Use of Residual Biomass from the Textile Industry as Carbon Source for Production of a Low-Molecular-Weight Xylanase from Aspergillus oryzae

Abstract
:1. Introduction
2. Experimental Section
2.1. Feed Residue, Substrate and Chemicals
2.2. Residue Pretreatment
2.3. Organism and Enzyme Production
2.4. Enzyme Assays
2.5. Determination of Protein Concentration
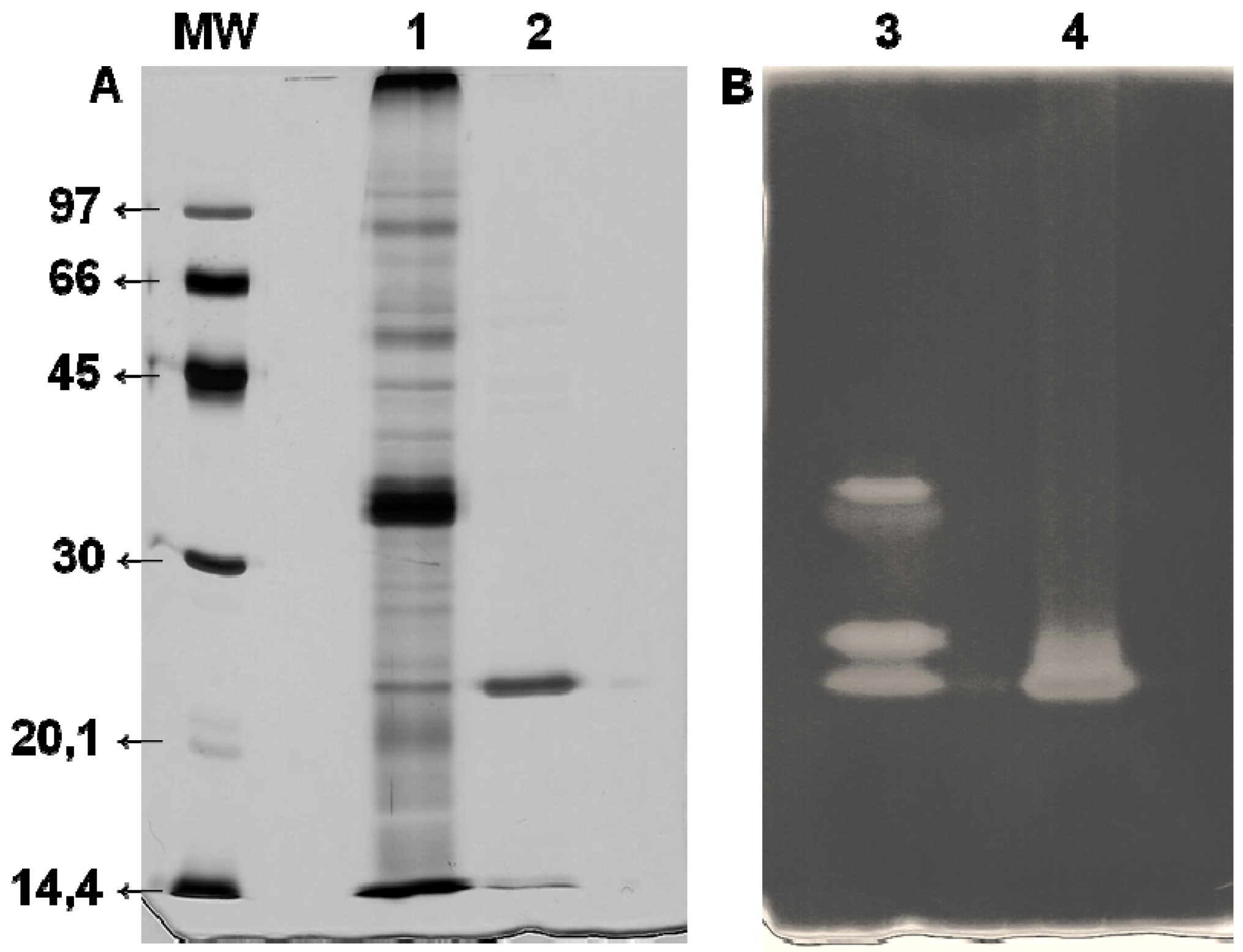
2.6. Enzyme Purification
2.7. Electrophoresis
2.8. Measurement of Substrate Specificity
2.9. Enzymatic Hydrolysis
3. Results and Discussion
3.1. Enzymatic Characterization of the FCS from Aspergillus oryzae Grown on PDCR

{kind=link}
{kind=link}
{kind=link}
| Enzymatic activity | IU·mL−1 |
|---|---|
| Xylanase | 0.488 ± 0.011 |
| Pectinase | 0.258 ± 0.048 |
| Mannanase | 0.098 ± 0.006 |
| CM-cellulase | 0.095 ± 0.004 |
