
Fifty Years of Clinical Application of Newcastle Disease Virus: Time to Celebrate!


Abstract
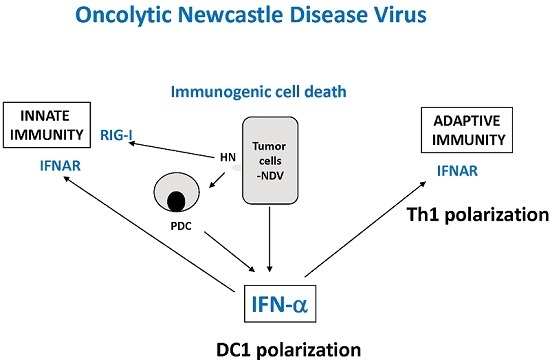
:
1. Introduction
2. Treatment of Cancer Patients by NDV (Newcastle Disease Virus) Oncolysate Vaccines
3. Treatment of Cancer Patients by Oncolytic NDV
4. Treatment of Cancer Patients by Autologous Tumor Cell Vaccine ATV-NDV
4.1. From Mouse to Man: Translational Research in the 1990s
4.2. Clinical Studies with ATV-NDV
5. Treatment of Cancer Patients by ATV-NDV with Attached Bispecific Antibody αHNxαCD28
6. Treatment of Cancer Patients by Autologous Viral Oncolysate-Pulsed DC (VOL-DC)
7. Results from China
8. Special Characteristics of NDV and Other Paramyxoviruses
8.1. High Safety Profile
8.2. Tumor-Selective Replication and Oncolysis
8.3. Immunogenic Cell Death (ICD)
8.4. Immune Response
8.5. Potential to Break Therapy Resistancies
9. Why Has It Taken So Long?
10. Summary
Acknowledgments
Conflicts of Interest
References
- Wheelock, E.F.; Dingle, J.H. Observations on the repeated administration of viruses to a patient with acute leukemia, a preliminary report. N. Engl. J. Med. 1964, 271, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, A.; Lindenmann, J. Virus interference. I. The interferon. Proc. R. Soc. Lond. B Biol. Sci. 1957, 147, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Lindenmann, J. Viruses as immunological adjuvants in cancer. Biochim. Biophys. Acta 1974, 355, 49–75. [Google Scholar] [PubMed]
- Sinkovics, J.G.; Horvath, J.C. Newcastle disease virus (NDV). Brief history of its oncolytic strains. J. Clin. Virol. 2000, 16, 1–15. [Google Scholar] [CrossRef]
- Cassel, W.A.; Garrett, R.E. Newcastle disease virus as an antineoplastic agent. Cancer 1965, 18, 863–868. [Google Scholar] [CrossRef]
- Cassel, W.A.; Garrett, R.E. Tumor immunity after viral oncolysis. J. Bacteriol. 1966, 92, 792. [Google Scholar] [PubMed]
- Murray, D.R.; Cassel, W.A.; Torbin, A.H.; Olkowski, Z.L.; Moore, M.E. Viral oncolysate in the management of malignant melanoma. II. Clinical studies. Cancer 1977, 40, 680–686. [Google Scholar] [CrossRef]
- Cassel, W.A.; Murray, D.R. A ten-year follow-up on stage II malignant melanoma patients treated postsurgically with Newcastle disease virus oncolysate. Med. Oncol. Tumor Pharmacother. 1992, 9, 169–171. [Google Scholar] [PubMed]
- Batliwalla, F.M.; Bateman, B.A.; Serrano, D.; Murray, D.; Macphail, S.; Maino, V.C.; Ansel, J.C.; Gregersen, P.K.; Armstrong, C.A. A 15-year follow-up of AJCC stage III malignant melanoma patients treated postsurgically with Newcastle disease virus (NDV) oncolysate and determination of alterations in the CD8 T cell repertoire. Mol. Med. 1998, 4, 783–794. [Google Scholar] [PubMed]
- Kirchner, H.H.; Anton, P.; Atzpodien, J. Adjuvant treatment of locally advanced renal cancer with autologous virus-modified tumor vaccines. World J. Urol. 1995, 13, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Csatary, L.K.; Eckhardt, S.; Bukosza, I.; Czegledi, F.; Fenyvesi, C.; Gergely, P.; Bodey, B.; Csatary, C.M. Attenuated veterinary virus vaccine for the treatment of cancer. Cancer Detect. Prev. 1993, 17, 619–627. [Google Scholar] [PubMed]
- Csatary, L.K.; Gosztonyi, G.; Szeberenyi, J.; Fabian, Z.; Liszka, V.; Bodey, B.; Csatary, C.M. MTH-68/H oncolytic viral treatment in human high-grade gliomas. J. Neurooncol. 2004, 67, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Freeman, A.I.; Zakay-Rones, Z.; Gomori, J.M.; Linetsky, E.; Rasooly, L.; Greenbaum, E.; Rozenman-Yair, S.; Panet, A.; Libson, E.; Irving, C.S.; et al. Phase I/II trial of intravenous NDV-HUJ oncolytic virus in recurrent glioblastoma multiforme. Mol. Ther. 2006, 13, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Lorence, R.M.; Rood, P.A.; Kelley, K.W. Newcastle disease virus as an antineoplastic agent: Induction of tumor necrosis factor-alpha and augmentation of its cytotoxicity. J. Natl. Cancer Inst. 1988, 80, 1305–1312. [Google Scholar] [CrossRef] [PubMed]
