
Stem-Cell-Driven Chondrogenesis: Perspectives on Amnion-Derived Cells
 ,
,  , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Scientometric Analysis
3. Pathways Regulating Chondrogenesis
3.1. Chondrogenesis during Embryogenesis
3.2. Morphogenic Pathways: TGF-β/BMP and Shh Pathways
3.3. Morphogenic Pathways: Wnt/β-Catenin Pathways
3.4. Other Mechanisms
3.4.1. Low O2 Tension
3.4.2. Epigenetic Regulation
4. Failure of Chondrogenic Mechanisms in Hypertrophic Chondro-Healing
5. Amnion-Derived Stem Cells in Chondrogenesis
In Vitro Amnion-Derived Stem Cells Lesson on Chondro-Related Pathways
6. New Strategies to Induce Chondrogenesis
6.1. 3D High-Density Cultures
6.2. Role of 3D Scaffold in Stem Cell Chondrogenic Differentiation
6.3. New Perspectives: Organoids
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Camarero-Espinosa, S.; Rothen-Rutishauser, B.; Foster, E.J.; Weder, C. Articular Cartilage: From Formation to Tissue Engineering. Biomater. Sci. 2016, 4, 734–767. [Google Scholar] [CrossRef] [PubMed]
- Messaoudi, O.; Henrionnet, C.; Bourge, K.; Loeuille, D.; Gillet, P.; Pinzano, A. Stem Cells and Extrusion 3D Printing for Hyaline Cartilage Engineering. Cells 2020, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Flik, K.R.; Verma, N.; Cole, B.J.; Bach, B.R. Articular Cartilage: Structure, Biology, and Function. In Cartilage Repair Strategies; Williams, R.J., Ed.; Humana Press: Totowa, NJ, USA, 2007; pp. 1–12. ISBN 978-1-58829-629-0. [Google Scholar]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, R.A. The Cell Density of Human Articular and Costal Cartilage. J. Anat. 1967, 101, 753–763. [Google Scholar] [PubMed]
- Huber, M.; Trattnig, S.; Lintner, F. Anatomy, Biochemistry, and Physiology of Articular Cartilage. Investig. Radiol. 2000, 35, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, A.M.; Richardson, J.B. Articular Cartilage: Structure, Injuries and Review of Management. Br. Med. Bull. 2008, 87, 77–95. [Google Scholar] [CrossRef] [PubMed]
- Carballo, C.B.; Nakagawa, Y.; Sekiya, I.; Rodeo, S.A. Basic Science of Articular Cartilage. Clin. Sports Med. 2017, 36, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.P.; Foster, R.J.; Mow, V.C. Composition and Dynamics of Articular Cartilage: Structure, Function, and Maintaining Healthy State. J. Orthop. Sports Phys. Ther. 1998, 28, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.; Weis, M.; Wu, J.-J. Articular Cartilage Collagen: An Irreplaceable Framework? eCM 2006, 12, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.B.; Knudson, W. Cartilage Proteoglycans. Semin. Cell Dev. Biol. 2001, 12, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Armiento, A.R.; Alini, M.; Stoddart, M.J. Articular Fibrocartilage—Why Does Hyaline Cartilage Fail to Repair? Adv. Drug Deliv. Rev. 2019, 146, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Muthu, S.; Korpershoek, J.V.; Novais, E.J.; Tawy, G.F.; Hollander, A.P.; Martin, I. Failure of Cartilage Regeneration: Emerging Hypotheses and Related Therapeutic Strategies. Nat. Rev. Rheumatol. 2023, 19, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Wuelling, M.; Vortkamp, A. Chondrocyte Proliferation and Differentiation. In Endocrine Development; Camacho-Hübner, C., Nilsson, O., Sävendahl, L., Eds.; S. Karger AG: Basel, Switzerland, 2011; Volume 21, pp. 1–11. ISBN 978-3-8055-9792-0. [Google Scholar]
- Schofield, D.; Cunich, M.; Shrestha, R.N.; Tanton, R.; Veerman, L.; Kelly, S.; Passey, M.E. The Long-Term Economic Impacts of Arthritis through Lost Productive Life Years: Results from an Australian Microsimulation Model. BMC Public Health 2018, 18, 654. [Google Scholar] [CrossRef] [PubMed]