| FPase | 0.081 ± 0.015 |
| Avicelase | 0.009 ± 0.001 |
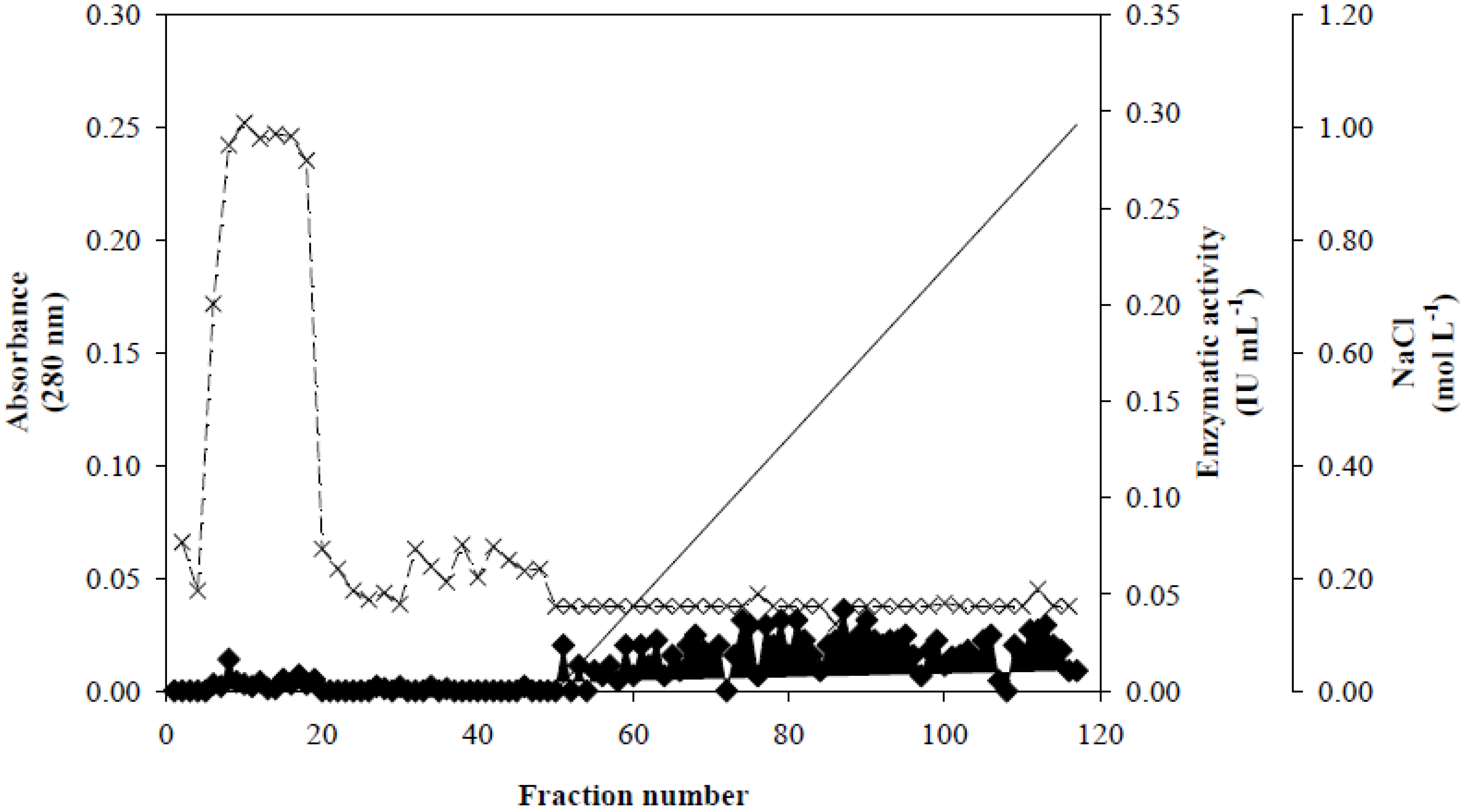
3.2. Xyl-O1 Purification
 ], xylanase activity is represented by thin Xs [
], xylanase activity is represented by thin Xs [  ], and the linear gradient (0–1.0 mol·L−1 NaCl in equilibration buffer) is represented by a solid line [
], and the linear gradient (0–1.0 mol·L−1 NaCl in equilibration buffer) is represented by a solid line [  ].
], xylanase activity is represented by thin Xs [ ], and the linear gradient (0–1.0 mol·L−1 NaCl in equilibration buffer) is represented by a solid line [ ].
].
], xylanase activity is represented by thin Xs [ ], and the linear gradient (0–1.0 mol·L−1 NaCl in equilibration buffer) is represented by a solid line [ ].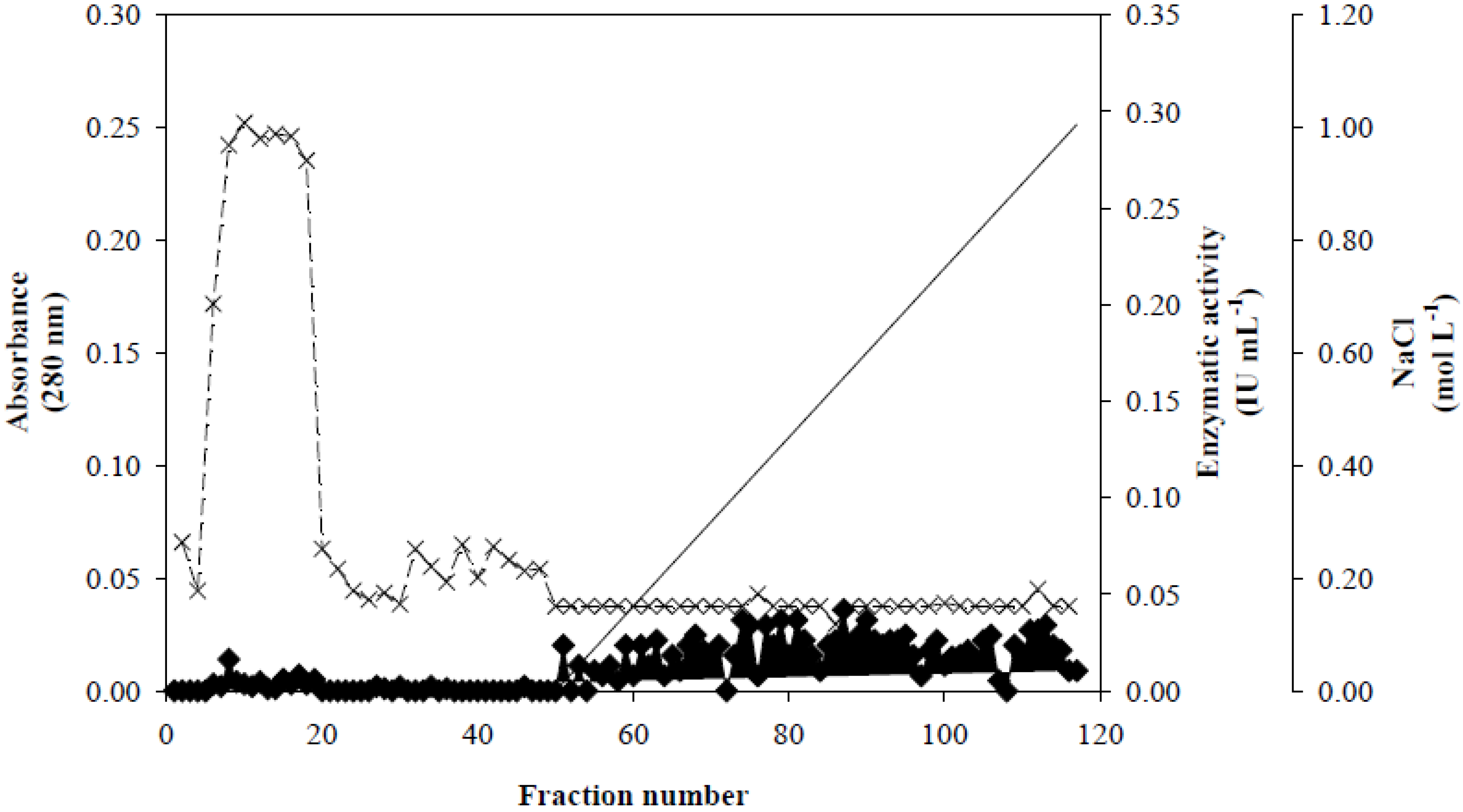