- Reichard, K.W.; Lorence, R.M.; Cascino, C.J.; Peeples, M.E.; Walter, R.J.; Fernando, M.B.; Reyes, H.M.; Greager, J.A. Newcastle disease virus selectively kills human tumor cells. J. Surg. Res. 1992, 52, 448–453. [Google Scholar] [CrossRef]
- Phuangsab, A.; Lorence, R.M.; Reichard, K.W.; Peeples, M.E.; Walter, R.J. Newcastle disease virus therapy of human tumor xenografts: Antitumor effects of local or systemic administration. Cancer Lett. 2001, 172, 27–36. [Google Scholar] [CrossRef]
- Pecora, A.L.; Rizvi, N.; Cohen, G.I.; Meropol, N.J.; Sterman, D.; Marshall, J.L.; Goldberg, S.; Gross, P.; O’Neil, J.D.; Groene, W.S.; et al. Phase I trial of intravenous administration of PV701, an oncolytic virus, in patients with advanced solid cancers. J. Clin. Oncol. 2002, 20, 2251–2266. [Google Scholar] [CrossRef] [PubMed]
- Lorence, R.M.; Pecora, A.L.; Major, P.P.; Hotte, S.J.; Laurie, S.A.; Roberts, M.S.; Groene, W.S.; Bamat, M.K. Overview of phase I studies of intravenous administration of PV701, an oncolytic virus. Curr. Opin. Mol. Ther. 2003, 5, 618–624. [Google Scholar] [PubMed]
- Kobayashi, H. Viral xenogenisation of intact tumor cells. Adv. Cancer Res. 1979, 30, 279–299. [Google Scholar] [PubMed]
- Heicappell, R.; Schirrmacher, V.; von Hoegen, P.; Ahlert, T.; Appelhans, B. Prevention of metastatic spread by postoperative immunotherapy with virally modified autologous tumor cells. I. Parameters for optimal therapeutic effects. Int. J. Cancer 1986, 37, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Heicappell, R. Prevention of metastatic spread by postoperative immunotherapy with virally modified autologous tumor cells. II. Establishment of specific systemic immunity. Clin. Exp. Metastab. 1987, 5, 147–156. [Google Scholar] [CrossRef]
- Von Hoegen, P.; Weber, E.; Schirrmacher, V. Modification of tumor cells by a low dose of Newcastle disease virus. I. Augmentation of the tumor-specific T cell response in the absence of an anti-viral response. Eur. J. Immunol. 1988, 18, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Schild, H.; von Hoegen, P.; Schirrmacher, V. Modification of tumor cells by a low dose of Newcastle disease virus. II. Augmented tumor-specific T cell responses as a result of CD4+ and CD8+ immune T cell cooperation. Cancer Immunol. Immunother. 1989, 28, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Von Hoegen, P.; Zawatzky, R.; Schirrmacher, V. Modification of tumor cells by a low dose of Newcastle disease virus. III. Potentiation of tumor-specific cytotoxic T cell activity via induction of interferon-alpha/beta. Cell. Immunol. 1990, 126, 80–90. [Google Scholar] [CrossRef]
- Hanna, M.G.; Pollack, V.A.; Peters, L.C.; Hoover, H.C. Active specific immunotherapy of established micrometastases with BCG plus tumor cell vaccines. Cancer 1982, 49, 659–664. [Google Scholar] [CrossRef]
- Hoover, H.C., Jr.; Brandhorst, J.S.; Peters, L.C.; Surdyke, M.G.; Takeshita, Y.; Madariaga, J.; Muenz, L.R.; Hanna, M.G., Jr. Adjuvant active-specific immunotherapy for human colorectal cancer: 6,5-year follow-up of a phase III prospectively randomized trial. J. Clin. Oncol. 1993, 11, 390–399. [Google Scholar] [PubMed]
