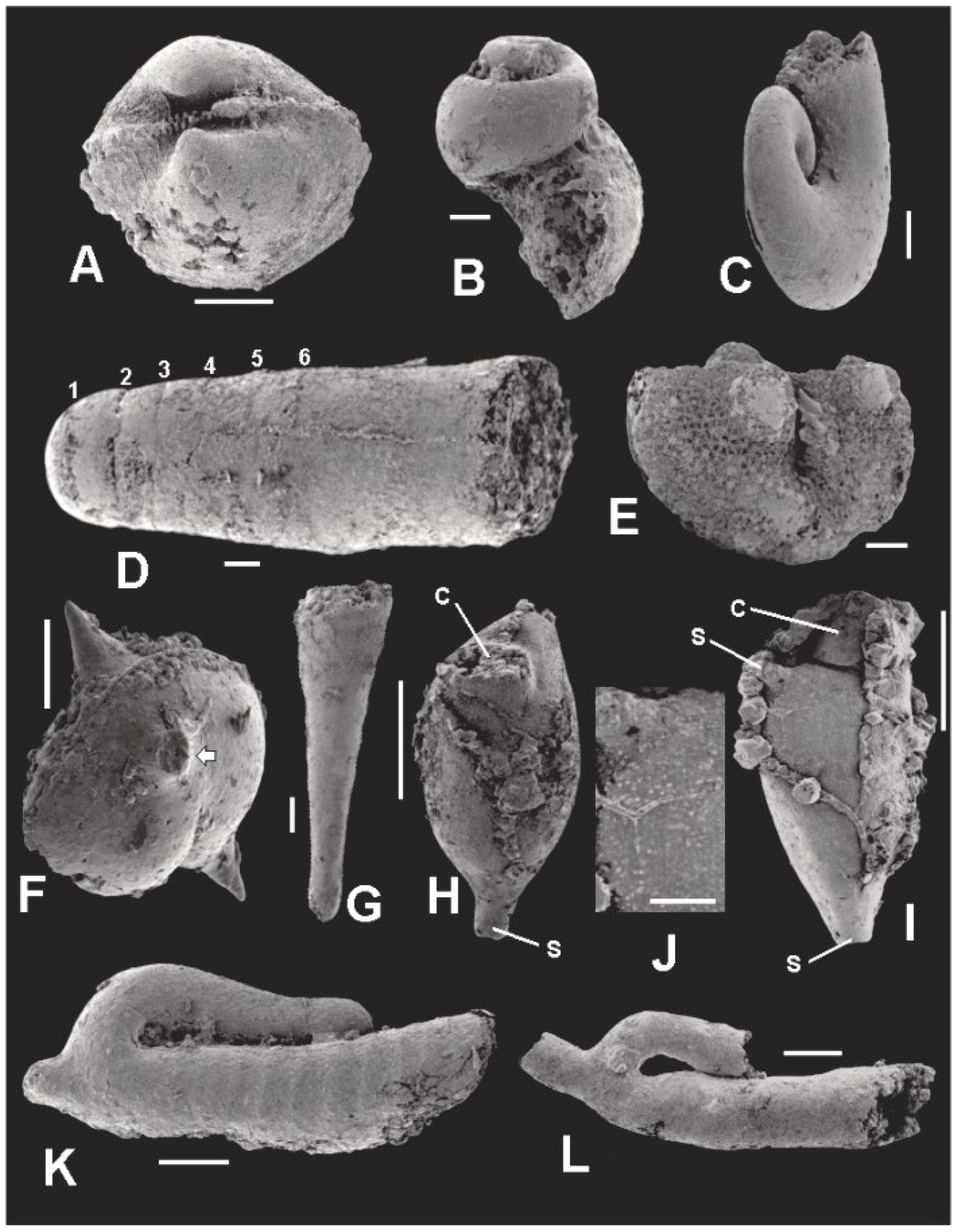
5. Co-Occurring Organisms
5.1. Molluscs
5.1.1. Bivalves
5.1.2. Gastropods
5.1.3. Cephalopods
5.2. Ostracodes
5.3. Dacryoconarids
5.4. Bryozoans
5.5. Jinonicellids
6. Discussion
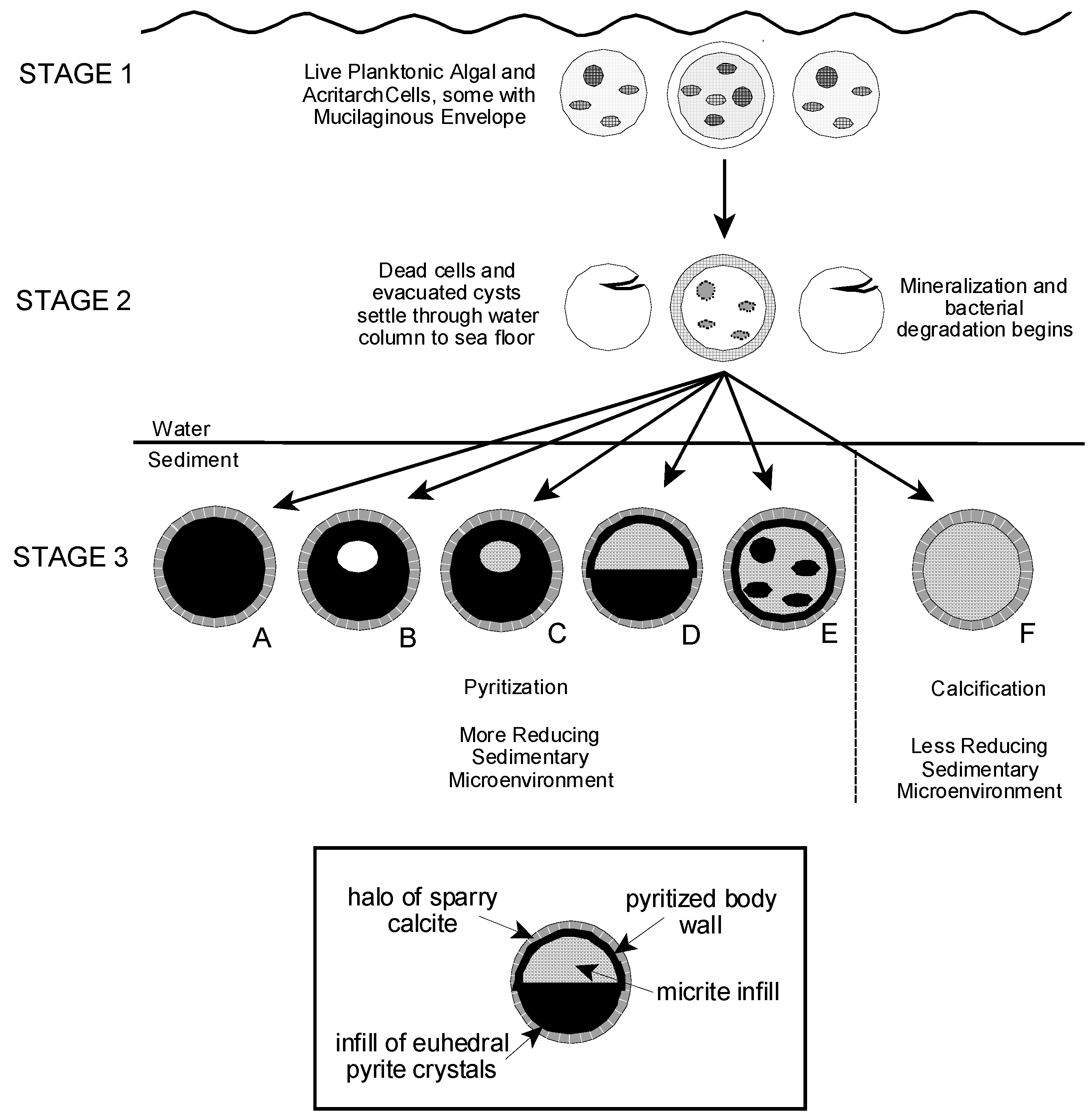
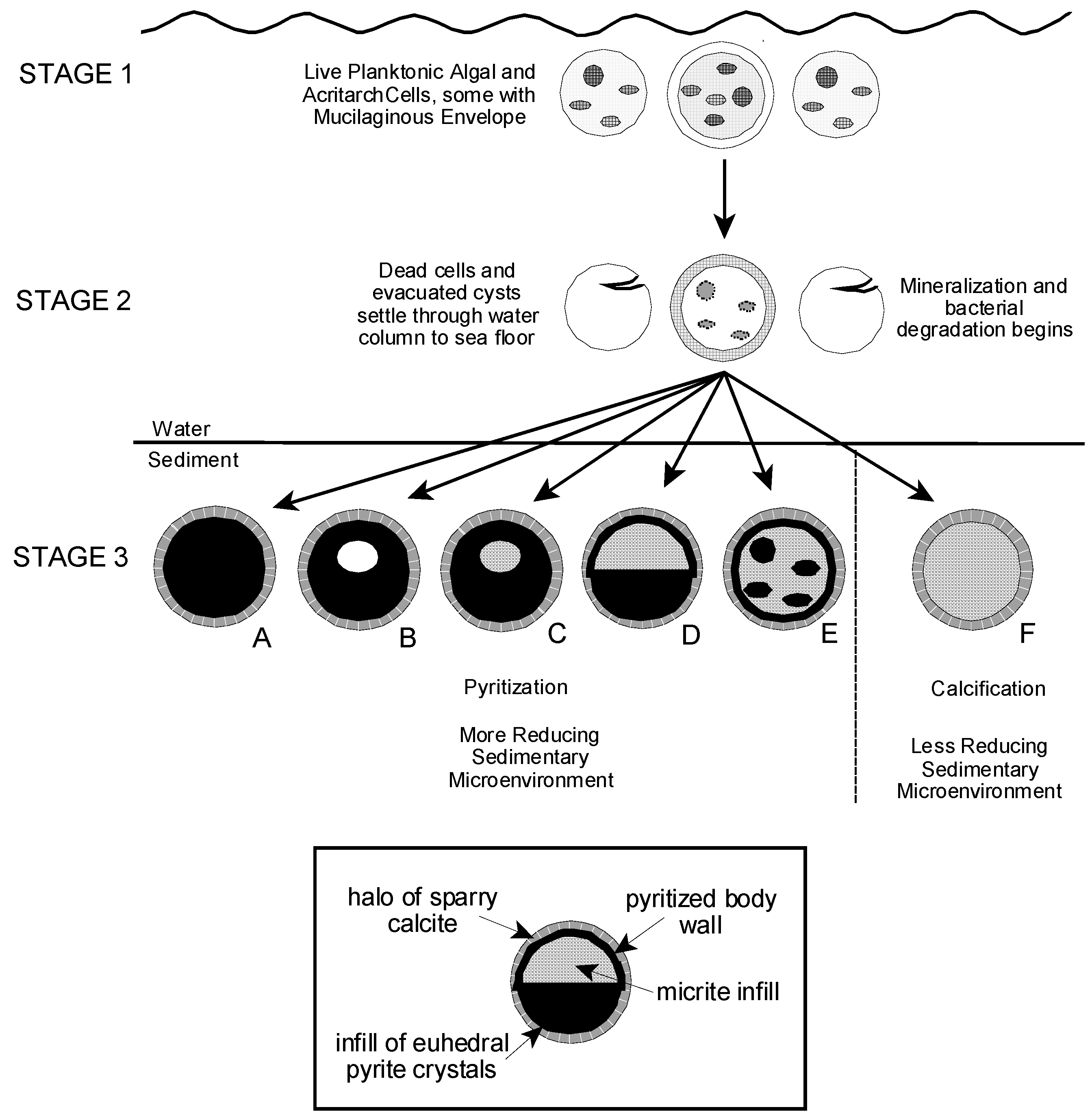
6.1. Algal and AcritarchTaphonomy
6.2. Preservation of the Tully Microflora
6.3. Rate of Mineralization
6.4. Depositional Environment of the Microfossil Horizon
6.5. Paleoecology of the TullyMicrobiota
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Williamson, W.C. On the organisation of the fossil plants of the Coal-Measures, Part X. Including an examination of the supposed radiolarians of the Carboniferous rocks. Philos. Trans. R. Soc. Lond. Ser. B 1881, 148, 9–43. [Google Scholar]
- Kaźmierczk, J.; Kremer, B. Early post-mortem calcified Devonian acritarchs as a source of calcispheric structures. Facies 2005, 51, 573–584. [Google Scholar] [CrossRef]
- Thomas, H.D. Origin of spheres in the Georgetown Limestone. J. Paleontol. 1932, 6, 100–101. [Google Scholar]
- Stanton, R.J., Jr. Radiosphaerid calcispheres in North America and remarks on calcisphere classification. Micropaleontology 1967, 13, 465–472. [Google Scholar] [CrossRef]
- Masters, B.A.; Scot, R.W. Calcispheres and nannoconids. In The Encyclopedia of Paleontology; Fairbridge, R.W., Jablonski, D., Eds.; Dowden, Hutchinson and Ross: Stroudsburg, PA, USA, 1979; pp. 167–170. [Google Scholar]
- Wanner, J. Gesteinsbeldende Foraminiferen aus Malm und Unterkreide des östlichen Ostendischen Archipels. Paläontol. Zeits 1940, 22, 75–99. [Google Scholar]
- Kettenbrink, E.C., Jr.; Toomey, D.F. Distribution and paleoecological implication of calcareous Foraminifera in the Devonian Cedar Valley Formation of Iowa. J. Foraminifer. Res. 1975, 5, 176–187. [Google Scholar] [CrossRef]
- Cayuex, L. Les Calcisphères typiques sont des Algues siphonées. C. R. Acad. Sci. Paris 1929, 188, 594–597. [Google Scholar]
- Lombard, A.; Monteyne, R. Calcisphères dans le Frasnien de Bois-de-Villiers (Namur). Bull. Soc. Belg. Géol. Paléont. Hydrol. 1952, 61, 13–25. [Google Scholar]
- Aldridge, R.J.; Armstrong, H. Spherical phosphatic microfossils from the Silurian of North Greenland. Nature 1981, 292, 531–533. [Google Scholar] [CrossRef]
- Holmer, L.E. Ordovician mazuelloids and other microfossils from Västergötland. Geologiska Föreningen i Stockholm Förhandandlingur 1986, 109, 66–71. [Google Scholar] [CrossRef]
- Kozur, H.W. Muellerisphaerida, eine neue Ordnung von Mikofossilien unbekannter sytematischer Stellung aus dem Silur und Unterdevon von Ungarn. Geol. Paläontol. Mittelungen Innsbruck 1984, 13, 125–148. [Google Scholar]
- Porebska, E.; Koszlowska, E. Mazzuelloids—Apatite algae from the Lower Paleozoic of the Bardzkie Mountains (Sudety Mountains, SW Poland). Przeglad Geologiczny 2001, 49, 1050–1060. [Google Scholar]
- Loydell, D.K.; McMillan, I.; Barron, H.F. Muellerisphaerids from the Llandovery of western mid-Wales. J. Micropaleontol. 1988, 7, 243–246. [Google Scholar] [CrossRef]