| Purification steps | Total volume (mL) | Total protein (mg) | Total activity (IU) | Specific activity (IU·mg−1) * | Purification (fold) | Yield (%) |
|---|---|---|---|---|---|---|
| Filtered culture supernatant | 110.00 | 5.830 | 53.350 | 9.151 | 1.000 | 100.00 |
| Ultrafiltrate | 100.00 | 1.500 | 29.400 | 19.600 | 2.142 | 55.11 |
| Lyophilization | 4.50 | 0.828 | 1.647 | 1.989 | 0.217 | 3.08 |
| Sephacryl S-100 (fractions 58–64) | 21.00 | 0.441 | 7.833 | 17.762 | 1.941 | 14.68 |
| Sephadex G-50 (fractions 71–84) | 40.00 | 0.080 | 9.880 | 123.500 | 13.496 | 18.52 |
| Q-Sepharose FF (fractions 7–18) | 36.00 | 0.036 | 8.928 | 248.000 | 27.101 | 16.74 |
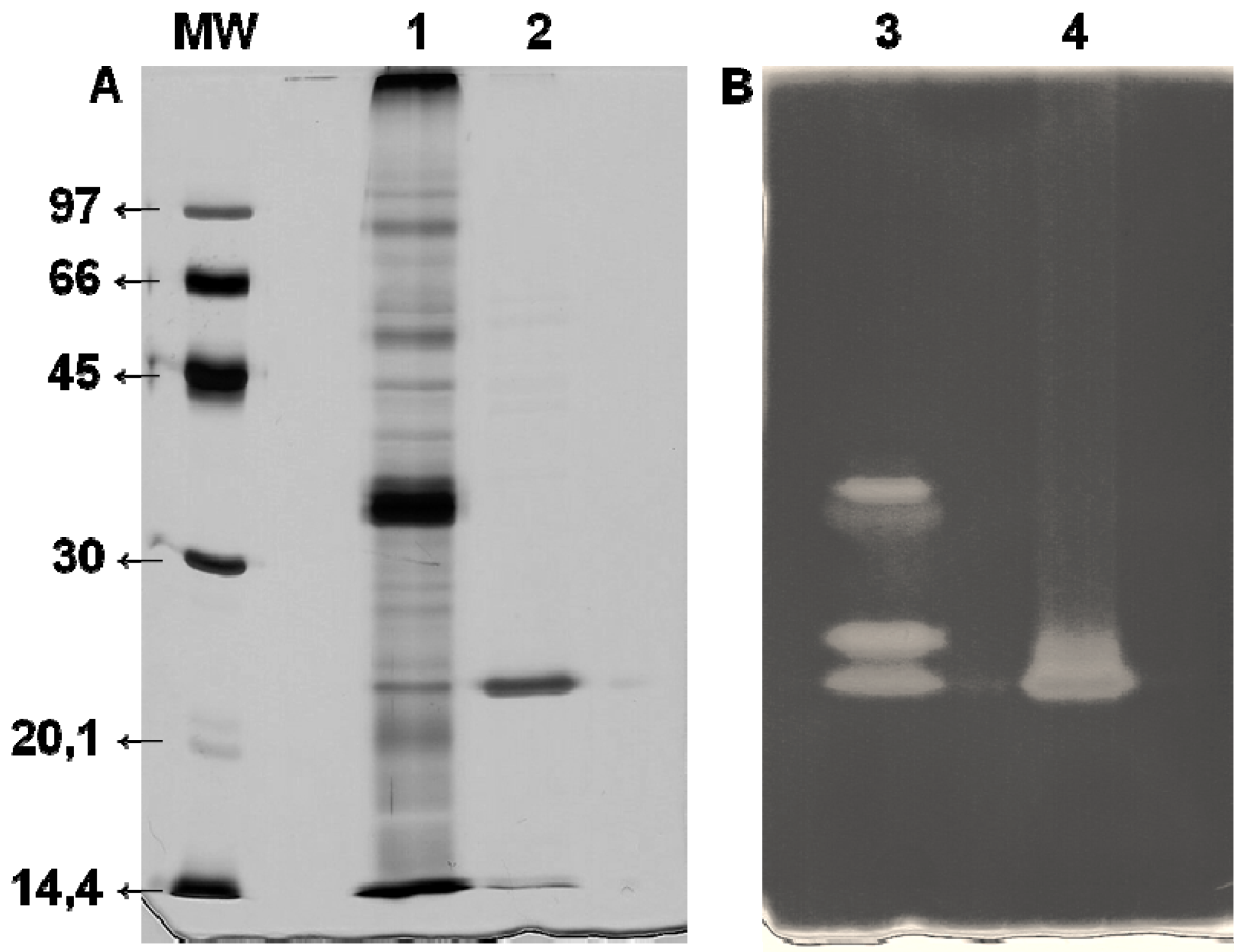
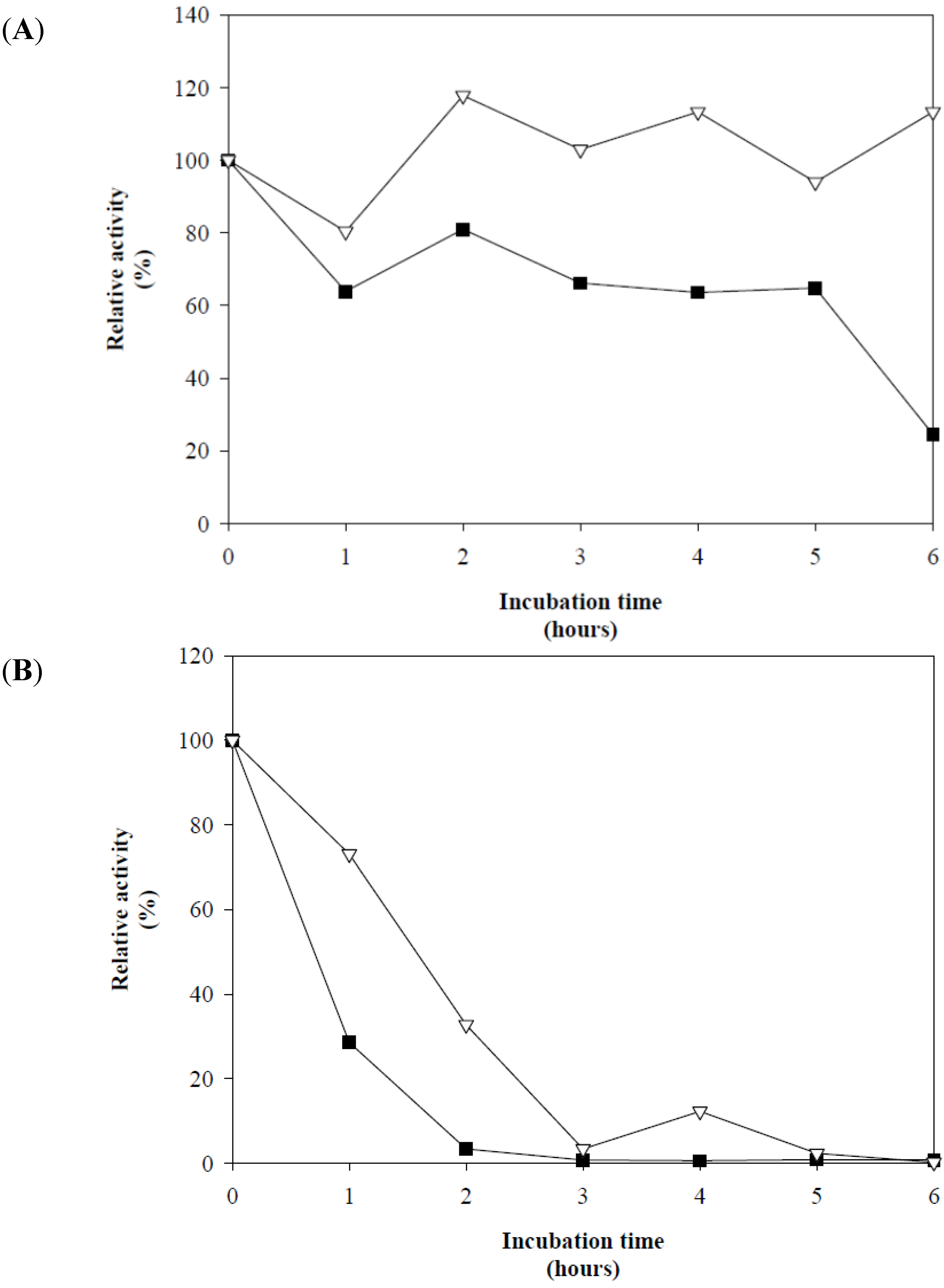
3.3. Enzyme Characterization
| Modifying agents or amino acids | Activity (IU·mL−1) | Relative activity (%) | Concentration (mmol·L−1) |
