Proteomic Analysis of Aphid-Resistant and -Sensitive Rose (Rosa Hybrida) Cultivars at Two Developmental Stages




Abstract
1. Introduction
2. Material and Methods
2.1. Sample Collection and Transportation
2.2. Protein Extraction and First Dimensional Gel Electrophoresis
2.3. Protein in Gel Digestion
2.4. Protein Identification Using MALDI-TOF MS and MS/MS Analysis
2.5. Immunoblot Analysis
2.6. First Dimensional Blue Native Page (BN-PAGE)
2.7. Sugar and Carbohydrate Content
2.8. Statistical Analysis
3. Results and Discussion
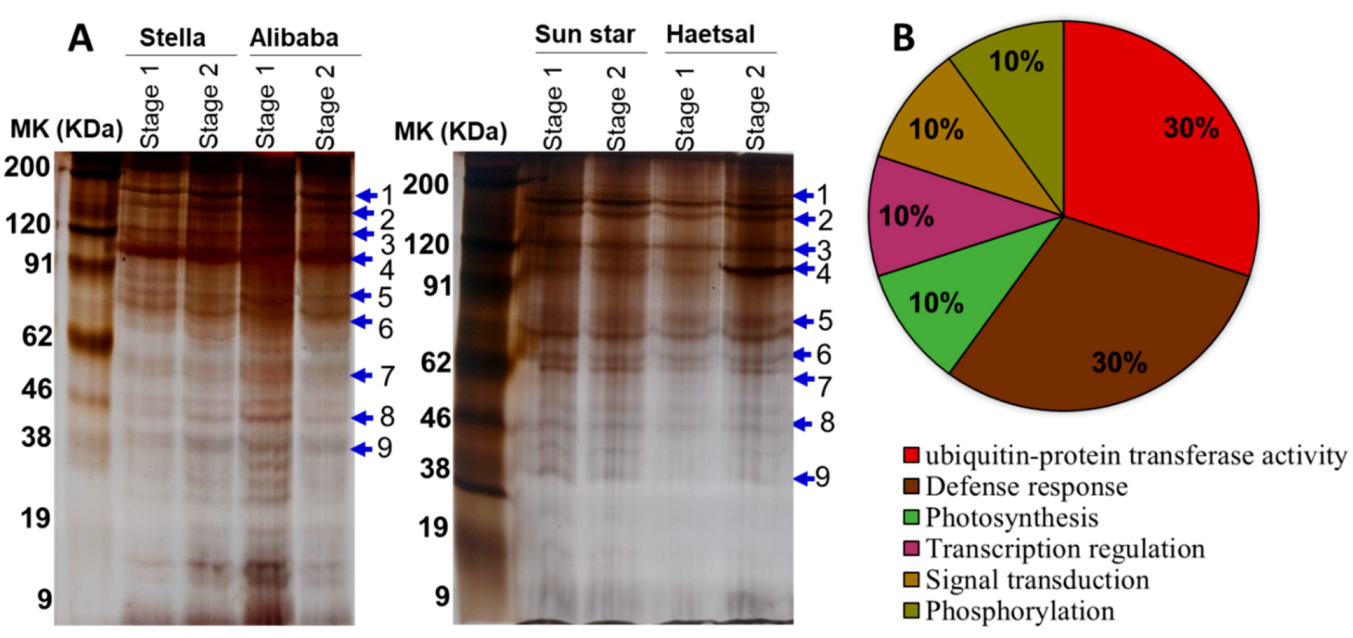
3.1. Relative Proteome Changes and Protein Identification
3.2. Proteins Related to Ubiquitin
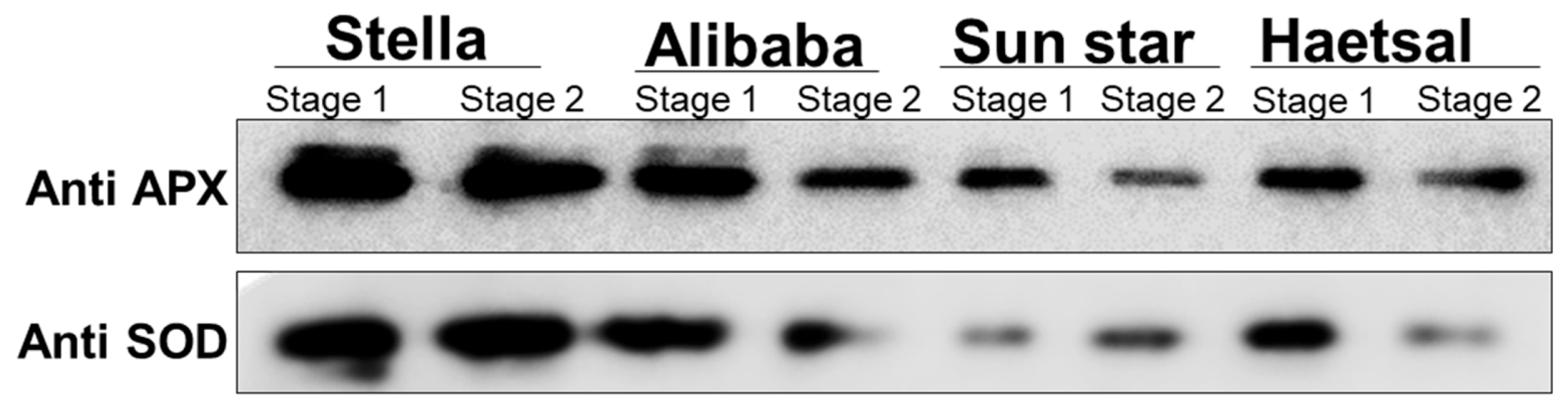
3.3. Proteins Related to the Defense Response
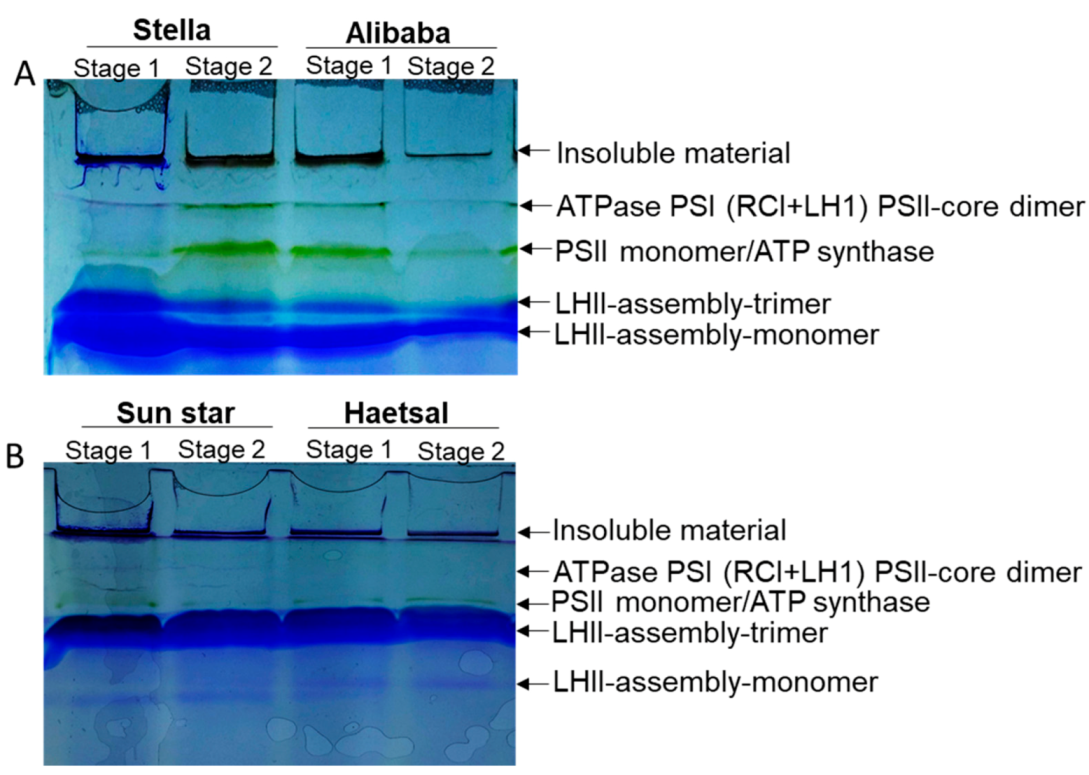
3.4. Proteins Related to Photosynthesis
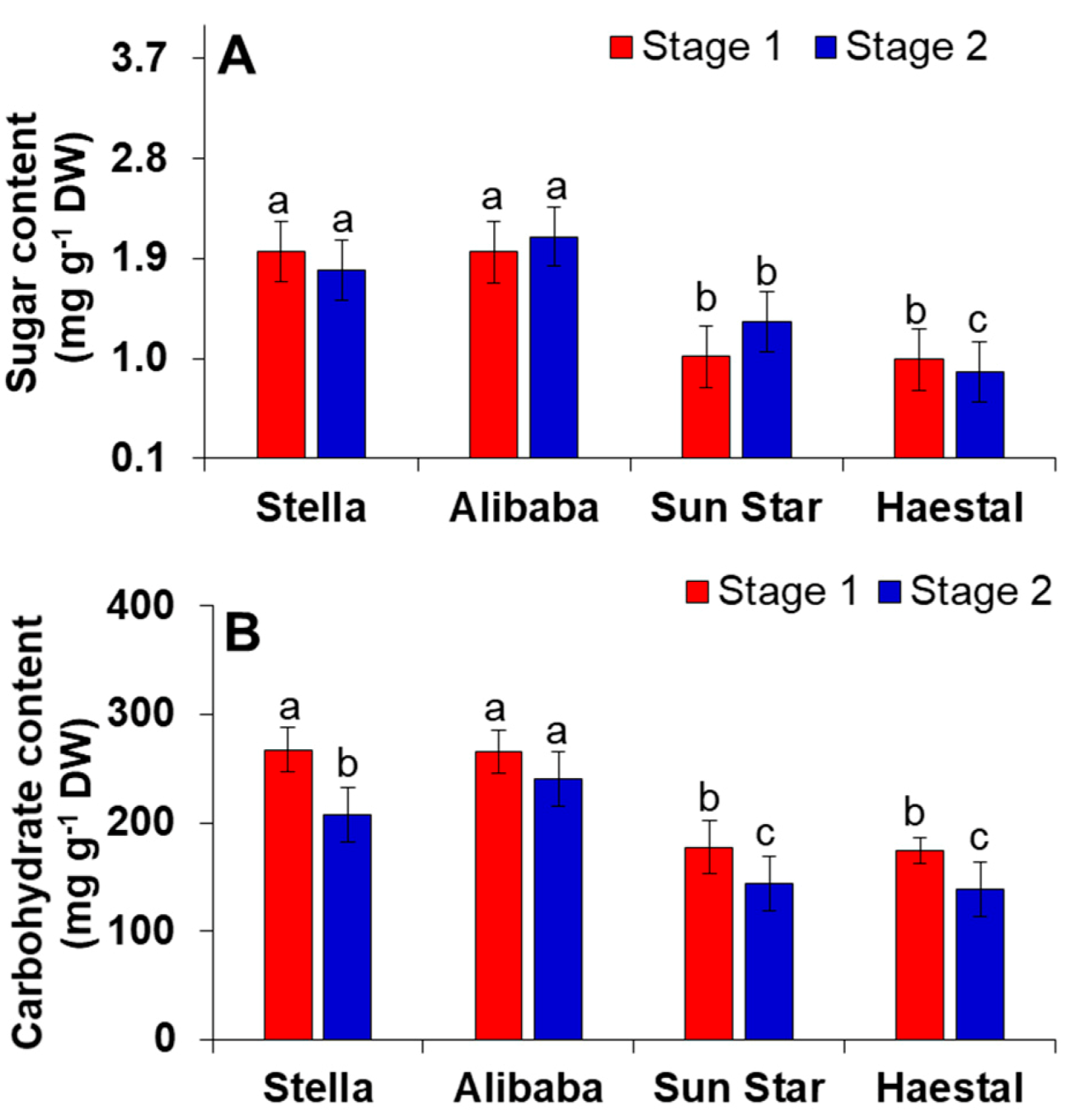
3.5. Metabolic Changes
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Gudin, S. Breeding. In Encyclopedia of Rose Science; Roberts, A.V., Debener, T., Gudin, S., Eds.; Academic Press: Oxford, UK, 2003; pp. 25–30. [Google Scholar]
- Debener, T.; Mattiesch, L. Construction of a genetic linkage map for roses using RAPD and AFLP markers. Theor. Appl. Genet. 1999, 99, 891–899. [Google Scholar] [CrossRef]
- Crespel, L.; Chirollet, M.; Durel, C.E.; Zhang, D.; Meynet, J.; Gudin, S. Mapping of qualitative and quantitative phenotypic traits in Rosa using AFLP markers. Theor. Appl. Genet. 2002, 105, 1207–1214. [Google Scholar] [PubMed]
- Dugo, M.L.; Satovic, Z.; Millan, T.; Cubero, J.I.; Rubiales, D.; Cabrera, A.; Torres, A.M. Genetic mapping of QTLs controlling horticultural traits in diploid roses. Theor. Appl. Genet. 2005, 111, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Denneboom, C.; Hattendorf, A.; Dolstra, O.; Debener, T.; Stam, P.; Visser, P.B. Construction of an integrated map of rose with AFLP, SSR, PK, RGA, RFLP, SCAR and morphological markers. Theor. Appl. Genet. 2005, 110, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Linde, M.; Hattendorf, A.; Kaufmann, H.; Debener, T. Powdery mildew resistance in roses: QTL mapping in different environments using selective genotyping. Theor. Appl. Genet. 2006, 113, 1081–1092. [Google Scholar] [CrossRef] [PubMed]