- Tibbs, S.L.; Briggs, D.E. G.; Prössl, K. Pyritisation of plant microfossils from the Devonian Hunsrück Slate of Germany. Palaöntol. Z. 2003, 77, 241–246. [Google Scholar] [CrossRef]
- Schieber, J.E.; Baird, G. On the origin and significance of pyrite spheres in Devonian black shales of North America. J. Sediment. Res. 2001, 71, 155–166. [Google Scholar] [CrossRef]
- Kaźmierczak, J.; Ittekot, V.; Degens, E.T. Biocalcification through time environmental challenge and cellular response. Paläontol. Z. 1985, 59, 15–33. [Google Scholar] [CrossRef]
- Kremer, B. Mazuelloids: Products of post-mortem phosphatization of acanthomorphic acritarchs. Palaios 2005, 20, 27–36. [Google Scholar] [CrossRef]
- Evitt, W.R. A discussion and proposals concerning fossil dinoflagellates, hytrichospheres, and acritarchs, I. Proc. Natl. Acad. Sci. USA 1963, 48, 158–164. [Google Scholar] [CrossRef]
- Evitt, W.R. A discussion and proposals concerning fossil dinoflagellates, hytrichospheres, and acritarchs, II. Proc. Natl. Acad. Sci. USA 1963, 48, 298–302. [Google Scholar] [CrossRef]
- Martin, F. Acritarchs: A review. Biol. Rev. 1993, 68, 475–538. [Google Scholar] [CrossRef]
- Colbath, G.K.; Grenfell, H.R. Review of biological affinities of Paleozoic acid-resistant, organic-walled eukaryotic algal microfossils (including acritarchs). Rev. Palaeobot. Palynol. 1995, 86, 287–314. [Google Scholar] [CrossRef]
- Kempe, A.; Schopf, J.W.; Altermann, W.; Kudryavtsev, A.B.; Heckl, W.M. Atomic force microscopy of Precambrian microscopic fossils. Proc. Natl. Acad. Sci. USA 2002, 99, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Kempe, A.; Wirth, A.; Altermann, W.; Stark, R.W.; Schopf, J.W.; Heckl, W.M. Focused ion beam preparation: An in situ nanoscopic study of Precambrian acritarchs. Precambr. Res. 2005, 140, 36–54. [Google Scholar] [CrossRef]
- Marshall, C.P.; Javaux, E.J.; Knoll, A.H.; Walter, M.R. Combined micro-Fourier transform infrared (FTIR) spectroscopy and micro-Raman spectroscopy of Precambrian acritarchs: A new approach to paleobiology. Precambr. Res. 2005, 138, 208–224. [Google Scholar] [CrossRef]
- Kaźmierczak, J.; Kremer, B. Spore-like bodies in some early Paleozoic acritarchs: Clues to chlorococcalean affinities. Acta Palaeontol. Pol. 2009, 54, 541–551. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Sergeev, V.N. Confocal laser scanning microscopy and Raman imagery of the Late Neoproterozoic Chichkan microbiota of south Kazakhstan. J. Paleontol. 2010, 84, 402–416. [Google Scholar] [CrossRef]
- Moczydƚowska, M.; Landing, E.; Zang, W. Palacios, T. Proterozoic plankton and timing of Chlorophyte origins. Palaeontology 2011, 54, 721–734. [Google Scholar] [CrossRef]
- Cohen, P.; Knoll, A.; Kodner, R. Large spinose microfossils in Ediacaran rocks as resting stages of early animals. Proc. Nat. Acad. Sci. USA 2009, 106, 6519–6524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, P.; Kelly, A.; Kodner, R. Taxonomic affinity of Late Devonian organic-walled microfossils: Interpretations and implications. Geol. Soc. Am. Abstr. Annu. Meet. 2016, 48. [Google Scholar] [CrossRef]