|---|---|---|---|
| H2O control | 0.262 ± 0.021 | 100.00 | --- |
| 4% Ethanol control | 0.285 ± 0.026 | 100.00 | --- |
| NBS | 0.285 ± 0.079 | 108.42 | 1 |
| DTNB * | 0.399 ± 0.025 | 140.02 | 2 |
| DTT | 0.300 ± 0.037 | 114.35 | 20 |
| L-Cysteine | 0.367 ± 0.076 | 140.04 | 20 |
| H2O control | 0.290 ± 0.029 | 100.00 | --- |
| 4% Ethanol control | 0.322 ± 0.006 | 100.00 | --- |
| DTP * | 0.285 ± 0.025 | 88.43 | 2 |
| L-tryptophan | 0.285 ± 0.012 | 98.13 | 5 |
| Iodoacetamide | 0.255 ± 0.046 | 87.71 | 5 |
| DEPC | 0.248 ± 0.037 | 85.32 | 5 |
| EDC | 0.291 ± 0.046 | 100.08 | 5 |
| β-Mercaptoethanol | 0.397 ± 0.026 | 136.68 | 5 |
| H2O control | 0.306 ± 0.031 | 100.00 | --- |
| SDS | 0.225 ± 0.025 | 73.56 | 20 |
 ]. The control without L-cysteine [
]. The control without L-cysteine [  ].