- Spiller, M.; Linde, M.; Oyant, H.S.; Tsai, C.J.; Byrne, D.H.; Smulders, M.J.; Foucher, F.; Debener, T. Towards a unified genetic map for diploid roses. Theor. Appl. Genet. 2011, 122, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, H.H.; Leus, L.; De Riek, J.; Van Huylenbroeck, J.; Van Bockstaele, E. Construction of a genetic linkage map with SSR, AFLP and morphological markers to locate QTLs controlling pathotype-specific powdery mildew resistance in diploid roses. Euphytica 2012, 184, 413–427. [Google Scholar] [CrossRef]
- Smith, C.M.; Boyko, E.V. The molecular bases of plant resistance and defense responses to aphid feeding: Current status. Entomol. Exp. Appl. 2007, 122, 1–16. [Google Scholar] [CrossRef]
- Pollard, D.G. Plant penetration by feeding aphids (Hemiptera, Aphidoidea): A review. Bull. Entomol. Res. 1972, 62, 631–714. [Google Scholar] [CrossRef]
- Miles, P.W. Aphid saliva. Biol. Rev. 1999, 74, 41e85. [Google Scholar] [CrossRef]
- Moloi, M.J.; van der Westhuizen, A.J. The reactive oxygen species are involved in resistance responses of wheat to the Russian wheat aphid. J. Plant Physiol. 2006, 163, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Carolan, J.C.; Fitzroy, C.F.; Ashton, P.D.; Douglas, A.E.; Wilkinson, T.L. The proteome of the pea aphid saliva characterized by LC/MS-MS. Proteomics 2009, 9, 2457–2467. [Google Scholar] [CrossRef] [PubMed]
- Kerchev, P.; Fenton, B.; Foyer, C.H.; Hancock, R.D. Infestation of potato (Solanum tuberosum L.) by the peach-potato aphid (Myzus persicae Sulzer) alters cellular redox status and is influenced by ascorbate. Plant Cell Environ. 2012, 35, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Łukasik, I.; Goławska, S.; Wójcicka, A. Effect of cereal aphid infestation on ascorbate content and ascorbate peroxidase activity in triticale. Pol. J. Environ. Stud. 2012, 21, 1937–1941. [Google Scholar]