- Grabau, A.W. Stratigraphic relationships of the Tully limestone and the Genesee shale in eastern North America. Geol. Soc. Am. Bull. 1917, 28, 945–958. [Google Scholar]
- Trainer, D.W., Jr. The Tully Limestone of central New York. N. Y. State Mus. Bull. 1932, 291, 1–43. [Google Scholar]
- Willard, B. Hamilton Group of central Pennsylvania. Geol. Soc. Am. Bull. 1935, 46, 195–224. [Google Scholar] [CrossRef]
- Willard, B. Middle and Upper Devonian. In The Devonian of Pennsylvania: Pennsylvania Geol. Survey; 4th Series; General Geol. Rp.t G19; Willard, B., Swartz, F.M., Cleaves, A.B., Eds.; Pennsylvania Geological Survey: Harrisburg, PA, USA, 1939; pp. 131–308. [Google Scholar]
- Johnson, K.G.; Friedman, G.M. The Tully clastic correlative (upper Devonian) of New York State: A model for recognition of alluvial, dune (?), tidal, nearshore (bar and lagoon), and offshore sedimentary environments in a tectonic delta complex. J. Sediment. Res. 1969, 39, 452–485. [Google Scholar]
- Heckel, P.H. Devonian Tully Limestone in Pennsylvanian and comparison of type Tully Limestone of New York. Pa. Geol. Surv. Bull. 4th Ser. 1969, IC60, 1–33. [Google Scholar]
- Heckel, P.H. Nature, Origin, and Significance of the Tully Limestone; Geological Society of America Special Paper 138; Geological Society of America: Boulder, CO, USA, 1973; p. 244. [Google Scholar]
- Baird, G.C.; Brett, C.E. Shelf and off-shelf deposits of the Tully Formation in New York and Pennsylvania: Faunal incursions, eustacy, and tectonics. Courier-Forschungsinstitut Senckenberg 2003, 242, 141–156. [Google Scholar]
- Baird, G.C.; Brett, C.E. Late Givetian Taghanic bioevents in New York State: New discoveries and questions. Bull. Geosci. 2008, 83, 357–370. [Google Scholar] [CrossRef]
- Kirchgasser, W.T.; Baird, G.C.; Brett, C.E. Regional placement of the Middle/Upper Devonian (Givetian-Frasnian) boundary in western New York State. In Devonian of the World; Memoir 14, Part 2; McMillan, N.J., Embry, A.F., Glass, D.J., Eds.; Canadian Society of Petroleum Geologists: Calgary, AB, Canada, 1988; pp. 113–118. [Google Scholar]
- Brett, C.E.; Baird, G.C. B-5 depositional sequences, cycles, and foreland basin dynamics in the late Middle Devonian (Givetian) of the Genesee Valley and western Finger Lakes region. In Field Trip Guidebook for New York State Geological Association 66th Annual Meeting; Brett, C.E., Scatterday, J., Eds.; New York State Geological Association: Albany, NY, USA, 1994; pp. 505–586. [Google Scholar]
- Weary, D.J.; Harris, A.G. Early Frasnian (Late Devonian) conodonts from the Harrell Shale, western foreland fold-and-thrust belt, West Virginia, Maryland, and Pennsylvania Appalachians, U.S.A. Cour. Forsch. Inst. Senckenburg 1994, 168, 195–225. [Google Scholar]
- Brown, J.O. Biostratigraphy and Paleoecology of the Givetian Hamilton Group in Pennsylvania and New York. Ph.D. Thesis, City University of New York, New York, NY, USA, 2001. [Google Scholar]