- Schirrmacher, V.; Haas, C.; Bonifer, R.; Ahlert, T.; Gerhards, R.; Ertel, C. Human tumor cell modification by virus infection: An efficient and safe way to produce cancer vaccine with pleiotropic immune stimulatory properties when using Newcastle disease virus. Gene Ther. 1999, 6, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Bohle, W.; Schlag, P.; Liebrich, W.; Hohenberger, P.; Manasterski, M.; Möller, P.; Schirrmacher, V. Postoperative active specific immunization in colorectal cancer patients with virus-modified autologous tumor cell vaccine. First clinical results with tumor-cell vaccines modified with live but avirulent Newcastle disease virus. Cancer 1990, 66, 1517–1523. [Google Scholar] [CrossRef]
- Schirrmacher, V. Cancer-reactive memory T cells from bone marrow: Spontaneous induction and therapeutic potential (Review). Int. J. Oncol. 2015, 47, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Ahlert, T.; Sauerbrei, W.; Bastert, G.; Ruhland, S.; Bartik, B.; Simiantonaki, N.; Schumacher, J.; Häcker, B.; Schumacher, M.; Schirrmacher, V. Tumor-cell number and viability as quality and efficacy parameters of autologous virus-modified cancer vaccines in patients with breast or ovarian cancer. J. Clin. Oncol. 1997, 15, 2763. [Google Scholar]
- Karcher, J.; Dyckhoff, G.; Beckhove, P.; Reisser, C.; Brysch, M.; Ziouta, Y.; Helmke, B.H.; Weidauer, H.; Schirrmacher, V.; Herold-Mende, C. Antitumor vaccination in patients with head and neck squamous cell carcinomas with autologous virus-modified tumor cells. Cancer Res. 2004, 64, 8057–8061. [Google Scholar] [CrossRef] [PubMed]
- Ockert, D.; Schirrmacher, V.; Beck, N.; Stoelben, E.; Ahlert, T.; Flechtenmacher, J.; Hagmüller, E.; Buchcik, R.; Nagel, M.; Saeger, H.D. Newcastle disease virus-infected intact autologous tumor cell vaccine for adjuvant active specific immunotherapy of resected colorectal carcinoma. Clin. Cancer Res. 1996, 2, 21–28. [Google Scholar] [PubMed]
- Pomer, S.; Schirrmacher, V.; Thiele, R.; Lohrke, H.; Brkovic, D.; Staehler, G. Tumor response and 4 year survival-data of patients with advanced renal-cell carcinoma treated with autologous tumor vaccine and subcutaneous r-IL-2 and IFN-alpha(2b). Int. J. Oncol. 1995, 6, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.H.; Bonsanto, M.M.; Beckhove, P.; Brysch, M.; Geletneky, K.; Ahmadi, R.; Schuele-Freyer, R.; Kremer, P.; Ranaie, G.; Matejic, D.; et al. Antitumor vaccination of patients with glioblastoma multiforme: A pilot study to assess feasibility, safety, and clinical benefit. J. Clin. Oncol. 2004, 22, 4272–4281. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.; Kemmner, W.; Weitz, J.; Wernecke, K.D.; Schirrmacher, V.; Schlag, P.M. Efficiency of adjuvant active specific immunization with Newcastle disease virus modified tumor cells in colorectal cancer patients following resection of liver metastases: Results of a prospective randomized trial. Cancer Immunol. Immunother. 2009, 58, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Fournier, P.; Schlag, P. Autologous tumor cell vaccines for post-operative active-specific immunotherapy of colorectal carcinoma: Long-term patient survival and mechanism of function. Expert Rev. Vaccines 2014, 13, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.; Schirrmacher, V. Bispecific antibodies and trispecific immunocytokines for targeting the immune system against cancer: Preparing for the future. BioDrugs 2013, 27, 35–53. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Bihari, A.S.; Stücker, W.; Sprenger, T. Long-term remission of prostate cancer with extensive bone metastases upon immuno- and virotherapy: A case report. Oncol. Lett. 2014, 8, 2403–2406. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Wang, H.; Sun, T.M.; Yao, W.Q.; Chen, L.L.; Jin, Y.; Li, C.L.; Meng, F.J. Application of autologous tumor cell vaccine and NDV vaccine in treatment of tumors of the digestive tract. World J. Gastroenterol. 