- Mai, V.C.; Bednarski, W.; Borowiak-Sobkowiak, B.; Wilkaniec, B.; Samardakiewicz, S.; Morkunas, I. Oxidative stress in pea seedling leaves in response to Acyrthosiphon pisum infestation. Phytochemistry 2013, 93, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Sytykiewicz, H. Differential expression of superoxide dismutase genes in aphid-stressed maize (Zea mays L.) seedlings. PLoS ONE 2014, 9, e94847. [Google Scholar] [CrossRef] [PubMed]
- Moran, P.J.; Cheng, Y.; Cassell, J.L.; Thompson, G.A. Gene expression profiling of Arabidopsis thaliana in compatible plant-aphid interactions. Arch. Insect Biochem. Physiol. 2002, 51, 182–203. [Google Scholar] [CrossRef] [PubMed]
- Divol, F.; Vilaine, F.; Thibivilliers, S.; Amselem, J.; Palauqui, J.C.; Kusiak, C.; Dinant, S. Systemic response to aphid infestation by Myzus persicae in the phloem of Apium graveolens. Plant Mol. Biol. 2005, 57, 517–540. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Huang, Y.; Ayoubi, P. Identification of expression profiles of sorghum genes in response to greenbug phloem-feeding using cDNA subtraction and microarray analysis. Planta 2006, 223, 932–947. [Google Scholar] [CrossRef] [PubMed]
- Delp, G.; Gradin, T.; Ahman, I.; Jonsson, L.M. Microarray analysis of the interaction between the aphid Rhopalosiphum padi and host plants reveals both differences and similarities between susceptible and partially resistant barley lines. Mol. Genet. Genom. 2009, 281, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [PubMed]
- Hoffer, M.; Pscheidt, J.; DeAngelis, J. Controlling Diseases and Aphids on Your Roses; Extension Service; Oregon State University: Corvallis, OR, USA, 2000; pp. 1–7. [Google Scholar]
- Goggin, F.L. Plant–aphid interactions: Molecular and ecological perspectives. Curr. Opin. Plant Biol. 2007, 10, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Ferry, N.; Stavroulakis, S.; Guan, W.; Davison, G.M.; Bell, H.A.; Weaver, R.J.; Down, R.E.; Gatehouse, J.A.; Gatehouse, A.M.R. Molecular interactions between wheat and cereal aphid (Sitobion avenae); analysis of changes to the wheat proteome. Proteomics 2011, 11, 1985–2002. [Google Scholar] [CrossRef] [PubMed]