- Zambito, J.J., IV; Brett, C.E.; Baird, G.C. The Late Middle Devonian (Givetian) Global Taghanic biocrisis in its type area (Northern Appalachian Basin): Geologically rapid faunal transitions driven by global and local environmental changes. In Earth and Life: International Year of Planet Earth; Talent, J., Ed.; Springer Science + Business Media B.V.: Dordrecht, The Netherlands, 2012; pp. 677–702. [Google Scholar]
- Johnson, J.G. Taghnanic onlap and the end of North American Devonian provinciality. Geol. Soc. Am. Bull. 1970, 81, 2077–2105. [Google Scholar] [CrossRef]
- Brett, C.E.; Baird, G.C.; Bartholomew, A.J. Biofacies recurrence in the Middle Devonian of New York State: An example with implications for evolutionary paleoecology. Palaios 2007, 22, 306–324. [Google Scholar] [CrossRef]
- Day, J. Faunal signatures of Middle-Upper Devonian depositional sequences and sea-level fluctuations in the Iowa Basin. In Paleozoic Sequence Stratigraphy: Views from the North American Craton; Geological Society of America Special Paper 306; Witzke, B.J., Ludvigson, G.A., Day, J., Eds.; Geological Society of America: Boulder, CO, USA, 1996; pp. 277–300. [Google Scholar]
- House, M.R. Strength, timing, setting and cause of the mid-Paleozoic extinctions. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 181, 5–25. [Google Scholar] [CrossRef]
- Aboussalam, S.Z. Das “Taghanic-Event” in höheren Mittel-Devonien von West-Europa und Marokko. Münstersche Forsch. Geol. Paläontol. 2003, 97, 1–322. [Google Scholar]
- Aboussalam, S.Z.; Becker, R.T. The global Taghanic Biocrisis (Givetian) in the eastern Anti-Atlas, Morocco. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 304, 136–164. [Google Scholar] [CrossRef]
- Harris, A.G.; Sweet, W.C. Mechanical and chemical techniques for separating microfossils from rock, sediment, and residue matrix. In Paleotechniques; Paleontological Society Special Paper 4; Feldman, R.M., Chapman, R.E., Hannibal, J.T., Eds.; The Paleontological Society: Boulder, CO, USA, 1989; pp. 70–86. [Google Scholar]
- Maples, C.G.; Waters, J.A. Use of drain cleaner in processing shale samples. J. Paleontol. 1990, 64, 484–485. [Google Scholar] [CrossRef]
- Pojeta, J., Jr.; Balanc, M. Heating and quenching of fossils. In Paleotechniques; Paleontological Society Special Paper 4; Feldman, R.M., Chapman, R.E., Hannibal, J.T., Eds.; The Paleontological Society: Boulder, CO, USA, 1989; pp. 218–222. [Google Scholar]
- Krukowski, S.T. Sodium metatungstate: A new heavy-mineral separation medium for the extraction of conodonts from insoluble residues. J. Paleontol. 1988, 62, 314–316. [Google Scholar] [CrossRef]
- Schieber, J. The role of an organic slime matrix in the formation of pyritized burrow trails and pyrite concretions. Palaios 2002, 17, 104–109. [Google Scholar] [CrossRef]
- Winslow, M.R. Plant Spores and Other Microfossils from Upper Devonian and Lower Mississippian Rocks of Ohio; United States Geological Survey Professional Paper 364; United States Geological Survey: Reston, VA, USA, 1962; p. 90.