- Adkar, S.S.; Brunger, J.M.; Willard, V.P.; Wu, C.-L.; Gersbach, C.A.; Guilak, F. Genome Engineering for Personalized Arthritis Therapeutics. Trends Mol. Med. 2017, 23, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Makris, E.A.; Gomoll, A.H.; Malizos, K.N.; Hu, J.C.; Athanasiou, K.A. Repair and Tissue Engineering Techniques for Articular Cartilage. Nat. Rev. Rheumatol. 2015, 11, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B.R.; Mooney, D.J. Biomaterials to Mimic and Heal Connective Tissues. Adv. Mater. 2019, 31, 1806695. [Google Scholar] [CrossRef] [PubMed]
- Nazempour, A.; Van Wie, B.J. Chondrocytes, Mesenchymal Stem Cells, and Their Combination in Articular Cartilage Regenerative Medicine. Ann. Biomed. Eng. 2016, 44, 1325–1354. [Google Scholar] [CrossRef] [PubMed]
- Statham, P.; Jones, E.; Jennings, L.M.; Fermor, H.L. Reproducing the Biomechanical Environment of the Chondrocyte for Cartilage Tissue Engineering. Tissue Eng. Part B Rev. 2022, 28, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Martel-Pelletier, J.; Barr, A.J.; Cicuttini, F.M.; Conaghan, P.G.; Cooper, C.; Goldring, M.B.; Goldring, S.R.; Jones, G.; Teichtahl, A.J.; Pelletier, J.-P. Osteoarthritis. Nat. Rev. Dis. Prim. 2016, 2, 16072. [Google Scholar] [CrossRef]
- Taruc-Uy, R.L.; Lynch, S.A. Diagnosis and Treatment of Osteoarthritis. Prim. Care Clin. Off. Pract. 2013, 40, 821–836. [Google Scholar] [CrossRef]
- Robinson, P.D.; McEwan, J.; Adukia, V.; Prabhakar, M. Osteoarthritis and Arthroplasty of the Hip and Knee. Br. J. Hosp. Med. 2018, 79, C54–C59. [Google Scholar] [CrossRef] [PubMed]
- Mahla, R.S. Stem Cells Applications in Regenerative Medicine and Disease Therapeutics. Int. J. Cell Biol. 2016, 2016, 6940283. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, W.; Dobrzyński, M.; Szymonowicz, M.; Rybak, Z. Stem Cells: Past, Present, and Future. Stem Cell Res. Ther. 2019, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Maraldi, T.; Russo, V. Amniotic Fluid and Placental Membranes as Sources of Stem Cells: Progress and Challenges. Int. J. Mol. Sci. 2022, 23, 5362. [Google Scholar] [CrossRef] [PubMed]
- Torre, P.D.; Flores, A.I. Current Status and Future Prospects of Perinatal Stem Cells. Genes 2020, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Z.; Liu, N. The Stemness of Perinatal Stem Cells. In Perinatal Stem Cells; Han, Z.C., Takahashi, T.A., Han, Z., Li, Z., Eds.; Springer: Singapore, 2019; pp. 27–37. ISBN 9789811327025. [Google Scholar]
- Miki, T. Amnion-Derived Stem Cells: In Quest of Clinical Applications. Stem Cell Res. Ther. 2011, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; De Crombrugghe, B. Sox9 Is Required for Cartilage Formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Smits, P.; Li, P.; Mandel, J.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; De Crombrugghe, B.; Lefebvre, V. The Transcription Factors L-Sox5 and Sox6 Are Essential for Cartilage Formation. Dev. Cell 2001, 1, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Chaboissier, M.-C.; Martin, J.F.; Schedl, A.; De Crombrugghe, B. The Transcription Factor Sox9 Has Essential Roles in Successive Steps of the Chondrocyte Differentiation Pathway and Is Required for Expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, L.; Xu, X.; Li, C.; Huang, C.; Deng, C.-X. TGF-β/Smad3 Signals Repress Chondrocyte Hypertrophic Differentiation and Are Required for Maintaining Articular Cartilage. J. Cell Biol. 2001, 153. [Google Scholar] [CrossRef]
