
Trying on tRNA for Size: RNase P and the T-box Riboswitch as Molecular Rulers



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
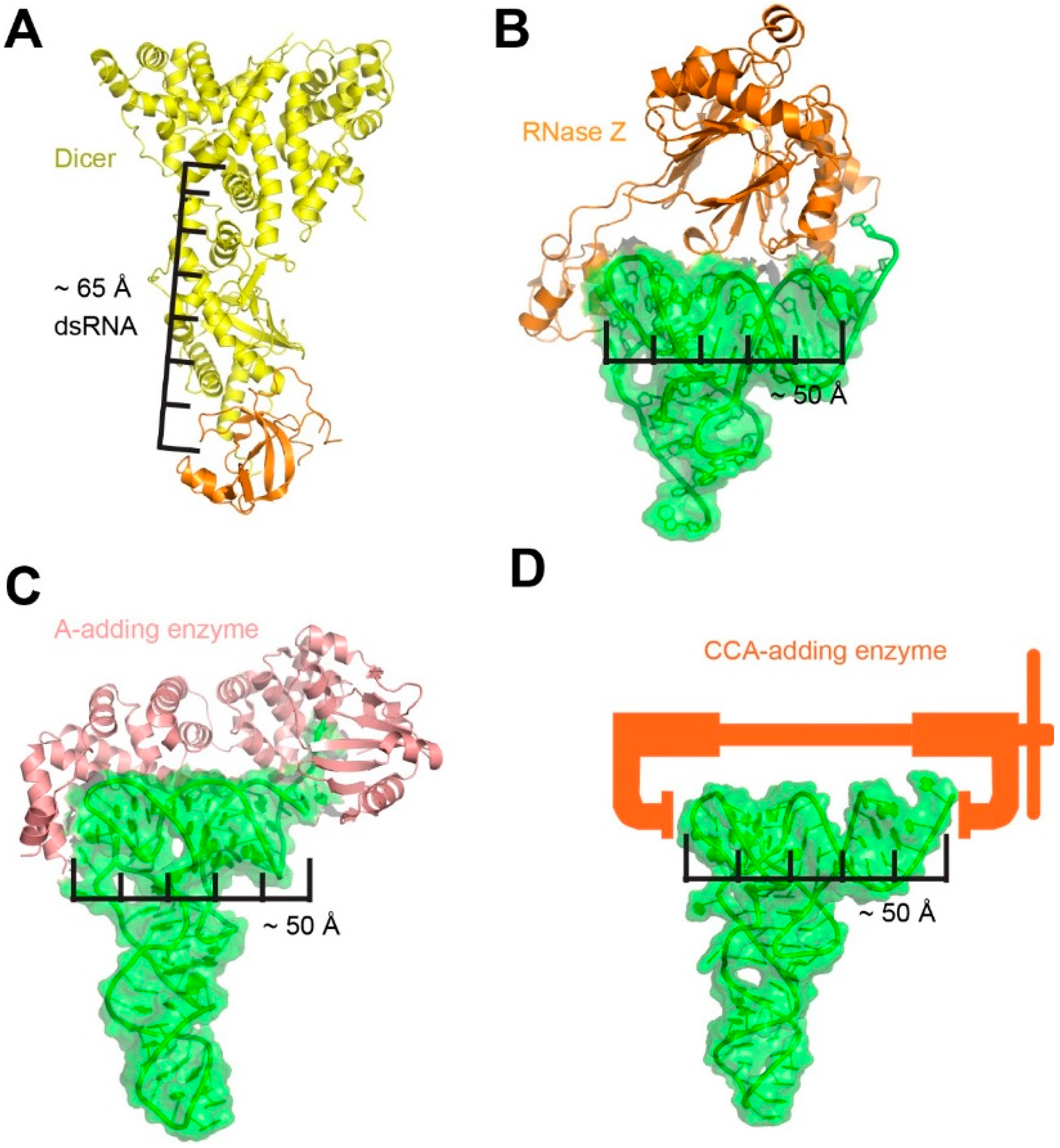
:1. Trying on tRNA for Size
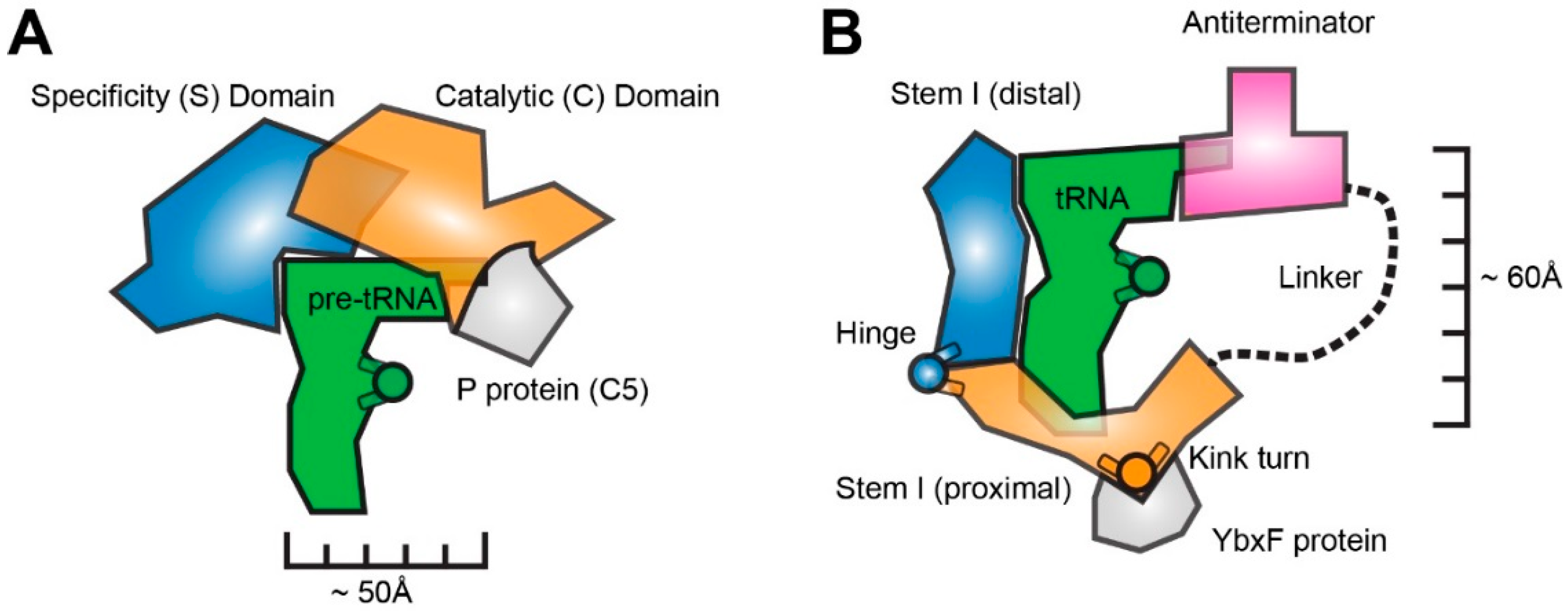
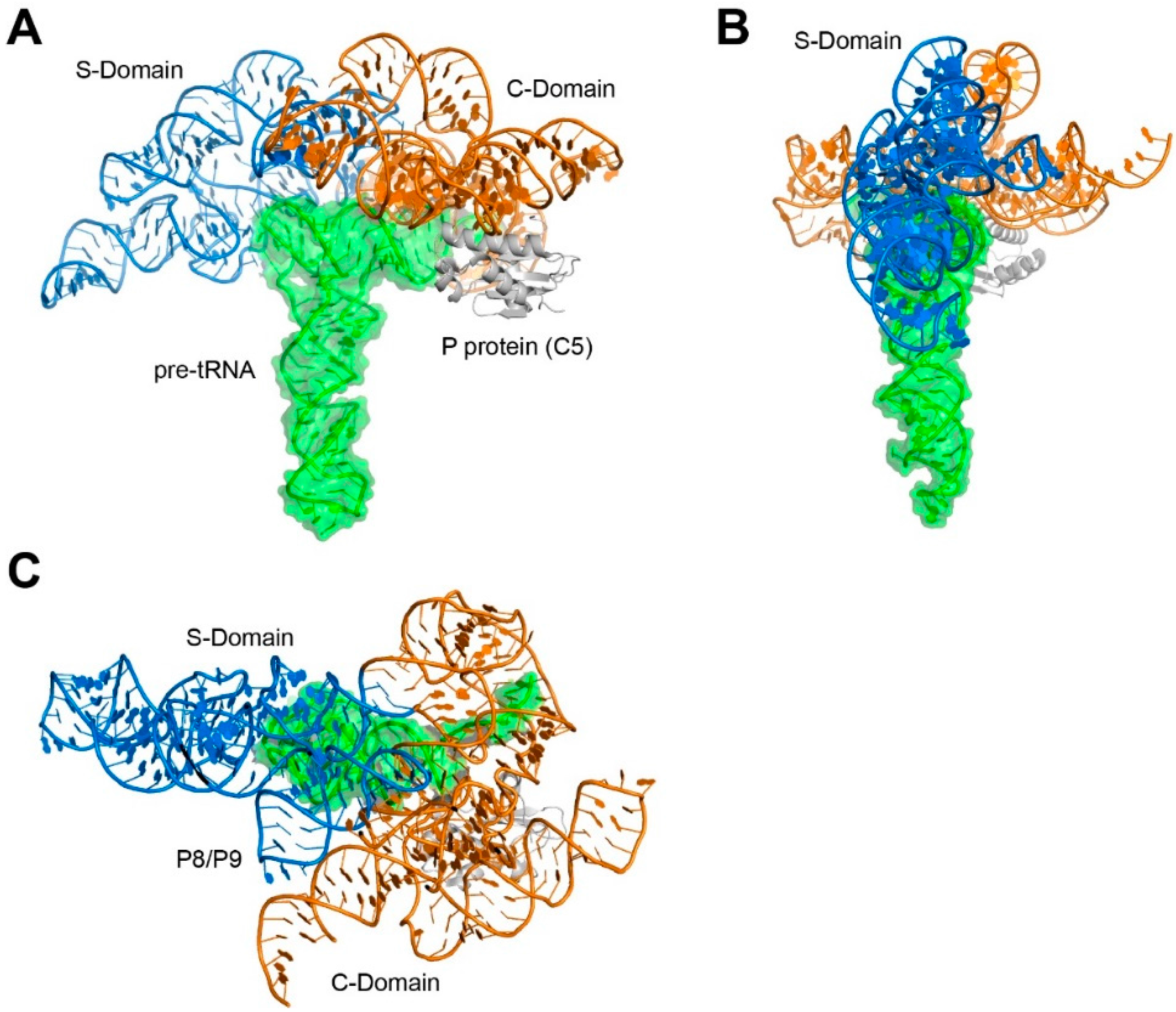
2. RNase P is a Rigid, Pre-Folded tRNA Ruler
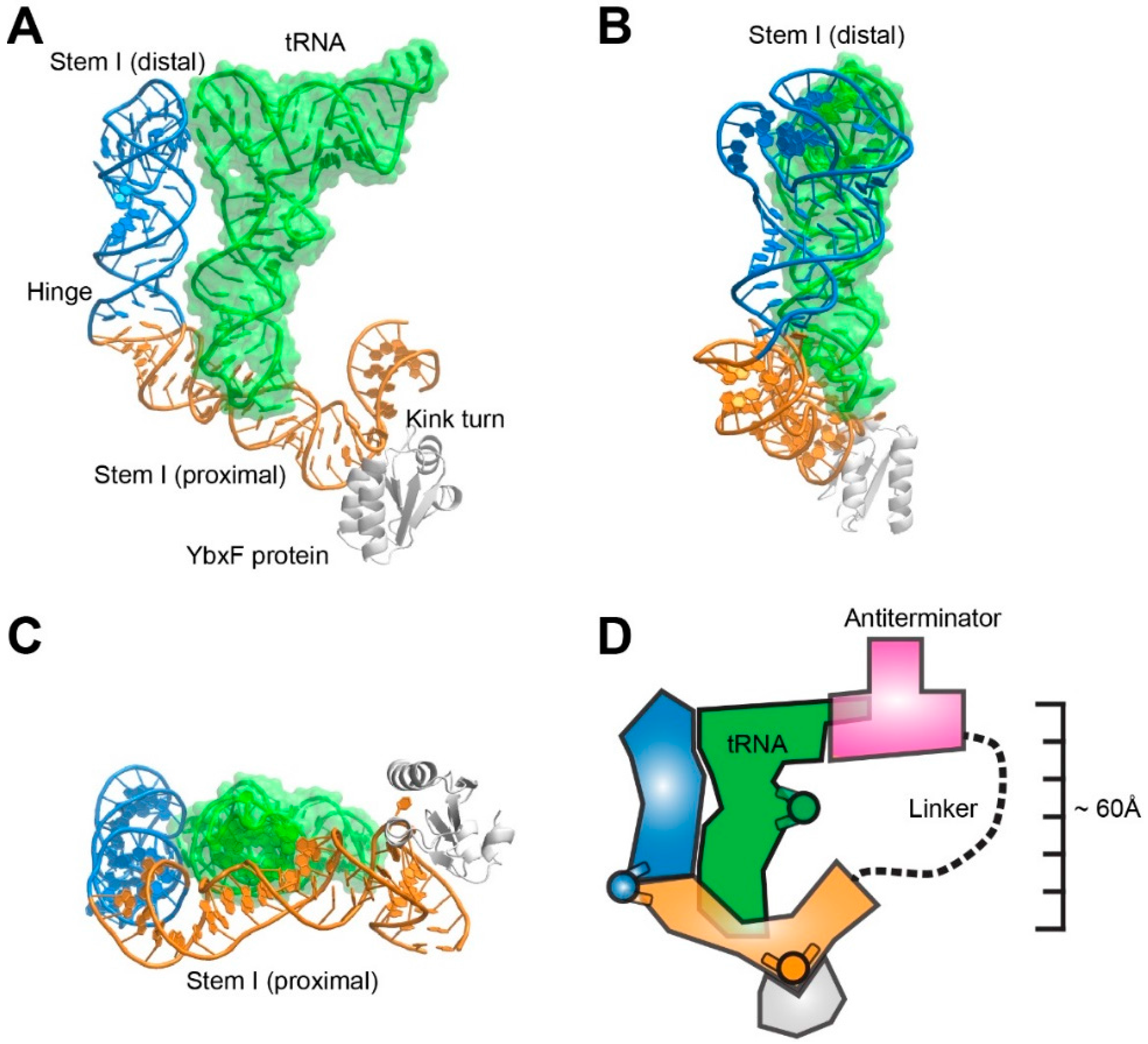
3. The T-box Riboswitch is a Flexible, Segmented tRNA Ruler
3.1. T-box Stem I is A Flexible, Hinged Ruler That Molds to tRNA.
3.2. The T-box Inter-Domain Linker and Antiterminator: A Molecular Lasso?
4. How to Make a Molecular Ruler with RNA
4.1. Shape, Phase, and Register
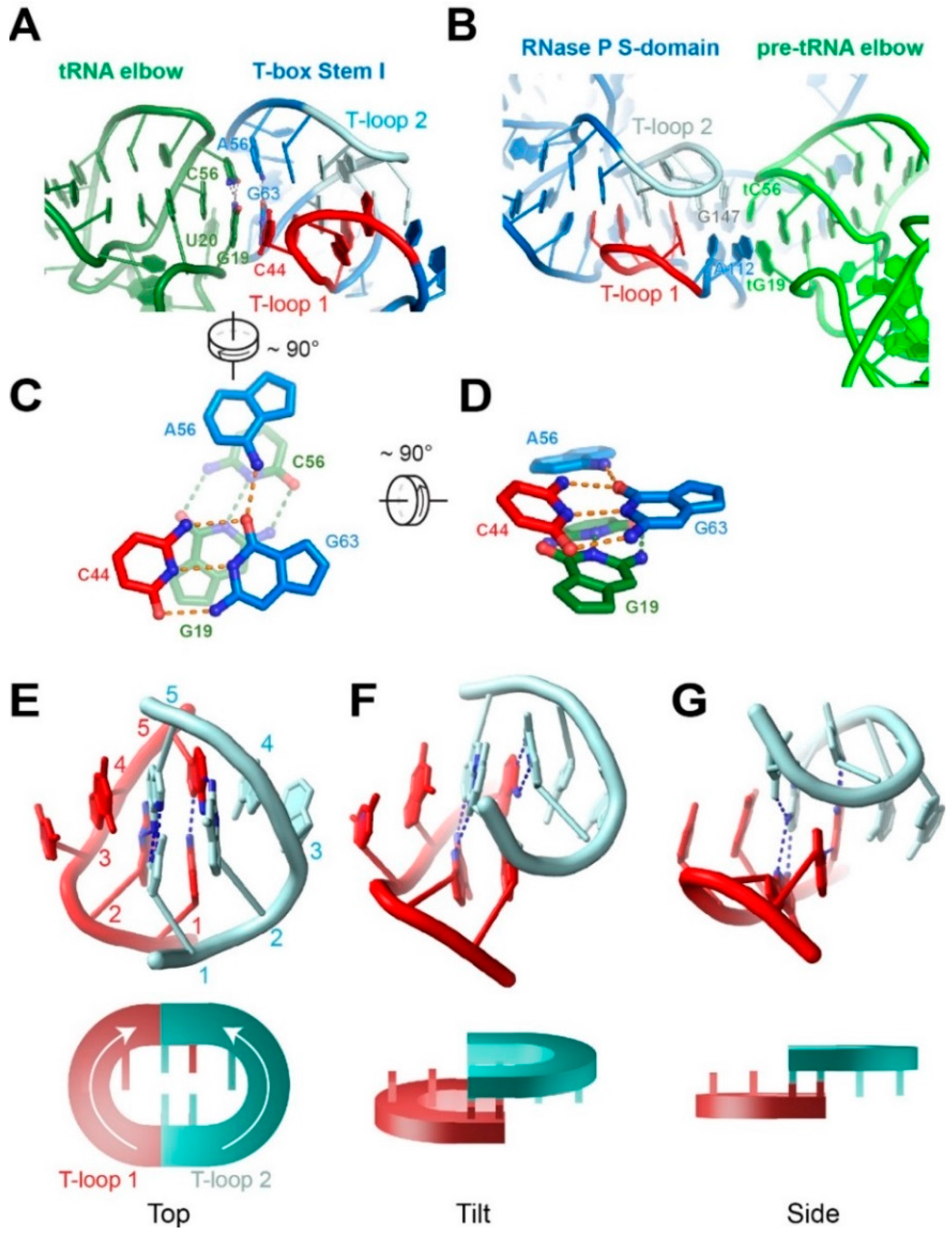
4.2. The Interdigitated T-Loop Motif
4.3. Non-tRNA Substrates of RNA Molecular Rulers
5. RNase P and T-box RNAs vs. Other Molecular Rulers
5.1. Diverse Definitions and Types of Molecular Rulers