- Grey, K.; Willman, S. Taphonomy of Ediacaran acritarchs from Australia: Significance for taxonomy and biostratigraphy. Palaios 2009, 24, 239–256. [Google Scholar] [CrossRef]
- Burden, E.T.; Quinn, L.; Nowlan, G.S.; Bailey-Nill, L.A. Palynology and micropaleontology of the Clam Bank Formation (Lower Devonian of western Newfoundland, Canada. Palynology 2002, 26, 185–215. [Google Scholar] [CrossRef]
- Colbath, G.K. Fossil prasinophycean phycomata (Chlorophyta) from the Silurian Bainbridge Formation, Silurian, USA. Phycologia 1983, 22, 249–265. [Google Scholar] [CrossRef]
- Filipiak, P. Late Devonian and Early Carboniferous acritarchs and prasinophytes from the Holy Cross Mountains (central Poland). Rev. Paleobot. Palynol. 2005, 134, 1–26. [Google Scholar] [CrossRef]
- Agić, H.; Moczydłowska, M.; Yin, L.-M. Affinity, life cycle, and intracellular complexity of organic-walled microfossils from the Mesoproterozoic of Shanxi, China. J. Paleontol. 2015, 89, 28–50. [Google Scholar] [CrossRef]
- Strother, P.K. Acritarchs. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Dallas, TX, USA, 1996; Volume 1, pp. 81–106. [Google Scholar]
- Filipiak, P. Lower Famennian phytoplankton from the Holy Cross Mountains, Central Poland. Rev. Paleobot. Palynol. 2009, 157, 326–338. [Google Scholar] [CrossRef]
- Ghavidel-Syooki, M.; Hassanzadeh, J.; Vecoli, M. Palynology and isotope geochronology of the Upper Ordovician–Silurian successions (Ghelli and Soltan Maidan Formations) in the Khoshyeilagh area, eastern Alborz Range, northern Iran; stratigraphic and palaeogeographic implications. Rev. Paleobot. Palynol. 2011, 164, 251–271. [Google Scholar] [CrossRef]
- Miller, M.A.; Williams, G.L. Velatasphaera hudsonii gen. et sp. nov., an Ordovician acritarch from Hudson Strait, North West Territories, Canada. Palynology 1988, 12, 121–127. [Google Scholar] [CrossRef]
- Fritsch, F.E. The Structure and Reproduction of the Algae; Cambridge University Press: Cambridge, UK, 1965; p. 791. [Google Scholar]
- Boney, A.D. Mucilage: The ubiquitous algal attribute. Br. Phycol. J. 1981, 16, 115–132. [Google Scholar] [CrossRef]
- Oertel, A.; Aichinger, N.; Hochrieter, R.; Thalhamer, J.; Ursula Lütz, U. Analysis of mucilage secretion and excretion in Micrasterias (Chlorophyta) by means of immunoelectron microscopy and digital time lapse video microscopy. J. Phycol. 2004, 40, 711–720. [Google Scholar] [CrossRef]
- Kreinitz, L.; Hegewald, E.H.; Hepperle, D.; Huss, V.A.R.; Rohr, T.; Wolf, F. Phylogenetic relationships of Chlorella and Parachlorella gen. nov. (Clorophyta, Trebouxiophyceae). Phycologia 2004, 43, 529–542. [Google Scholar] [CrossRef]
- Moldowan, J.M.; Talyzina, N.M. Biogeochemical evidences for dinoflagellate ancestors in the early Cambrian. Science 1998, 281, 1168–1170. [Google Scholar] [CrossRef] [PubMed]
- Arouri, K.; Greenwood, P.F.; Walter, M.R. A possible chlorophyacean affinity of some Neoproterozoic acritarchs. Org. Geochem. 1999, 30, 1323–1337. [Google Scholar] [CrossRef]
- Javaux, E.J.; Marshall, C.P. A new approach in deciphering early protist paleobiology and evolution: Combined microscopy and microchemistry of single Proterozoic acritarchs. Rev. Paleobot. Palynol. 2006, 139, 1–15. [Google Scholar] [CrossRef]
- Kodner, R.B.; Pearson, A.; Summons, R.E.; Knoll, A.H. Sterols in red and green algae: Quantification, phylogeny, and relevance for the interpretation of geologic steranes. Geobiology 2008, 6, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Moczydƚowska, M. Life cycle of early Cambrian microalgae from the Skiagia-plexus acritarchs. J. Paleontol. 2010, 84, 216–230. [Google Scholar] [CrossRef]
- Lewis, L.A.; McCourt, R. Green algae and the origin of land plants. Am. J. Bot. 2004, 91, 1535–1556. [Google Scholar] [CrossRef] [PubMed]
- Adl, S.M.; Alastair, G.B.; Simpson, M.A.; Farmer, R.A.; Andersen, O.R.; Barta, J.R.; Bowser, S.S.; Brugerolle, G.; Fensome, R.A.; Fredericq, S.; et al. The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. J. Eukaryot. Biol. 2005, 52, 399–451. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Ezpeleteta, N.; Brinkmsann, H.N.; Burger, G.; Roger, A.J.; Gray, M.W.; Herve, P.; Lan, B.F. Toward resolving the eukaryotic tree: The phylogenetic positions of jakobids and cercozoans. Curr. Biol. 2005, 17, 1420–1425. [Google Scholar] [CrossRef] [PubMed]
- O’Kelly, C.J. The origin and early evolution of green plants. In Evolution of Primary Producers in the Sea; Falkowski, P.G., Knoll, A.H., Eds.; Academic Press Elsevier: Amsterdam, The Netherlands, 2007; pp. 287–309. [Google Scholar]
- Termel, M.; Brouard, J.-S.; Gagnon, C.; Otis, C.; Lemieux, C. Deep division in the Chlorophyceae (Chlorophyta) revealed by Chloroplast phylogenomic analyses. J. Phycol. 2008, 44, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Wicander, E.R.; Schopf, J.W. Microorganisms from the Kalkberg Limestone (Lower Devonian) of New York State. J. Paleontol. 1974, 48, 74–77. [Google Scholar]