- Li, T.-F.; Darowish, M.; Zuscik, M.J.; Chen, D.; Schwarz, E.M.; Rosier, R.N.; Drissi, H.; O’Keefe, R.J. Smad3-Deficient Chondrocytes Have Enhanced BMP Signaling and Accelerated Differentiation. J. Bone Min. Res. 2005, 21, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Kawakatsu, M.; Kanno, S.; Gui, T.; Gai, Z.; Itoh, S.; Tanishima, H.; Oikawa, K.; Muragaki, Y. Loss of Smad3 Gives Rise to Poor Soft Callus Formation and Accelerates Early Fracture Healing. Exp. Mol. Pathol. 2011, 90, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Johnson, M.; Filvaroff, E.H.; LaBorde, J.; Sheehan, D.M.; Derynck, R.; Moses, H.L. Expression of a Truncated, Kinase-Defective TGF-β Type II Receptor in Mouse Skeletal Tissue Promotes Terminal Chondrocyte Differentiation and Osteoarthritis. J. Cell Biol. 1997, 139, 541–552. [Google Scholar] [CrossRef]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone Morphogenetic Protein-4 Is Required for Mesoderm Formation and Patterning in the Mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bradley, A. Mice Deficient for BMP2 Are Nonviable and Have Defects in Amnion/Chorion and Cardiac Development. Development 1996, 122, 2977–2986. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.; Tsuji, K.; Cox, K.; Harfe, B.D.; Rosen, V.; Tabin, C.J. Genetic Analysis of the Roles of BMP2, BMP4, and BMP7 in Limb Patterning and Skeletogenesis. PLoS Genet. 2006, 2, e216. [Google Scholar] [CrossRef] [PubMed]
- Retting, K.N.; Song, B.; Yoon, B.S.; Lyons, K.M. BMP Canonical Smad Signaling through Smad1 and Smad5 Is Required for Endochondral Bone Formation. Development 2009, 136, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.S.; Ovchinnikov, D.A.; Yoshii, I.; Mishina, Y.; Behringer, R.R.; Lyons, K.M. Bmpr1a and Bmpr1b Have Overlapping Functions and Are Essential for Chondrogenesis in Vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 5062–5067. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.S.; Pogue, R.; Ovchinnikov, D.A.; Yoshii, I.; Mishina, Y.; Behringer, R.R.; Lyons, K.M. BMPs Regulate Multiple Aspects of Growth-Plate Chondrogenesis through Opposing Actions on FGF Pathways. Development 2006, 133, 4667–4678. [Google Scholar] [CrossRef] [PubMed]
- Brunet, L.J.; McMahon, J.A.; McMahon, A.P.; Harland, R.M. Noggin, Cartilage Morphogenesis, and Joint Formation in the Mammalian Skeleton. Science 1998, 280, 1455–1457. [Google Scholar] [CrossRef] [PubMed]
- Joeng, K.S.; Schumacher, C.A.; Zylstra-Diegel, C.R.; Long, F.; Williams, B.O. Lrp5 and Lrp6 Redundantly Control Skeletal Development in the Mouse Embryo. Dev. Biol. 2011, 359, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.E.; Cantley, L.; Yasuaha, R.; Iwamoto, M.; Pacifici, M.; Enomoto-Iwamoto, M. Distribution of Slow-Cycling Cells in Epiphyseal Cartilage and Requirement of β-Catenin Signaling for Their Maintenance in Growth Plate: Slow-Cycling Cells in Epiphysis. J. Orthop. Res. 2014, 32, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Topol, L.; Lee, H.; Wu, J. Wnt5a and Wnt5b Exhibit Distinct Activities in Coordinating Chondrocyte Proliferation and Differentiation. Development 2003, 130, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Colvin, J.S.; Bohne, B.A.; Harding, G.W.; McEwen, D.G.; Ornitz, D.M. Skeletal Overgrowth and Deafness in Mice Lacking Fibroblast Growth Factor Receptor 3. Nat. Genet. 1996, 12, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Wynshaw-Boris, A.; Zhou, F.; Kuo, A.; Leder, P. Fibroblast Growth Factor Receptor 3 Is a Negative Regulator of Bone Growth. Cell 1996, 84, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhang, R.; Chen, H.; Du, X.; Chen, S.; Huang, J.; Liu, M.; Xu, M.; Luo, F.; Jin, M.; et al. Fgfr3 Mutation Disrupts Chondrogenesis and Bone Ossification in Zebrafish Model Mimicking CATSHL Syndrome Partially via Enhanced Wnt/β-Catenin Signaling. Theranostics 2020, 10, 7111–7130. [Google Scholar] [CrossRef] [PubMed]
- Iyer, N.V.; Kotch, L.E.; Agani, F.; Leung, S.W.; Laughner, E.; Wenger, R.H.; Gassmann, M.; Gearhart, J.D.; Lawler, A.M.; Yu, A.Y.; et al. Cellular and Developmental Control of O 2 Homeostasis by Hypoxia-Inducible Factor 1α. Genes Dev. 1998, 12, 149–162. [Google Scholar] [CrossRef]