- Coppola, V.; Coppola, M.; Rocco, M.; Digillio, M.C.; Rocco, M.; Digilio, M.C.; D’Ambrosio, C.; Renzone, G.; Martinelli, R.; Scaloni, A.; et al. Transcriptomic and proteomic analysis of compatible tomato-aphid interaction reveals a predominant salicylic acid-dependent plant response. BMC Genom. 2013, 14, 515. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Ko, C.H.; Wei, H.; Chen, Y.; Jeong, B.R. Physiological and proteomic investigations to study the response of tomato graft unions under temperature stress. PLoS ONE 2016, 11, e0157439. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.G.; Jeong, B.R. Red and Blue Light Emitting Diodes (LEDs) Participate in Mitigation of Hyperhydricity in In Vitro-Grown Carnation Genotypes (Dianthus Caryophyllus). J. Plant Growth Regul. 2017. [Google Scholar] [CrossRef]
- Chow, P.S.; Landhausser, S.M. A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiol. 2004, 24, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Hedge, J.E.; Hofreiter, B.T. Carbohydrate Chemistry; Whistler, R.L., Be Miller, J.N., Eds.; Academic Press: New York, NY, USA, 1962; Volume 17. [Google Scholar]
- Geddes, J.; Eudes, F.; Laroche, A.; Selinger, L.B. Differential expression of proteins in response to the interaction between the pathogen Fusarium graminearum and its host, Hordeum Vulgare. Proteomics 2008, 8, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Li, J.; Liu, H.; Chong, K.; Xu, Y. Roles of ubiquitination-mediated protein degradation in plant responses to abiotic stresses. Environ. Exp. Bot. 2015, 114, 92–103. [Google Scholar] [CrossRef]
- Sharma, B.; Joshi, D.; Bhatt, T.K. Role of ubiquitin-mediated degradation system in plant biology. Front. Plant Sci. 2016, 7, 806. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, Y.; Sako, K.; Maki, Y.; Yamazaki, N.; Yamamoto, H.; Ikeda, A.; Yamaguchi, J. Regulation of leaf organ size by the Arabidopsis RPT2a 19S proteasome subunit. Plant J. 2009, 60, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Dong, X. How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Timperio, A.M.; D’Amici, G.M.; Barta, C.; Loreto, F.; Zolla, L. Proteomics, pigment composition, and organization of thylakoid membranes in iron-deficient spinach leaves. J. Exp. Bot. 2007, 58, 3695–3710. [Google Scholar] [CrossRef] [PubMed]