- Wood, G.D.; Clendening, J.A. Organic-Walled Microphytoplankton and Chitinozoans from the Middle Devonian (Givetian) Boyle Dolomite of Kentucky, U.S.A. Palynology 1985, 9, 133–145. [Google Scholar] [CrossRef]
- Knoll, A.H.; Golubic, S. Anatomy and taphonomy of a Precambrian algal stromatolite. Precambr. Res. 1979, 10, 115–151. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. A revised six-kingdom system of life. Biol. Rev. 1998, 73, 203–266. [Google Scholar] [CrossRef] [PubMed]
- Tappan, H. The Paleobiology of Plant Protists; W.H. Freeman & Co.: San Francisco, CA, USA, 1980; p. 1028. [Google Scholar]
- Newton, E.T. On “tasmanite” and Australian “white coal”. Geol. Mag. Ser. 2 1875, 12, 337–342. [Google Scholar] [CrossRef]
- Schopf, J.M.; Wilson, L.R.; Bentall, R. An Annotated Synopsis of Paleozoic Fossil Spores and the Definition of Generic Groups; Illinois State Geological Survey, Report of Investigations 91; State of Illinois, Division of the Geological Survey: Urbana, IL, USA, 1944; p. 74.
- Knoll, A.H.; Swett, K. Micropalaeontology of the Late Proterozoic Veteranen Group, Spitsbergen. Palaeontology 1985, 28, 451–473. [Google Scholar]
- Samuelsson, J.; Dawes, P.R.; Vidal, G. Organic-walled microfossils from the Proterozoic Thule Supergroup, Northwest Greenland. Precambr. Res. 1999, 96, 1–23. [Google Scholar] [CrossRef]
- Boalch, G.T.; Guy-Ohlson, G. Tasmanites, the correct name for Pachysphaera (Prasinophyceae, Pterospermataceae). Taxonomy 1992, 41, 529–531. [Google Scholar] [CrossRef]
- Fensome, R.A.; Williams, G.L.; Barss, M.S.; Freeman, J.M.; Hill, J.M. Acritarchs and Fossil Prasinophytes: An Index to Genera, Species, and Intraspecific Taxa; American Association of Stratigraphic Palynologists Foundation Contributions Series No. 25; American Association of Stratigraphic Palynologists Foundation: Houston, TX, USA, 1990; p. 771. [Google Scholar]
- Mullins, G.L.; Aldridge, R.J.; Dorning, K.J.; LeE Herisse, A.; Jun, L.; Moczydƚwska-Vidal, M.; Molyneux, S.G.; Servais, T.; Wicander, R. The PhytoPal Taxonomic Database (Taxon List). 2007. Available online: www.le.ac.uk/geology/glm2/phytopal/taxa.pdf (accessed on 27 December 2014).
- Wicander, E.R. A Catalog and Biostratigraphic Distribution of North American Devonian Acritarchs; American Association of Stratigraphic Palynologists Foundation Contributions Series No. 10; American Association of Stratigraphic Palynologists Foundation: Houston, TX, USA, 1983; p. 133. [Google Scholar]
- Downie, C.; Evitt, W.R.; Sarjeant, W.A.S. Dinoflagellates, hystrichospheres and the classification of the acritarchs. Stanf. Univ. Publ. Geol. Sci. 1963, 7, 1–16. [Google Scholar]
- Staplin, F.L.; Jansonius, J.; Pocock, S.A.J. Evaluation of some acritarchous hystrichosphere genera. Neues Jahrb. Geol. Paläontol. Abhand. 1965, 123, 167–201. [Google Scholar]
- Sarjeant, W.A.S. Microplankton from the Upper Callovian and Lower Oxfordian of Normandy. Rev. Micropaleontol. 1968, 10, 221–242. [Google Scholar]
- Moczydƚowska, M. Cambrian acritarchs from Upper Silesia, Poland—biochronology and tectonic implications. Foss. Strat. 1998, 46, 1–121. [Google Scholar]
- Deflandre, G. Microplancton des mers jurassiques conservé dans les marnes de Villers-sur-Mer (Calvados). Étude liminaire et considérations générales. Travaux Station Zoologique Wimereux 1938, 13, 147–200. [Google Scholar]
- Moczydlowska, M. Acritarch biostratigraphy of the Lower Cambrian and the Precambrian-Cambrian boundary in southeastern Poland. Foss. Strat. 1991, 29, 1–127. [Google Scholar]
- Moczydƚowska, M.; Stockfors, M. Acritarchs from the Cambrian-Ordovician Boundary Interval on Kolguev Island, Arctic Russia. Palynology 2004, 28, 15–73. [Google Scholar] [CrossRef]
- Heisecke, A.M. Microplancton de la Formación Roca de la Provincia de Neuquén. Ameghiniana 1970, 7, 225–263. [Google Scholar]
- Wicander, E.R. Upper Devonian-Lower Mississippian acritarchs and prasinophycean algae from Ohio, U.S.A. Palaeontogr. Abt. B 1974, 148, 9–43. [Google Scholar]
- Wicander, E.R.; Playford, G. Acritarchs and Spores from the Upper Devonian Lime Creek Formation, Iowa, U.S.A. Micropaleontology 1985, 31, 97–138. [Google Scholar] [CrossRef]
- Wicander, R.; Wood, G.D. The use of microplankton and chinozoa for interpreting transgressive/regressive cycles in the Rapid Member of the Cedar Valley Formation (Middle Devonian) Iowa. Rev. Palaeobot. Palynol. 1997, 98, 125–152. [Google Scholar] [CrossRef]
- Smelror, M.; Leereveld, H. Dinoflagellate and acritarch assemblages from theLate Bathonian to Early Oxfordian of Montagne Crussol, Rhône Valley, Southern France. Palynology 1989, 13, 121–141. [Google Scholar] [CrossRef]