]. The control without L-cysteine [ ].
].
]. The control without L-cysteine [ ].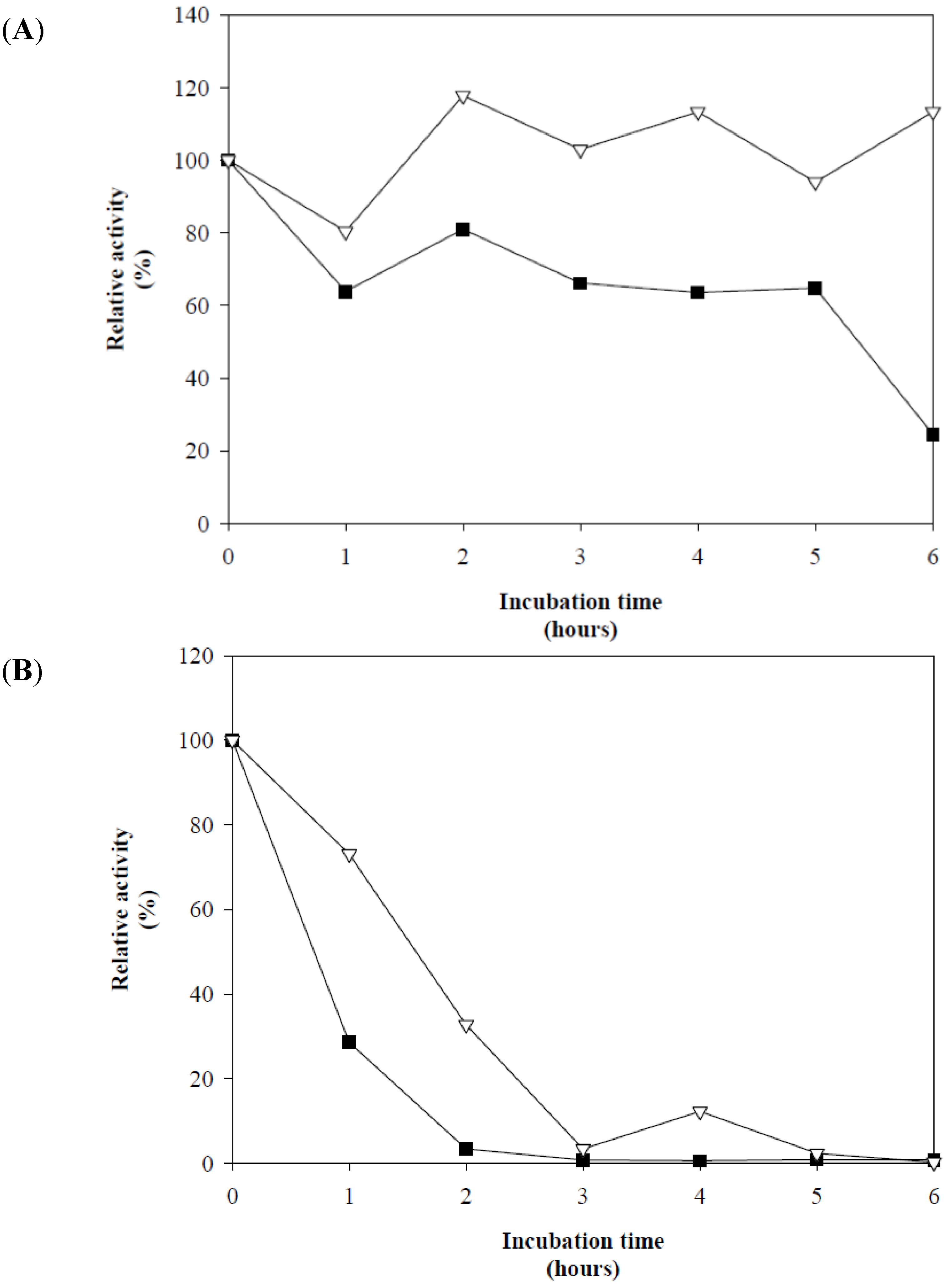
| Substrate | Main chain linkage | Activity (IU·mL−1) | Purified Xyl-O1 (% RA) a |
|---|---|---|---|
| Oat spelt xylan (S) b | β-1,4 | 0.659 ± 0.005 | 100.00 |
| Oat spelt xylan (I) c | β-1,4 | 0.003 ± 0.004 | 0.39 |
| Birchwood xylan (S) | β-1,4 | 0.560 ± 0.011 | 85.03 |
| Birchwood xylan (I) | β-1,4 | 0.228 ± 0.025 | 34.60 |
| Beechwood xylan (S) | β-1,4 | 0.556 ± 0.030 | 84.38 |
| Beechwood xylan (I) | β-1,4 | 0.024 ± 0.030 | 3.58 |
| 4- O-methyl-glucurono-D-xylan (I) | β-1,4 | 0.328 ± 0.028 | 49.87 |
| Filter paper | β-1,4 | 0.010 ± 0.007 | 1.46 |
| CM-cellulose | β-1,4 | 0.045 ± 0.004 | 6.82 |
| Avicel | β-1,4 | 0.000 ± 0.002 | 0.00 |
| Laminarin | β-1,3 | 0.039 ± 0.018 | 5.93 |
| Pectin | β-1,4 | 0.024 ± 0.019 | 3.62 |
| Mannan | β-1,4 | 0.002 ± 0.003 | 0.37 |
| PNPX | PNP-β-1,4 | 0.000 ± 0.000 | 0.00 |
| PNPA | PNP-α-1,4 | 0.000 ± 0.000 | 0.00 |
| PNPG | PNP-β-1,4 | 0.000 ± 0.000 | 0.00 |
| PNPM | PNP-β-1,4 | 0.000 ± 0.000 | 0.00 |
| Hydrolysis time (h) | Soluble | Insoluble | ||
|---|---|---|---|---|
| Specific reducing sugar (mg) * | Hydrolysis (%) | Specific reducing sugar (mg) * | Hydrolysis (%) | |
| 0 | 0.000 ± 0.000 | 0.00 | 0.031 ± 0.006 | 1.55 |
| 6 | 0.434 ± 0.011 | 21.72 | 0.209 ± 0.036 | 10.47 |
| 12 | 0.667 ± 0.029 | 33.35 | 0.351 ± 0.057 | 17.54 |
| 18 | 0.554 ± 0.025 | 27.69 | 0.345 ± 0.039 | 17.23 |
| 24 | 0.554 ± 0.025 | 27.70 | 0.350 ± 0.037 | 17.50 |
| Xylooligosaccharide products * | Hydrolysis (%) Soluble Fraction | Hydrolysis (%) Insoluble Fraction | ||||||
|---|---|---|---|---|---|---|---|---|
| Incubation time (h) | Incubation time (h) | |||||||
| 6 | 12 | 18 | 24 | 6 | 12 | 18 | 24 | |
| X1 | 0.005 | 0.000 | 0.000 | 0.000 | 0.015 | 0.000 | 0.000 | 0.000 |
| X2 | 0.178 | 0.280 | 0.000 | 0.000 | 0.245 | 0.000 | 0.000 | 0.000 |
| X3 | 0.078 | 0.000 | 0.008 | 0.000 | 0.083 | 0.047 | 0.000 | 0.000 |
| X4 | 0.022 | 0.000 | 0.045 | 0.016 | 0.102 | 0.063 | 0.027 | 0.000 |
| X5 | 0.031 | 0.000 | 0.005 | 0.005 | 0.066 | 0.055 | 0.005 | 0.000 |
| X6 | 0.033 | 0.000 | 0.005 | 0.000 | 0.052 | 0.022 | 0.010 | 0.000 |
4. Conclusions
Acknowledgments
References
- Howard, R.L.; Abotsi, E.; van Rensburg, E.L.J.; Howard, S. Lignocellulose biotechnology: Issues of bioconversion and enzyme production. Afr. J. Biotechnol. 2003, 2, 602–619. [Google Scholar]
- Andreaus, J.; Filho, E.X.F.; Bon, E.P.S. A Review on Biotechnology of Holocellulose-Degrading Enzymes. In Biocatalysis and Bioenerggy; Hou, C.T., Shaw, J.F., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; pp. 195–229. [Google Scholar]
- Siqueira, F.G.; Siqueira, E.G.; Jaramillo, P.M.D.; Silveira, M.H.L.; Andreaus, J.; Couto, F.A.; Batista, L.R.; Filho, E.X.F. The potential of agro-industrial residues for production of holocellulase from filamentous fungi. Int. Biodeterior. Biodegrad. 2010, 64, 20–26. [Google Scholar] [CrossRef]
- Csiszár, E.; Szakács, G.; Rusznák, I. Combining traditional cotton scouring with cellulase enzymatic treatment. Text. Res. J. 1998, 68, 163–167. [Google Scholar] [CrossRef]
- Csiszár, E.; Urbánszki, K.; Szakács, G. Biotreatment of desized cotton fabric by commercial cellulase and xylanase enzymes. J. Mol. Catal. B 2001, 11, 1065–1072. [Google Scholar] [CrossRef]
- Turner, P.; Mamo, G.; Karlsson, E. Potential and utilization of thermophiles and thermostable enzymes in biorefining. Microb. Cell. Fact. 2007, 6, 1–23. [Google Scholar] [CrossRef]