5.2. RNase P and T-box are Trans-Acting Molecular Calipers
5.3. Protein Molecular Rulers that Measure RNA
5.4. Evolution and Application of RNA Molecular Rulers
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, S.H.; Suddath, F.L.; Quigley, G.J.; McPherson, A.; Sussman, J.L.; Wang, A.H.; Seeman, N.C.; Rich, A. Three-dimensional tertiary structure of yeast phenylalanine transfer RNA. Science 1974, 185, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Structure of yeast phenylalanine tRNA at 3 Å resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Olejniczak, M.; Dale, T.; Fahlman, R.P.; Uhlenbeck, O.C. Idiosyncratic tuning of trnas to achieve uniform ribosome binding. Nat. Struct. Mol. Biol. 2005, 12, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Giegé, R.; Jühling, F.; Pütz, J.; Stadler, P.; Sauter, C.; Florentz, C. Structure of transfer RNAs: Similarity and variability. Wiley Interdiscip. Rev. RNA 2011, 3, 37–61. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.W.; Nolan, J.M.; Haas, E.S.; Rubio, M.A.; Major, F.; Pace, N.R. Comparative analysis of ribonuclease P RNA using gene sequences from natural microbial populations reveals tertiary structural elements. Proc. Natl. Acad. Sci. USA 1996, 93, 3001–3006. [Google Scholar] [CrossRef] [PubMed]
- Kirsebom, L.A.; Svard, S.G. Base pairing between Escherichia coli RNase P RNA and its substrate. EMBO J. 1994, 13, 4870–4876. [Google Scholar] [PubMed]
- Kole, R.; Altman, S. Properties of purified ribonuclease P from Escherichia coli. Biochemistry 1981, 20, 1902–1906. [Google Scholar] [CrossRef] [PubMed]
- Engelke, D.R.; Fierke, C.A. The evolution of RNase P. RNA 2015, 21, 517–518. [Google Scholar] [CrossRef] [PubMed]