2003, 9, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Wilden, H.; Fournier, P.; Peeters, B.; Schirrmacher, V. In vivo efficacy of systemic tumor targeting of a viral RNA vector with oncolytic properties using a bispecific adapter protein. Int. J. Oncol. 2006, 29, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Wei, D.; Nan, G.; Cui, S.J.; Chen, Z.N.; Bian, H. Construction of a minigenome rescue system for Newcastle disease virus strain Italien. Arch. Virol. 2011, 156, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Sun, N.; Nan, G.; Wang, Y.; Liu, H.Q.; Peeters, B.; Chen, Z.N.; Bian, H. Construction of recombinant Newcastle disease virus Italien strain for oncolytic virotherapy of tumors. Hum. Gene Ther. 2012, 23, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Li, Q.; Wang, X.L.; Wang, Y.; Xu, J.; Feng, F.; Nan, G.; Wang, B.; Li, C.; Guo, T.; et al. Oncolytic Newcastle disease virus expressing chimeric antibody enhanced anti-tumor efficacy in orthotopic hepatoma-bearing mice. J. Exp. Clin. Cancer Res. 2015, 34, 153. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Wu, J.; Wei, D.; Zhang, D.W.; Feng, H.; Chen, Z.N.; Bian, H. Newcastle disease virus represses the activation of human hepatic stellate cells and reverses the development of hepatic fibrosis in mice. Liver Int. 2009, 29, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Zhang, T.Y.; Yin, J.C.; Wang, H.; Sun, T.; Chen, L.Q.; Bai, F.L.; Wu, W.; Ren, G.P.; Li, D.S. Enhancement of anti-tumor activity of Newcastle disease virus by the synergistic effect of cytosine deaminase. Asian Pac. J. Cancer Prev. 2013, 14, 7489–7496. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Koopmann, J.; Fiola, C.; Fournier, P.; Schirrmacher, V. Dendritic cells pulsed with viral oncolysates potently stimulate autologous T cells from cancer patients. Int. J. Oncol. 2002, 21, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Fournier, P.; Schirrmacher, V. Induction of interferon-alpha and tumor necrosis factor-related apoptosis-inducing ligand in human blood mononuclear cells by hemagglutinin-neuraminidase but not F protein of Newcastle disease virus. Virology 2002, 297, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Schirrmacher, V.; Fournier, P. The hemagglutinin-neuraminidase gene of Newcastle Disease Virus: A powerfull molecular adjuvant for DNA anti-tumor vaccination. Vaccine 2010, 28, 6891–6900. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Z.; Wu, Y.; Zheng, D.; Sun, C.; Bi, D.; Zuo, Y.; Xu, T. Molecular epidemiological analysis of Newcastle disease virus isolated in China in 2005. J. Virol. Methods 2007, 140, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Westover, K.M.; Hughes, A.L. Molecular evolution of viral fusion and matrix protein genes and phylogenetic relationships among the Paramyxoviridae. Mol. Phylogenet. Evol. 2001, 21, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Ahlert, T.; Schirrmacher, V. Isolation of a human melanoma adapted Newcastle disease virus mutant with highly selective replication patterns. Cancer Res. 1990, 50, 5962–5968. [Google Scholar] [PubMed]
- Fàbiàn, Z.; Csatary, C.M.; Szeberényi, J.; Csatary, L.K. P53-independent endoplasmic reticulum stress-mediated cytotoxicity of a Newcasle disease virus strain in tumor cell lines. J. Virol. 2007, 81, 2817–2830. [Google Scholar] [CrossRef] [PubMed]