- Murthy, P.; Naidu, M. Production and application of xylanase from Penicillium sp. utilizing coffee by-products. Food Bioprocess Technol. 2010, 5, 657–664. [Google Scholar] [CrossRef]
- Polizeli, M.L.T.M.; Rizzatti, A.C.S.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef]
- Salles, B.C.; Cunha, R.B.; Fontes, W.; Sousa, M.V.; Ferreira-Filho, E.X. Purification and characterization of a new xylanase from Acrophialophora nainiana. J. Biotechnol. 2000, 81, 199–204. [Google Scholar]
- Vries, R.P.; Visser, J. Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiol. Mol. Biol. Rev. 2001, 65, 497–522. [Google Scholar] [CrossRef]
- Cardoso, O.A.V.; Filho, E.X.F. Purification and characterization of a novel cellulase-free xylanase from Acrophialophora nainiana. FEMS Microbiol. Lett. 2003, 223, 309–314. [Google Scholar] [CrossRef]
- Kobayashi, T.; Abe, K.; Asai, K.; Gomi, K.; Juvvadi, P.R.; Kato, M.; Kitamoto, K.; Takeuchi, M.; Machida, M. Genomics of Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2007, 71, 646–670. [Google Scholar] [CrossRef]
- Maeda, H.; Yamagata, Y.; Abe, K.; Hasegawa, F.; Machida, M.; Ishioka, R.; Gomi, K.; Nakajima, T. Purification and characterization of a biodegradable plastic-degrading enzyme from Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2005, 67, 778–788. [Google Scholar] [CrossRef]
- Filho, E.X.F.; Puls, J.; Coughlan, M.P. Biochemical characteristics of two endo-β-1,4-xylanases produced by Penicillium capsulatum. J. Ind. Microbiol. Biotechnol. 1993, 11, 171–180. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of cellulase activities. Int. Union Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Ximenes, F.A.; Silveira, F.Q.P.; Filho, E.X.F. Production of β-xylosidase activity by Trichoderma harzianum strains. Curr. Microbiol. 1996, 33, 71–77. [Google Scholar]
- Leatherbarrow, R.J. Enzfitter Manual, a Non-Linear Curve Fitting Program for Windows; Biosoft: London, UK, 1999. [Google Scholar]
- Segel, I.H. Effects of pH and Temperature. In Enzyme Kinetics—Behavior and Analysis of Rapid Equilibrium and Steady-State Enzyme Systems; John Wiley & Sons, Inc.: New York, NY, USA, 1993; pp. 885–942. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Blum, H.; Beier, H.; Gross, H.J. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Sunna, A.; Puls, J.; Antranikian, G. Characterization of the xylanolytic enzyme system of the extreme thermophilic anaerobic bacteria Thermotoga maritima, T. neapolitana, and T. thermarum. Comp. Biochem. Physiol. A 1997, 118, 453–461. [Google Scholar]
- Noronha, E.F.; Ulhoa, C.J. Characterization of a 29-kDa β-1,3-glucanase from Trichoderma harzianum. FEMS Microbiol. Lett. 2000, 183, 119–123. [Google Scholar]
- Ryan, S.E.; Nolan, K.; Thompson, R.; Gubitz, G.M.; Savage, A.V.; Tuohy, M.G. Purification and characterization of a new low molecular weight endoxylanase from Penicillium capsulatum. Enzyme Microb. Technol. 2003, 33, 775–785. [Google Scholar] [CrossRef]
- Adsul, M.G.; Ghule, J.E.; Singh, R.; Shaikh, H.; Bastawde, K.B.; Gokhale, D.V.; Varma, A.J. Polysaccharides from bagasse: Applications in cellulase and xylanase production. Carbohydr. Polym. 2004, 57, 67–72. [Google Scholar] [CrossRef]
- Gawande, P.V.; Kamat, M.Y. Production of Aspergillus xylanase by lignocellulosic waste fermentation and its application. J. Appl. Microbiol. 1999, 87, 511–519. [Google Scholar] [CrossRef]