- James, B.D.; Olsen, G.J.; Liu, J.S.; Pace, N.R. The secondary structure of ribonuclease P RNA, the catalytic element of a ribonucleoprotein enzyme. Cell 1988, 52, 19–26. [Google Scholar] [CrossRef]
- Reich, C.; Olsen, G.J.; Pace, B.; Pace, N.R. Role of the protein moiety of ribonuclease p, a ribonucleoprotein enzyme. Science 1988, 239, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Svärd, S.G.; Kirsebom, L.A. Determinants of Escherichia coli RNase p cleavage site selection: A detailed in vitro and in vivo analysis. Nucleic Acids Res. 1993, 21, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kahle, D.; Wehmeyer, U.; Krupp, G. Substrate recognition by RNase P and by the catalytic M1 RNA: Identification of possible contact points in pre-tRNAs. EMBO J. 1990, 9, 1929–1937. [Google Scholar] [PubMed]
- Carrara, G.; Calandra, P.; Fruscoloni, P.; Doria, M.; Tocchini-Valentini, G.P. Site selection by xenopus laevis RNAase P. Cell 1989, 58, 37–45. [Google Scholar] [CrossRef]
- Bothwell, A.L.; Stark, B.C.; Altman, S. Ribonuclease P substrate specificity: Cleavage of a bacteriophage phi80-induced RNA. Proc. Natl. Acad. Sci. USA 1976, 73, 1912–1916. [Google Scholar] [CrossRef] [PubMed]
- Reiter, N.J.; Osterman, A.; Torres-Larios, A.; Swinger, K.K.; Pan, T.; Mondragón, A. Structure of a bacterial ribonuclease P holoenzyme in complex with tRNA. Nature 2010, 468, 784–789. [Google Scholar] [CrossRef] [PubMed]
- Torres-Larios, A.; Swinger, K.K.; Krasilnikov, A.S.; Pan, T.; Mondragón, A. Crystal structure of the RNA component of bacterial ribonuclease P. Nature 2005, 437, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Mondragon, A. Structural studies of RNase P. Annu. Rev. Biophys. 2013, 42, 537–557. [Google Scholar] [CrossRef] [PubMed]
- The pymol molecular graphics system. Available online: http://www.pymol.org/.