- Elankumaran, S.; Rockeman, D.; Samal, S.K. Newcastle disease virus exerts oncolysis by both intrinsic and extrinsic caspase-dependent pathways of cell death. J. Virol. 2006, 80, 7522–7534. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V. Oncolytic Newcastle disease virus as a prospective anti-cancer therapy. A biological agent with potential to break therapy resistance. Expert Opin. Biol. Ther. 2015, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Garcia-Sastre, A.; Cros, J.F.; Basler, C.F.; Palese, P. Newcastle disease virus V protein is a determinant of host range restriction. J. Virol. 2003, 77, 9522–9532. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.A.; Batich, K.A.; Gunn, M.D.; Huang, M.-N.; Sanchez-Perez, L.; Nair, S.K.; Congdon, K.L.; Reap, E.A.; Archer, G.E.; Desjardins, A.; et al. Tetanus toxoid and CCL3 improve dendritic cell vaccines in mice and glioblastoma patients. Nature 2015, 519, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Marchini, A.; Scott, E.M.; Rommelare, J. Overcoming barriers in oncolytic virotherapy with HDAC inhibitors and immune checkpoint blockade. Viruses 2016, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Wilden, H.; Fournier, P.; Zawatzky, R.; Schirrmacher, V. Expression of RIG-I, IRF3, IFN-beta and IRF7 determines resistance or susceptibility of cells to infection by Newcastle disease virus. Int. J. Oncol. 2009, 34, 971–982. [Google Scholar] [PubMed]
- Jarahian, M.; Watzl, C.; Fournier, P.; Arnold, A.; Djandji, D.; Zahedi, S.; Cerwenka, A.; Paschen, A.; Schirrmacher, V.; Momburg, F. Activation of natural killer cells by Newcastle disease virus hemagglutinin-neuraminidase. J. Virol. 2009, 83, 8108–8121. [Google Scholar] [CrossRef] [PubMed]
- Koks, C.A.; Garg, A.D.; Ehrhardt, M.; Riva, M.; Vandenberk, L.; Boon, L.; De Vleeschouwer, S.; Agostinis, P.; Graf, N.; Van Gool, S.W. Newcastle disease virotherapy induces long-term survival and tumor-specific immune memory in orthotopic glioma through the induction of immunogenic cell death. Int. J. Cancer 2015, 136, E313–E325. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.; Wilden, H.; Schirrmacher, V. Importance of retinoic acid-inducible gene I and of receptor for type I interferon for cellular resistance to infection by Newcastle disease virus. Int. J. Oncol. 2012, 40, 287–298. [Google Scholar] [PubMed]
- Zaslavsky, E.; Hershberg, U.; Seto, J.; Pham, A.M.; Marquez, S.; Duke, J.L.; Wetmur, J.G.; Tenoever, B.R.; Sealfon, S.C.; Kleinstein, S.H. Antiviral response dictated by choreographed cascade of transcription factors. J. Immunol. 2010, 184, 2908–2917. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, F.; Lapenta, C.; Donati, S.; Abalsamo, L.; Barnaba, V.; Belardelli, F.; Santini, S.M.; Ferrantini, M. IFN-α enhances cross-presentation in human dendritic cells by modulating antigen survival, endocytic routing, and processing. Blood 2012, 119, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Washburn, B.; Schirrmacher, V. Human tumor cell infection by Newcastle disease virus leads to upregulation of HLA and cell adhesion molecules and to induction of interferons, chemokines and finally apoptosis. Int. J. Oncol. 2002, 21, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Fiola, C.; Peeters, B.; Fournier, P.; Arnold, A.; Bucur, M.; Schirrmacher, V. Tumor selective replication of Newcastle disease virus: Association with defects of tumor cells in antiviral defense. Int. J. Cancer 2006, 119, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.; Palese, P.; Zamarin, D. Oncolytic specificity of Newcastle disease virus is mediated by selectively for apoptosis-resistant cells. J. Virol. 2011, 85, 6015–6023. [Google Scholar] [CrossRef] [PubMed]