- Chipeta, Z.; du Preez, J.; Szakacs, G.; Christopher, L. Xylanase production by fungal strains on spent sulphite liquor. Appl. Microbiol. Biotechnol. 2005, 69, 71–78. [Google Scholar] [CrossRef]
- Chipeta, Z.; du Preez, J.; Christopher, L. Effect of cultivation pH and agitation rate on growth and xylanase production by Aspergillus oryzae in spent sulphite liquor. J. Ind. Microbiol. Biotechnol. 2008, 35, 587–594. [Google Scholar] [CrossRef]
- Kronbauer, E.A.W.; Peralta, R.M.; Osaku, C.A.; Kadowaki, M.K. Xylanase production by Aspergillus casielus induced with different carbon sources. Bol. Cent. Pesqui. Process Aliment. 2007, 25, 207–216. [Google Scholar]
- Isil, S.; Nilufer, A. Investigation of factors affecting xylanase activity from Trichoderma harzianum 1073 D3. Braz. Arch. Biol. Technol. 2005, 48, 187–193. [Google Scholar]
- Wong, K.K.; Tan, L.U.; Saddler, J.N. Multiplicity of beta-1,4-xylanase in microorganisms: Functions and applications. Microbiol. Rev. 1988, 52, 305–317. [Google Scholar]
- Ximenes, F.A.; Sousa, M.V.; Puls, J.; da Silva, F.G.; Filho, E.X.F. Purification and characterization of a low-molecular-weight xylanase produced by Acrophialophora nainiana. Curr. Microbiol. 1999, 38, 18–21. [Google Scholar]
- Silva, C.H.C.; Puls, J.; Sousa, M.V.; Filho, E.X.F. Purification and characterization of a low molecular weight xylanase from solid-state cultures of Aspergillus fumigatus Fresenius. Rev. Microbiol. 1999, 30, 114–119. [Google Scholar]
- Poutanen, K. An α-L-arabinofuranosidase of Trichoderma reesei. J. Biotechnol. 1988, 7, 271–281. [Google Scholar]
- Fang, H.-Y.; Chang, S.-M.; Lan, C.-H.; Fang, T.J. Purification and characterization of a xylanase from Aspergillus carneus M34 and its potential use in photoprotectant preparation. Process Biochem. 2008, 43, 49–55. [Google Scholar]
- Panagiotou, G.; Olsson, L. Effect of compounds released during pretreatment of wheat straw on microbial growth and enzymatic hydrolysis rates. Biotechnol. Bioeng. 2007, 96, 250–258. [Google Scholar] [CrossRef]
- Kang, M.; Maeng, P.; Rhee, Y. Purification and characterization of two xylanases from alkalophilic Cephalosporium sp. strain RYM-202. Appl. Environ. Microbiol. 1996, 62, 3480–3482. [Google Scholar]
- Teixeira, R.S.S.; Siqueira, F.G.; Souza, M.V.; Filho, E.X.F.; Bon, E.P.S. Purification and characterization studies of a thermostable β-xylanase from Aspergillus awamori. J. Ind. Microbiol. Biotechnol. 2010, 37, 1041–1051. [Google Scholar] [CrossRef]
- Kimura, T.; Suzuki, H.; Furuhashi, H.; Aburatani, T.; Morimoto, K.; Karita, S.; Sakka, K.; Ohmiya, K. Molecular cloning, overexpression, and purification of a major xylanase from Aspergillus oryza. Biosci. Biotechnol. Biochem. 2000, 64, 2734–2738. [Google Scholar] [CrossRef]
- Sandrim, V.C.; Rizzatti, A.C.S.; Terenzi, H.F.; Jorge, J.A.; Milagres, A.M.F.; Polizeli, M.L.T.M. Purification and biochemical characterization of two xylanases produced by Aspergillus caespitosus and their potential for kraft pulp bleaching. Process Biochem. 2005, 40, 1823–1828. [Google Scholar] [CrossRef]
- Soren, D.; Jana, M.; Sengupta, S.; Ghosh, A. Purification and characterization of a low molecular weight endo-xylanase from mushroom Termitomyces clypeatus. Appl. Biochem. Biotechnol. 2010, 162, 373–389. [Google Scholar] [CrossRef]
- Carmona, E.C.; Fialho, M.B.; Buchgnani, É.B.; Coelho, G.D.; Brocheto-Braga, M.R.; Jorge, J.A. Production, purification and characterization of a minor form of xylanase from Aspergillus versicolor. Process Biochem. 2005, 40, 359–364. [Google Scholar] [CrossRef]