- Zhang, J.; Ferré-D'Amaré, A.R. Structure and mechanism of the T-box riboswitches. Wiley Interdiscip. Rev. RNA 2015, 6, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Grundy, F.J.; Winkler, W.C.; Henkin, T.M. tRNA-mediated transcription antitermination in vitro: Codon-anticodon pairing independent of the ribosome. Proc. Natl. Acad. Sci. USA 2002, 99, 11121–11126. [Google Scholar] [CrossRef] [PubMed]
- Grundy, F.J.; Henkin, T.M. tRNA as a positive regulator of transcription antitermination in b. Subtilis. Cell 1993, 74, 475–482. [Google Scholar] [CrossRef]
- Vitreschak, A.G.; Mironov, A.A.; Lyubetsky, V.A.; Gelfand, M.S. Comparative genomic analysis of T-box regulatory systems in bacteria. RNA 2008, 14, 717–735. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Preciado, A.; Henkin, T.M.; Grundy, F.J.; Yanofsky, C.; Merino, E. Biochemical features and functional implications of the RNA-based T-box regulatory mechanism. Microbiol. Mol. Biol. Rev. 2009, 73, 36–61. [Google Scholar] [CrossRef] [PubMed]
- Apostolidi, M.; Saad, N.Y.; Drainas, D.; Pournaras, S.; Becker, H.D.; Stathopoulos, C. A glys T-box riboswitch with species-specific structural features responding to both proteinogenic and nonproteinogenic trnagly isoacceptors. RNA 2015, 21, 1790–1806. [Google Scholar] [CrossRef] [PubMed]
- Saad, N.Y.; Stamatopoulou, V.; Braye, M.; Drainas, D.; Stathopoulos, C.; Becker, H.D. Two-codon T-box riboswitch binding two trnas. Proc. Natl. Acad. Sci. USA 2013, 110, 12756–12761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ferré-D'Amaré, A.R. Co-crystal structure of a T-box riboswitch stem I domain in complex with its cognate tRNA. Nature 2013, 500, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ferré-D'Amaré, A.R. Direct evaluation of tRNA aminoacylation status by the T-box riboswitch using tRNA-mRNA stacking and steric readout. Mol. Cell 2014, 55, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Grigg, J.C.; Ke, A. Structural determinants for geometry and information decoding of trna by T box leader RNA. Structure 2013, 21, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.; Zavialov, A.; Li, W.; Stagg, S.M.; Sengupta, J.; Nielsen, R.C.; Nissen, P.; Harvey, S.C.; Ehrenberg, M.; Frank, J. Incorporation of aminoacyl-tRNA into the ribosome as seen by cryo-electron microscopy. Nat. Struct. Biol. 2003, 10, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Korostelev, A.; Trakhanov, S.; Laurberg, M.; Noller, H. Crystal structure of a 70s ribosome-tRNA complex reveals functional interactions and rearrangements. Cell 2006, 126, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Dunkle, J.A.; Wang, L.; Feldman, M.B.; Pulk, A.; Chen, V.B.; Kapral, G.J.; Noeske, J.; Richardson, J.S.; Blanchard, S.C.; Cate, J.H. Structures of the bacterial ribosome in classical and hybrid states of trna binding. Science 2011, 332, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.D. RNA versatility governs tRNA function: Why tRNA flexibility is essential beyond the translation cycle. Bioessays 2016. [Google Scholar] [CrossRef] [PubMed]
- Grigg, J.C.; Chen, Y.; Grundy, F.J.; Henkin, T.M.; Pollack, L.; Ke, A. T box RNA decodes both the information content and geometry of tRNA to affect gene expression. Proc. Natl. Acad. Sci. USA 2013, 110, 7240–7245. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.J.; Schmeing, T.M.; Moore, P.B.; Steitz, T.A. The kink-turn: A new RNA secondary structure motif. EMBO J. 2001, 20, 4214–4221. [Google Scholar] [CrossRef] [PubMed]
- Winkler, W.C.; Grundy, F.J.; Murphy, B.A.; Henkin, T.M. The GA motif: An RNA element common to bacterial antitermination systems, rRNA, and eukaryotic RNAs. RNA 2001, 7, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lilley, D.M. The kink turn, a key architectural element in RNA structure. J. Mol. Biol. 2016, 428, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Lilley, D.M. The K-turn motif in riboswitches and other RNA species. Biochim. Biophys. Acta 2014, 1839, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.R.; Grundy, F.J.; Henkin, T.M. tRNA requirements for glyqs antitermination: A new twist on tRNA. RNA 2003, 9, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, A.V.; Grundy, F.J.; Henkin, T.M. T box riboswitches in actinobacteria: Translational regulation via novel tRNA interactions. Proc. Natl. Acad. Sci. USA 2015, 112, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Landick, R. A two-way street: Regulatory interplay between RNA polymerase and nascent RNA structure. Trends Biochem. Sci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Grundy, F.J.; Yousef, M.R.; Henkin, T.M. Monitoring uncharged tRNA during transcription of the Bacillus subtilis glyqs gene. J. Mol. Biol. 2005, 346, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lau, M.W.; Ferré-D'Amaré, A.R. Ribozymes and riboswitches: Modulation of RNA function by small molecules. Biochemistry 2010, 49, 9123–9131. [Google Scholar] [CrossRef] [PubMed]
- Abels, J.A.; Moreno-Herrero, F.; van der Heijden, T.; Dekker, C.; Dekker, N.H. Single-molecule measurements of the persistence length of double-stranded RNA. Biophys. J. 2005, 88, 2737–2744. [Google Scholar] [CrossRef] [PubMed]
- Kebbekus, P.; Draper, D.E.; Hagerman, P. Persistence length of RNA. Biochemistry 1995, 34, 4354–4357. [Google Scholar] [CrossRef] [PubMed]
- Richert, C.; Roughton, A.L.; Benner, S.A. Nonionic analogs of RNA with dimethylene sulfone bridges. J. Am. Chem. Soc. 1996, 118, 4518–4531. [Google Scholar] [CrossRef]
- Lescoute, A.; Leontis, N.B.; Massire, C.; Westhof, E. Recurrent structural RNA motifs, isostericity matrices and sequence alignments. Nucleic Acids Res. 2005, 33, 2395–2409. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, L.; Westhof, E.; Leontis, N.B. Tectorna: Modular assembly units for the construction of RNA nano-objects. Nucleic Acids Res. 2001, 29, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Hamma, T.; Ferré-D'Amaré, A.R. Pseudouridine synthases. Chem. Biol. 2006, 13, 1125–1135. [Google Scholar] [CrossRef] [PubMed]
- Hoang, C.; Ferré-D'Amaré, A.R. Cocrystal structure of a tRNA Psi55 pseudouridine synthase: Nucleotide flipping by an rna-modifying enzyme. Cell 2001, 107, 929–939. [Google Scholar] [CrossRef]
- Dalluge, J.J.; Hashizume, T.; Sopchik, A.E.; McCloskey, J.A.; Davis, D.R. Conformational flexibility in RNA: The role of dihydrouridine. Nucleic Acids Res. 1996, 24, 1073–1079. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.W.; Chetnani, B.; Mondragón, A. Structure and function of the T-loop structural motif in noncoding RNAs. Wiley Interdiscip. Rev. RNA 2013, 4, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Jossinet, F.; Gautheret, D. A universal RNA structural motif docking the elbow of trna in the ribosome, rnase P and T-box leaders. Nucleic Acids Res. 2013, 41, 5494–5502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ferré-D'Amaré, A.R. The trna elbow in structure, recognition and evolution. Life 2016. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, T.; Watanabe, Y. T-armless tRNAs and elongated elongation factor Tu. IUBMB Life 2007, 59, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Wende, S.; Platzer, E.G.; Juhling, F.; Putz, J.; Florentz, C.; Stadler, P.F.; Morl, M. Biological evidence for the world’s smallest tRNAs. Biochimie 2014, 100, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Giege, T.; Giege, R.; Giege, P. Trna biology in mitochondria. Int. J. Mol. Sci. 2015, 16, 4518–4559. [Google Scholar] [CrossRef] [PubMed]
- Komine, Y.; Kitabatake, M.; Yokogawa, T.; Nishikawa, K.; Inokuchi, H. A tRNA-like structure is present in 10sa RNA, a small stable RNA from Escherichia coli. Proc. Natl. Acad. Sci. USA 1994, 91, 9223–9227. [Google Scholar] [CrossRef] [PubMed]
- Bothwell, A.L.; Garber, R.L.; Altman, S. Nucleotide sequence and in vitro processing of a precursor molecule to Escherichia coli 4.5 s RNA. J. Biol. Chem. 1976, 251, 7709–7716. [Google Scholar] [PubMed]
- Lyons, A.J.; Robertson, H.D. Detection of tRNA-like structure through RNase P cleavage of viral internal ribosome entry site RNAs near the AUG start triplet. J. Biol. Chem. 2003, 278, 26844–26850. [Google Scholar] [CrossRef] [PubMed]
- Wilusz, J.E.; Freier, S.M.; Spector, D.L. 3' end processing of a long nuclear-retained noncoding RNA yields a tRNA-like cytoplasmic RNA. Cell 2008, 135, 919–932. [Google Scholar] [CrossRef] [PubMed]