- Ch’ng, W.C.; Stanbridge, E.J.; Yussoff, K.; Shafee, N. The oncolytic activity of Newcastle disease virus in clear cell carcinoma cells in normoxic and hypoxic conditions. The interplay between VHL and interferon-beta signaling. J. Interferon Cytokine Res. 2013, 33, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Schlude, C.; Weitz, J.; Beckhove, P. Strong T-cell costimulation can reactivate tumor antigen-specific T cells in late-stage metastasized colorectal carcinoma patients: Results from a phase I clinical study. Int. J. Oncol. 2015, 46, 71–77. [Google Scholar] [PubMed]
- Termeer, C.C.; Schirrmacher, V.; Bröcker, E.B.; Becker, J.C. Newcastle disease virus infection induces B7-1/B7-2-independent T-cell costimulatory activity in human melanoma cells. Cancer Gene Ther. 2000, 7, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Zamarin, D.; Holmgaard, R.B.; Subudhi, S.K.; Park, J.S.; Mansour, M.; Palese, P.; Merghoub, T.; Wolchok, J.D.; Allison, J.P. Localized oncolytic virotherapy overcomes systemic tumor resistance to immune checkpoint blockade immunotherapy. Sci. Transl. Med. 2014, 6, 226–332. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. China approves world’s first oncolytic virus therapy for cancer treatment. J. Natl. Cancer Inst. 2006, 98, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Andtbacka, R.H.; Kaufman, H.L.; Collichio, F.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.A.; Spitler, L.E.; Puzanov, I.; Agarwala, S.S.; et al. Talimogene laherparepvec improves durable response rate in patients with advanced melanoma. J. Clin. Oncol. 2015, 33, 2780–2788. [Google Scholar] [CrossRef] [PubMed]
- Puzanov, I.; Milhem, M.M.; Minor, D.; Hamid, O.; Li, A.; Chen, L.; Chastain, M.; Gorski, K.S.; Anderson, A.; Chou, J.; et al. Talimogene laherparepvec in combination with Ipilimumab in previously untreated, unresectable stage IIIB-IV melanoma. J. Clin. Oncol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.J. Scientific interest in Newcastle disease virus is reviving. J. Natl. Cancer Inst. 1999, 91, 1708–1710. [Google Scholar] [CrossRef] [PubMed]
- Schirrmacher, V.; Stücker, W.; Lulei, M.; Bihari, A.S.; Sprenger, T. Long-term survival of a breast cancer patient with extensive liver metastases upon immune and virotherapy: A case report. Immunotherapy 2015, 7, 855–860. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Time Period | Basic Research | Clinical Application | References |
|---|---|---|---|
| 1960s | viral oncolysis, post-oncol immunity | single case observations | [1,2,3] |
| 1970s | studies on mechanisms; viral oncolysate | post-op vaccination studies | [5,6,7,8,9,19] |
| 1980s | ATV-NDV live cell vaccine | vaccination with viral oncolysates | [14,20,21,22,23,24] |
| 1990s | genome sequencing, molecular studies | phase I–II studies (ATV-NDV) | [27,28,30,32,33] |
| immune cell activation mechanisms | vaccination by inhalation | [11,41,42,51] | |
| 2000s | recombinant improved NDV strains | phase II/III studies | [31,34,35,39,40] |
| - | systemic dose-escalation study | [17,18,44,46,53] | |
| 2010s | ICD mechanisms | VOL-DC, compassionate use studies | [36,37,38,48,60,61,62,68,75] |
| high quality (GMP) virus production | - | [41,42,43,60,66,67,70] |
© 2016 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schirrmacher, V. Fifty Years of Clinical Application of Newcastle Disease Virus: Time to Celebrate! Biomedicines 2016, 4, 16. https://doi.org/10.3390/biomedicines4030016
Schirrmacher V. Fifty Years of Clinical Application of Newcastle Disease Virus: Time to Celebrate! Biomedicines. 2016; 4(3):16. https://doi.org/10.3390/biomedicines4030016
Chicago/Turabian StyleSchirrmacher, Volker. 2016. "Fifty Years of Clinical Application of Newcastle Disease Virus: Time to Celebrate!" Biomedicines 4, no. 3: 16. https://doi.org/10.3390/biomedicines4030016