- Bashir, H.; Ahmad, J.; Bagheri, R.; Nauman, M.; Qureshi, M.I. Limited sulfur resource forces Arabidopsis thaliana to shift towards non-sulfur tolerance under cadmium stress. Environ. Exp. Bot. 2013, 94, 19–32. [Google Scholar] [CrossRef]
- Qureshi, M.I.; D’Amici, G.M.; Fagioni, M.; Rinalducci, S.; Zolla, L. Iron stabilizes thylakoid protein–pigment complexes in Indian mustard during Cd-phytoremediation as revealed by BN-SDS-PAGE and ESI-MS/MS. J. Plant Physiol. 2010, 167, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.G.; Manivannan, A.; Soundararajan, P.; Jeong, B.R. Physiological and proteomic analysis in chloroplasts of Solanum lycopersicum L. under silicon efficiency and salinity stress. Int. J. Mol. Sci. 2014, 15, 21803–21824. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Jeong, B.R. Silicon decreases Fe deficiency responses by improving photosynthesis and maintaining composition of thylakoid multiprotein complex proteins in soybean plants (Glycine max L.). J. Plant Growth Regul. 2015, 34, 485–498. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, B.R.; Lee, J.H.; Kim, T.H. S nutrition alleviates salt stress by maintaining the assemblage of photosynthetic organelles in Kentucky bluegrass (Poa pratensis L.). Plant Growth Regul. 2015, 79, 367–375. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, X.; Lv, Q.; Zhu, D.; Qiu, T.; Xu, Y.; Bao, F.; He, Y.; Hu, Y. Physcomitrella patens dehydrins (PpDHNA and PpDHNC) confer salinity and drought tolerance to transgenic Arabidopsis plants. Front. Plant Sci. 2017, 8, 1316. [Google Scholar] [CrossRef] [PubMed]
- Wind, J.; Smeekens, S.; Hanson, J. Sucrose: Metabolite and signaling molecule. Phytochemistry 2010, 71, 1610–1614. [Google Scholar] [CrossRef] [PubMed]
- Stokes, M.E.; Chattopadhyay, A.; Wilkins, O.; Nambara, E.; Campbell, M.M. Interplay between sucrose and folate modulates auxin signaling in Arabidopsis. Plant Physiol. 2013, 162, 1552–1565. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant 2014, 36, 1607–1619. [Google Scholar] [CrossRef]