- Colussi, T.M.; Costantino, D.A.; Hammond, J.A.; Ruehle, G.M.; Nix, J.C.; Kieft, J.S. The structural basis of transfer RNA mimicry and conformational plasticity by a viral RNA. Nature 2014, 511, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Hammond, J.A.; Rambo, R.P.; Filbin, M.E.; Kieft, J.S. Comparison and functional implications of the 3D architectures of viral tRNA-like structures. RNA 2009, 15, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, H. Pocketknife tRNA hypothesis: Anticodons in mammal mitochondrial tRNA side-arm loops translate proteins? Biosystems 2013, 113, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Seligmann, H.; Raoult, D. Unifying view of stem-loop hairpin rna as origin of current and ancient parasitic and non-parasitic RNAs, including in giant viruses. Curr. Opin. Microbiol. 2015, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Derksen, M.; Mertens, V.; Pruijn, G.J. RNase P-mediated sequence-specific cleavage of RNA by engineered external guide sequences. Biomolecules 2015, 5, 3029–3050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, W.F. How cells measure length on subcellular scales. Trends Cell Biol. 2015, 25, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Wee, D.H.; Hughes, K.T. Molecular ruler determines needle length for the Salmonella Spi-1 injectisome. Proc. Natl. Acad. Sci. USA 2015, 112, 4098–4103. [Google Scholar] [CrossRef] [PubMed]
- Journet, L.; Agrain, C.; Broz, P.; Cornelis, G.R. The needle length of bacterial injectisomes is determined by a molecular ruler. Science 2003, 302, 1757–1760. [Google Scholar] [CrossRef] [PubMed]
- Katsura, I. Determination of bacteriophage lambda tail length by a protein ruler. Nature 1987, 327, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Katsura, I.; Hendrix, R.W. Length determination in bacteriophage lambda tails. Cell 1984, 39, 691–698. [Google Scholar] [CrossRef]
- Butler, P.J.G. Self-assembly of tobacco mosaic virus: The role of an intermediate aggregate in generating both specificity and speed. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1999, 354, 537–550. [Google Scholar] [CrossRef] [PubMed]
- MacRae, I.J.; Zhou, K.; Li, F.; Repic, A.; Brooks, A.N.; Cande, W.Z.; Adams, P.D.; Doudna, J.A. Structural basis for double-stranded RNA processing by dicer. Science 2006, 311, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Li de la Sierra-Gallay, I.; Mathy, N.; Pellegrini, O.; Condon, C. Structure of the ubiquitous 3′ processing enzyme RNase Z bound to transfer RNA. Nat. Struct. Mol. Biol. 2006, 13, 376–377. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, O.; Li de la Sierra-Gallay, I.; Piton, J.; Gilet, L.; Condon, C. Activation of tRNA maturation by downstream uracil residues in B. subtilis. Structure 2012, 20, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Martinez, A.; Tomita, K. Measurement of acceptor-tpsic helix length of tRNA for terminal a76-addition by a-adding enzyme. Structure 2015, 23, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, C.D.; Wilusz, J.E.; Zheng, Y.; Beal, P.A.; Joshua-Tor, L. On-enzyme refolding permits small RNA and tRNA surveillance by the CCA-adding enzyme. Cell 2015, 160, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Steitz, T.A. Mechanism of transfer RNA maturation by CCA-adding enzyme without using an oligonucleotide template. Nature 2004, 430, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Esakova, O.; Krasilnikov, A.S. Of proteins and RNA: The RNase P/MRP family. RNA 2010, 16, 1725–1747. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Ferré-DAmaré, A.R. Trying on tRNA for Size: RNase P and the T-box Riboswitch as Molecular Rulers. Biomolecules 2016, 6, 18. https://doi.org/10.3390/biom6020018
Zhang J, Ferré-DAmaré AR. Trying on tRNA for Size: RNase P and the T-box Riboswitch as Molecular Rulers. Biomolecules. 2016; 6(2):18. https://doi.org/10.3390/biom6020018
Chicago/Turabian StyleZhang, Jinwei, and Adrian R. Ferré-DAmaré. 2016. "Trying on tRNA for Size: RNase P and the T-box Riboswitch as Molecular Rulers" Biomolecules 6, no. 2: 18. https://doi.org/10.3390/biom6020018