- Brown, J.O.; Chamberlain, J.A., Jr.; Perlmutter, B. A pyritic micro-molluscan fauna atthe base of the Lower Tully Formation (upper Givetian) from Lock Haven, Pennsylvania. Geol. Soc. Am. Abstr. Program Annu. Meet. 1998, 30, A-34. [Google Scholar]
- Chamberlain, J.A., Jr.; Brown, J.O. First occurrence of the enigmatic microfossil Jinonicella in North America: Tully Limestone (Givetian) of central Pennsylvania. Geol. Soc. Am. Abstr. Program Annu. Meet. 2000, 32, A-10. [Google Scholar]
- Bailey, J.B. Systematics, Hinge, and Internal Morphology of the Devonian Bivalve, Nuculoidea corbuliformis (Hall and Whitfield). J. Paleontol. 1986, 60, 1177–1185. [Google Scholar] [CrossRef]
- Linsley, D.M. Devonian Paleontology of New York; Special Publication 21; Paleontological Research Institution: Ithaca, NY, USA, 1994; p. 472. [Google Scholar]
- Brett, C.E.; Cottrell, J.F. Substrate specificity in the Devonian tabulate coral Pleurodictyum. Lethaia 1982, 15, 247–262. [Google Scholar] [CrossRef]
- Flower, R.H. Classification of Devonian Nautiloids. Am. Midl. Nat. 1945, 33, 675–724. [Google Scholar] [CrossRef]
- Tillman, J.R. Ostracodes of the Superfamilies Beyrichiacea and Drepanellacea from Middle Devonian Rocks of Central Ohio. J. Paleontol. 1984, 58, 234–253. [Google Scholar]
- Lindemann, R.H.; Yochelson, E.L. Viriatellina (Dacryoconarida) from the Middle Devonian Ludlowville Formation at Alden, New York. J. Paleontol. 1992, 66, 93–199. [Google Scholar] [CrossRef]
- Lindemann, R.H.; Skidmore College, Saratoga Springs, NY, USA. Personal communication, 2014.
- Ulrich, E.O.; Bassler, R.S. A revision of the Paleozoic Bryozoa, Part 1. On genera and species of Ctenostomata. Smithson. Misc. Collect. Q. Issue 1904, 2, 256–298. [Google Scholar]
- Bassler, R.S. Treatise on Invertebrate Paleontology. In Part G, Bryozoa; Moore, R.C., Ed.; Geological Society of America: Boulder, CO, USA; University of Kansas of Kansas Press: Lawrence, KS, USA, 1953; p. 253. [Google Scholar]
- Stumm, E.C.; Wright, J.D. Check list of fossil invertebrates described from the Middle Devonian rocks of the Thedford-Arkona Region of southwestern Ontario. Contrib. Mus. Paleontol. Univ. Mich. 1958, 14, 81–132. [Google Scholar]
- Olempska, E. Exceptional soft-tissue preservation in boring ctenostome bryozoans and associated “fungal” borings from the Early Devonian of Podolia, Ukraine. Acta Palaeontol. Pol. 2012, 57, 925–940. [Google Scholar] [CrossRef]
- Wilson, M.A.; Taylor, P.D. The Paleozoic encrusting sclerobionts Allonema and Ascodictyon: Component parts of organisms belonging to the same problematic group. Geol. Soc. Am. Abstr. Program Annu. Meet. 2013, 45, 300–311. [Google Scholar]
- Wilson, M.A.; Taylor, P.D. The morphology and affinities of Allonema and Ascodictyon, two abundant Palaeozoic encrusters commonly misattributed to the ctenostome bryozoans. Studi Trentini Scienze Naturali 2014, 94, 259–266. [Google Scholar]
- Wilson, M.A.; Palmer, T.J. Patterns in the Ordovician bioerosion revolution. Ichnos 2006, 13, 109–112. [Google Scholar] [CrossRef]
- Taylor, P.D. Preservation of soft-bodied and other organisms by bioimmuration—A review. Palaeontology 1990, 33, 1–17. [Google Scholar]
- Todd, J.A.; Taylor, P.D.; Favorskaya, T.A. A bioimmured ctenostome bryozoan from the early Cretaceous of the Crimea and the new genus Simplicidium. Geobios 1997, 30, 205–213. [Google Scholar] [CrossRef]
- Todd, J.A.; Hagdorn, N.H. First record of Muschelkalk Bryozoa: The earliest ctenostome body fossils. In Muschelkalk. Schöntaler Symposium 1991; Hagdorn, H., Seilacher, A., Eds.; Sonderbände der Gesellschaft für Naturkunde in Würtemburg 2; Goldschneck: Stuttgart, Germany, 1993; pp. 285–286. [Google Scholar]
- Peel, J.S.; Jeppson, L. The problematic fossil Jinonicella from the Lower Silurian of Gotland. GFF 2006, 28, 39–42. [Google Scholar] [CrossRef]
- Pokorny, V. Jinonicellina, a new suborder of presumed Archaeogastropoda. Bull. Czech Geol. Surv. 1978, 53, 39–42. [Google Scholar]
- Dzik, J. Larval development, musculature, and relationships of Sinuitopsis andrelated Baltic bellerophonts. Norsk Geol. Tidsskrift 1981, 61, 111–121. [Google Scholar]
- Dzik, J. Machaeridians, chitons, and conchiferan molluscs of the Mójcza Limestone. Acta Palaeontol. Pol. 1994, 53, 213–252. [Google Scholar]
- Peel, J.S. Scaphopodization in Palaeozoic mollusks. Palaeontology. 2006, 49, 1357–1364. [Google Scholar] [CrossRef]
- Fryda, J. Taxonomic position of suborder Jinonicellina. Bull. Czech Geol. Surv. 1999, 74, 27–29. [Google Scholar]
- Yochelson, E.L. Comments on Janospira. Lethaia 1977, 10, 204. [Google Scholar] [CrossRef]
- Love, L.G.; Zimmerman, D.O. Bedded pyrite and micro-organisms from the Mount Isa Shale. Econ. Geol. 1961, 56, 873–896. [Google Scholar] [CrossRef]
- Love, L.G.; Murray, J.W. Biogenic pyrite in recent sediments of Christchurch Harbour, England. Am. J. Sci. 1983, 261, 433–448. [Google Scholar] [CrossRef]