- Keskar, S.S.; Srinivasan, M.C.; Deshpande, V.V. Chemical modification of a xylanase from a thermotolerant Streptomyces. Evidence for essential tryptophan and cysteine residues at the active site. Biochem. J. 1989, 261, 49–40. [Google Scholar]
- Marui, M.; Nakanishi, K.; Yasui, T. Chemical modification of xylanases from Streptomyces sp. Biosci. Biotech. Biochem. 1993, 57, 662–663. [Google Scholar] [CrossRef]
- Christov, L.P.; Szakacs, G.; Balakrishnan, H. Production, partial characterization and use of fungal cellulase-free xylanases in pulp bleaching. Process Biochem. 1999, 34, 511–517. [Google Scholar] [CrossRef]
- Biely, P.; VrŠAnskÁ, M. Synthesis and hydrolysis of 1,3-β-xylosidic linkages by endo-1,4-β-xylanase of Cryptococcus albidus. Eur. J. Biochem. 1983, 129, 645–651. [Google Scholar]
- Kambourova, M.; Mandeva, R.; Fiume, I.; Maurelli, L.; Rossi, M.; Morana, A. Hydrolysis of xylan at high temperature by co-action of the xylanase from Anoxybacillus flavithermus BC and the β-xylosidase/α-arabinosidase from Sulfolobus solfataricus Oα. J. Appl. Microbiol. 2007, 102, 1586–1593. [Google Scholar]
- Eneyskaya, E.V.; Brumer, H.; Backinowsky, L.V.; Ivanen, D.R.; Kulminskaya, A.A.; Shabalin, K.A.; Neustroev, K.N. Enzymatic synthesis of β-xylanase substrates: transglycosylation reactions of the β-xylosidase from Aspergillus sp. Carbohydr. Res. 2003, 338, 313–325. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhu, Y.; Li, L.; Yu, X.; Kusakabe, I.; Kitaoka, M.; Hayashi, K. Transglycosylation reaction of xylanase B from the hyperthermophilic Thermotoga maritima with the ability of synthesis of tertiary alkyl β-D-xylobiosides and xylosides. J. Biotechnol. 2004, 114, 125–134. [Google Scholar]
- Fujita, M.; Shoda, S.-I. Xylanase-catalyzed synthesis of a novel polysaccharide having a glucose-xylose repeating unit, a cellulose-xylan hybrid polymer. J.Am.Chem.Soc. 1998, 120, 6411–6412. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Duarte, G.C.; Moreira, L.R.d.S.; Gómez-Mendoza, D.P.; Siqueira, F.G.d.; Batista, L.R.; Amaral, L.I.V.d.; Ricart, C.A.O.; Filho, E.X.F. Use of Residual Biomass from the Textile Industry as Carbon Source for Production of a Low-Molecular-Weight Xylanase from Aspergillus oryzae. Appl. Sci. 2012, 2, 754-772. https://doi.org/10.3390/app2040754
Duarte GC, Moreira LRdS, Gómez-Mendoza DP, Siqueira FGd, Batista LR, Amaral LIVd, Ricart CAO, Filho EXF. Use of Residual Biomass from the Textile Industry as Carbon Source for Production of a Low-Molecular-Weight Xylanase from Aspergillus oryzae. Applied Sciences. 2012; 2(4):754-772. https://doi.org/10.3390/app2040754
Chicago/Turabian StyleDuarte, Gilvan Caetano, Leonora Rios de Souza Moreira, Diana Paola Gómez-Mendoza, Félix Gonçalves de Siqueira, Luís Roberto Batista, Lourdes Isabel Velho do Amaral, Carlos André Ornelas Ricart, and Edivaldo Ximenes Ferreira Filho. 2012. "Use of Residual Biomass from the Textile Industry as Carbon Source for Production of a Low-Molecular-Weight Xylanase from Aspergillus oryzae" Applied Sciences 2, no. 4: 754-772. https://doi.org/10.3390/app2040754
APA StyleDuarte, G. C., Moreira, L. R. d. S., Gómez-Mendoza, D. P., Siqueira, F. G. d., Batista, L. R., Amaral, L. I. V. d., Ricart, C. A. O., & Filho, E. X. F. (2012). Use of Residual Biomass from the Textile Industry as Carbon Source for Production of a Low-Molecular-Weight Xylanase from Aspergillus oryzae. Applied Sciences, 2(4), 754-772. https://doi.org/10.3390/app2040754