- Rosa, M.; Prada, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble sugars—Metabolism, sensing and abiotic stress. Plant Signal. Behav. 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Usadel, B.; Poree, F.; Stitt, M.; Pons-Kühnemann, J.; Sonnewald, U.; Kahmann, R.; et al. Reprogramming a maize plant: Transcriptional and metabolic changes induced by the fungal biotroph Ustilago maydis. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Narozna, D.; Nowak, W.; Samardakiewicz, W.; Remlein-Starosta, D. Cross-talk interactions of sucrose and Fusarium oxysporum in the phenylpropanoid pathway and the accumulation and localization of flavonoids in embryo axes of yellow lupine. J. Plant Physiol. 2011, 168, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Bolouri Moghaddam, M.R.; Van den Eden, W. Sugars and plant innate immunity. J. Exp. Bot. 2012, 63, 3989–3998. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangle, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Gordon-Weeks, R.; Pickett, J.; Ton, J. Natural variation in priming of basal resistance: From evolutionary origin to agricultural exploitation. Mol. Plant Pathol. 2010, 11, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone crosstalk in plant disease and defense: More than just jasmonate–salicylate antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef] [PubMed]
- Chołuj, D.; Moliszewska, E.B. The influence of Aphanomyces cochlioides on selected physiological processes in sugar beet leaves and yield parameters. Eur. J. Plant Pathol. 2012, 132, 59–70. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band No. | Protein Name | Plant Species | Accession Number | Peptides | Biological Function | Mr Value | Sequence Coverage |
|---|---|---|---|---|---|---|---|
| Resistant cultivars | |||||||
| Stella | |||||||
| 1 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 2 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 3 | Probable protein phosphatase | Arabidopsis thaliana | P2C20_ARATH | R. EILHKMK.V | Defense response | 31,827 | 19 |
| 4 | 30S ribosomal protein S11, chloroplastic | Huperzia lucidula | RR11_HUPLU | R. GQAVSWSSAGACGSRGTK.K | Photosynthesis | 14,124 | 43 |