- Kohn, M.J.; Riciputi, L.R.; Stakes, D.; Orange, D.L. Sulfur isotope variability in biogenic pyrite: Reflections of heterogeneous bacterial colonization. Am. Mineral. 1998, 83, 1454–1468. [Google Scholar] [CrossRef]
- Folk, R. Nannobacteria and the formation of framboidal pyrite: Textural evidence. J. Earth Syst. Sci. 2005, 114, 369–374. [Google Scholar] [CrossRef]
- Maclean, L.C.W.; Tyliszczak, T.; Gilbert, P.U.P.A.; Zhou, D.; Pray, T.J.; Onstott, T.C.; Southam, G. A high-resolution chemical and structural study of framboidal pyrite within a low-temperature bacterial biofilm. Geobiology 2008, 6, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Love, L.G. Early diagenetic iron sulphide in recent sediments of the Wash (England). Sedimentology 1967, 9, 327–352. [Google Scholar] [CrossRef]
- Lein, A.Y. Formation of carbonate and sulfide minerals during diagenesis of reduced sediment. In Environmental Biogeochemistry and Geomicrobiology; Krumbein, W.E., Ed.; Ann Arbor Science: Ann Arbor, MI, USA, 1978; pp. 339–354. [Google Scholar]
- Allison, P.A. The role of anoxia in the decay and mineralization of proteinaceous macro-fossils. Paleobiology 1988, 14, 139–154. [Google Scholar] [CrossRef]
- Rickard, D.T.; Schoonen, M.A.A.; Luther, G.W. Chemistry of iron sulfides in sedimentary environments. In Geochemical Transformations of Sedimentary Sulfur; American Chemical Society Symposium Series, v. 612; Vairavamurthy, M.A., Schoonen, M.A.A., Eds.; American Chemical Society: Washington, DC, USA, 1995; pp. 168–193. [Google Scholar]
- Wilkin, R.T.; Barnes, H.L. Pyrite formation by reactions of iron monosulfides with dissolved inorganic and organic sulfur species. Geochim. Cosmochim. Acta 1996, 60, 4167–4179. [Google Scholar] [CrossRef]
- Wilkin, R.T.; Barnes, H.L. Formation processes of framboidal pyrite. Geochim. Cosmochim. Acta 1997, 61, 323–339. [Google Scholar] [CrossRef]
- Ettensohn, F.R. Controls on development of Catskill Delta complex basin-facies. In The Catskill Delta; Geological Society of America Special Paper 201; Woodrow, D.L., Sevon, W.D., Eds.; Geological Society of America: Boulder, CO, USA, 1985; pp. 65–77. [Google Scholar]
- Stasiuk, L.D.; Fowler, M.G. Organic facies in Devonian and Mississippian strata of Western Canada Sedimentary Basin: Relation to kerogen type, paleoenvironment, and paleogeography. Bull. Can. Petrol. Geol. 2004, 52, 234–255. [Google Scholar]
- Baird, G.C.; Brett, C.E. Late Middle Devonian (Late Givetian) Tully Formation in Pennsylvania: Comparison with Tully Limestone and equivalent clastic deposits in New York State. Geol. Soc. Am. Abstr. Program Annu. Meet. 2007, 39, 39. [Google Scholar]
- Wignall, P.B.; Hallam, A. Biofacies, stratigraphic distribution and depositional models of British onshore Jurassic black shales. In Modern and Ancient Continental Shelf Dysoxia; Special Publication 58; Tyson, R.V., Pearson, T.H., Eds.; Geologocal Society London: London, UK, 1991; pp. 291–309. [Google Scholar]
- Allison, P.A.; Wignall, P.B.; Brett, C.A. Palaeo-oxygenation: Effects and recognition. In Marine Palaeoenvironmental Analysis from Fossils; Special Publication 83; Bosence, D.W.J., Allison, P.A., Eds.; Geological Society London: London, UK, 1995; pp. 97–112. [Google Scholar]
- Boyer, D.I.; Droser, M.I. Palaeoecological patterns within the dysaerobic biofacies: Examples from Devonian black shales of New York State. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 276, 206–216. [Google Scholar] [CrossRef]
- Mapes, R.H.; Nützel, A. Late Palaeozoic mollusc reproduction: Cephalopod egg-laying behavior and gastropod larval palaeobiology. Lethaia 2009, 42, 341–356. [Google Scholar] [CrossRef]
- Arnold, J.M.; Carlson, B.A. Living Nautilus embryos: Preliminary observations. Science 1986, 232, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Okubo, S.; Tsujii, T.; Watabe, N.; Williams, D.F. Hatching of Nautilus belauensis Saunders 1981, in captivity: Culture, growth and stable isotope compositions of shells, and histology and immunohistochemistry of the mantle epithelium of the juveniles. Veliger 1995, 38, 192–202. [Google Scholar]



© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chamberlain, J.A.; Chamberlain, R.B.; Brown, J.O. A Mineralized Alga and Acritarch Dominated Microbiota from the Tully Formation (Givetian) of Pennsylvania, USA. Geosciences 2016, 6, 57. https://doi.org/10.3390/geosciences6040057
Chamberlain JA, Chamberlain RB, Brown JO. A Mineralized Alga and Acritarch Dominated Microbiota from the Tully Formation (Givetian) of Pennsylvania, USA. Geosciences. 2016; 6(4):57. https://doi.org/10.3390/geosciences6040057
Chicago/Turabian StyleChamberlain, John A., Rebecca B. Chamberlain, and James O. Brown. 2016. "A Mineralized Alga and Acritarch Dominated Microbiota from the Tully Formation (Givetian) of Pennsylvania, USA" Geosciences 6, no. 4: 57. https://doi.org/10.3390/geosciences6040057