| 5 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | R. GDTILIKGK.K | Cell division | 89,275 | 14 |
| 6 | Shikimate kinase 1, chloroplastic | Oryza sativa | SK1_ORYSJ | MEAGVGLALQSRAAGFGGSDR.R | Phosphorylation | 33,289 | 54 |
| 7 | Pathogenesis-related protein 1 | Asparagus officinalis | AB29G_ARATH | MSSGSWSHEVAVNVAAGRMFK.A | Defense response | 160,195 | 10 |
| Alibaba | |||||||
| 1 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 2 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 3 | Probable protein phosphatase | Arabidopsis thaliana | P2C20_ARATH | R. EILHKMK.V | Defense response | 31,827 | 19 |
| 4 | 30S ribosomal protein S11, chloroplastic | Huperzia lucidula | RR11_HUPLU | R. GQAVSWSSAGACGSRGTK.K | Photosynthesis | 14,124 | 43 |
| 5 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | R. GDTILIKGK.K | Cell division | 89,275 | 14 |
| 6 | Shikimate kinase 1, chloroplastic | Oryza sativa | SK1_ORYSJ | MEAGVGLALQSRAAGFGGSDR.R | Phosphorylation | 33,289 | 54 |
| 7 | Pathogenesis-related protein 1 | Asparagus officinalis | AB29G_ARATH | MSSGSWSHEVAVNVAAGRMFK.A | Defense response | 160,195 | 10 |
| 8 | 1,4-alpha-glucan-branching enzyme | Solanum tuberosum | GLGB_SOLTU | K. VSSGASRNK.I | Starch biosynthesis | 99,021 | 11 |
| Sensitive cultivars | |||||||
| Sun Star | |||||||
| 1 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 2 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 3 | Probable protein phosphatase | Arabidopsis thaliana | P2C20_ARATH | R. EILHKMK.V | Defense response | 31,827 | 19 |
| 4 | 30S ribosomal protein S11, chloroplastic | Huperzia lucidula | RR11_HUPLU | R. GQAVSWSSAGACGSRGTK.K | Photosynthesis | 14,124 | 43 |
| 5 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | R. GDTILIKGK.K | Cell division | 89,275 | 14 |
| 6 | Shikimate kinase 1, chloroplastic | Oryza sativa | SK1_ORYSJ | MEAGVGLALQSRAAGFGGSDR.R | Phosphorylation | 33,289 | 54 |
| 7 | Pathogenesis-related protein 1 | Asparagus officinalis | AB29G_ARATH | MSSGSWSHEVAVNVAAGRMFK.A | Defense response | 160,195 | 10 |
| 8 | 1,4-alpha-glucan-branching enzyme | Solanum tuberosum | GLGB_SOLTU | K. VSSGASRNK.I | Starch biosynthesis | 99,021 | 11 |
| Haetsal | |||||||
| 1 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 2 | armadillo/beta-catenin repeat family protein | Populus trichocarpa | gi|224066241 | METSSVRCLINSISR.F | ubiquitin-protein transferase activity | 83,169 | 12 |
| 3 | Probable protein phosphatase | Arabidopsis thaliana | P2C20_ARATH | R. EILHKMK.V | Defense response | 31,827 | 19 |
| 4 | 30S ribosomal protein S11, chloroplastic | Huperzia lucidula | RR11_HUPLU | R. GQAVSWSSAGACGSRGTK.K | Photosynthesis | 14,124 | 43 |
| 5 | Cell division cycle protein 48 homolog | Capsicum annuum | CDC48_CAPAN | R. GDTILIKGK.K | Cell division | 89,275 | 14 |
| 6 | Shikimate kinase 1, chloroplastic | Oryza sativa | SK1_ORYSJ | MEAGVGLALQSRAAGFGGSDR.R | Phosphorylation | 33,289 | 54 |
| 7 | Pathogenesis-related protein 1 | Asparagus officinalis | AB29G_ARATH | MSSGSWSHEVAVNVAAGRMFK.A | Defense response | 160,195 | 10 |
| 8 | 1,4-alpha-glucan-branching enzyme | Solanum tuberosum | GLGB_SOLTU | K. VSSGASRNK.I | Starch biosynthesis | 99,021 | 11 |
| 9 | Probable fructokinase-5 | Arabidopsis thaliana | SCRK5_ARATH | K. APGGAPANVACAITKLGGK.S | Sugar synthesis | 34,666 | 27 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muneer, S.; Jeong, H.K.; Park, Y.G.; Jeong, B.R. Proteomic Analysis of Aphid-Resistant and -Sensitive Rose (Rosa Hybrida) Cultivars at Two Developmental Stages. Proteomes 2018, 6, 25. https://doi.org/10.3390/proteomes6020025
Muneer S, Jeong HK, Park YG, Jeong BR. Proteomic Analysis of Aphid-Resistant and -Sensitive Rose (Rosa Hybrida) Cultivars at Two Developmental Stages. Proteomes. 2018; 6(2):25. https://doi.org/10.3390/proteomes6020025
Chicago/Turabian StyleMuneer, Sowbiya, Hai Kyoung Jeong, Yoo Gyeong Park, and Byoung Ryong Jeong. 2018. "Proteomic Analysis of Aphid-Resistant and -Sensitive Rose (Rosa Hybrida) Cultivars at Two Developmental Stages" Proteomes 6, no. 2: 25. https://doi.org/10.3390/proteomes6020025
APA StyleMuneer, S., Jeong, H. K., Park, Y. G., & Jeong, B. R. (2018). Proteomic Analysis of Aphid-Resistant and -Sensitive Rose (Rosa Hybrida) Cultivars at Two Developmental Stages. Proteomes, 6(2), 25. https://doi.org/10.3390/proteomes6020025