- Schipani, E.; Ryan, H.E.; Didrickson, S.; Kobayashi, T.; Knight, M.; Johnson, R.S. Hypoxia in Cartilage: HIF-1α Is Essential for Chondrocyte Growth Arrest and Survival. Genes Dev. 2001, 15, 2865–2876. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, W.; Sigaux, J.; Modrowski, D.; Devignes, C.-S.; Funck-Brentano, T.; Richette, P.; Ea, H.-K.; Provot, S.; Cohen-Solal, M.; Haÿ, E. Interaction of HIF1α and β-Catenin Inhibits Matrix Metalloproteinase 13 Expression and Prevents Cartilage Damage in Mice. Proc. Natl. Acad. Sci. USA 2016, 113, 5453–5458. [Google Scholar] [CrossRef]
- Vega, R.B.; Matsuda, K.; Oh, J.; Barbosa, A.C.; Yang, X.; Meadows, E.; McAnally, J.; Pomajzl, C.; Shelton, J.M.; Richardson, J.A.; et al. Histone Deacetylase 4 Controls Chondrocyte Hypertrophy during Skeletogenesis. Cell 2004, 119, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Zhao, X.; Mao, G.; Zhang, Z.; Wen, X.; Zhang, C.; Liao, W.; Zhang, Z. MicroRNA-455-3p Promotes TGF-β Signaling and Inhibits Osteoarthritis Development by Directly Targeting PAK2. Exp. Mol. Med. 2019, 51, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Li, H.; Sun, H.; Zeng, A.; Lin, R.; Zhao, J.; Zhang, Z. MiR-455-3p Reduces Apoptosis and Alleviates Degeneration of Chondrocyte through Regulating PI3K/AKT Pathway. Life Sci. 2020, 253, 117718. [Google Scholar] [CrossRef] [PubMed]
- Miyaki, S.; Sato, T.; Inoue, A.; Otsuki, S.; Ito, Y.; Yokoyama, S.; Kato, Y.; Takemoto, F.; Nakasa, T.; Yamashita, S.; et al. MicroRNA-140 Plays Dual Roles in Both Cartilage Development and Homeostasis. Genes Dev. 2010, 24, 1173–1185. [Google Scholar] [CrossRef]
- Nakamura, Y.; Inloes, J.B.; Katagiri, T.; Kobayashi, T. Chondrocyte-Specific MicroRNA-140 Regulates Endochondral Bone Development and Targets Dnpep To Modulate Bone Morphogenetic Protein Signaling. Mol. Cell. Biol. 2011, 31, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The Control of Chondrogenesis. J. Cell. Biochem. 2006, 97, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.; Wang, G.; Beier, F. Regulation of Chondrocyte Differentiation by the Actin Cytoskeleton and Adhesive Interactions. J. Cell. Physiol. 2007, 213, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Grafe, I.; Alexander, S.; Peterson, J.R.; Snider, T.N.; Levi, B.; Lee, B.; Mishina, Y. TGF-β Family Signaling in Mesenchymal Differentiation. Cold Spring Harb. Perspect. Biol. 2018, 10, a022202. [Google Scholar] [CrossRef] [PubMed]
- Kozhemyakina, E.; Lassar, A.B.; Zelzer, E. A Pathway to Bone: Signaling Molecules and Transcription Factors Involved in Chondrocyte Development and Maturation. Development 2015, 142, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Onyekwelu, I.; Goldring, M.B.; Hidaka, C. Chondrogenesis, Joint Formation, and Articular Cartilage Regeneration. J. Cell. Biochem. 2009, 107, 383–392. [Google Scholar] [CrossRef]
- Chijimatsu, R.; Saito, T. Mechanisms of Synovial Joint and Articular Cartilage Development. Cell. Mol. Life Sci. 2019, 76, 3939–3952. [Google Scholar] [CrossRef]
- Tong, W.; Tower, R.J.; Chen, C.; Wang, L.; Zhong, L.; Wei, Y.; Sun, H.; Cao, G.; Jia, H.; Pacifici, M.; et al. Periarticular Mesenchymal Progenitors Initiate and Contribute to Secondary Ossification Center Formation During Mouse Long Bone Development. Stem Cells 2019, 37, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Rigueur, D.; Lyons, K.M. TGFβ Signaling in Cartilage Development and Maintenance. Birth Defects Res. Part C 2014, 102, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-L.; Yu, B.; Deng, P.; Wang, C.-Y.; Hong, C. Transforming Growth Factor-β-Induced KDM4B Promotes Chondrogenic Differentiation of Human Mesenchymal Stem Cells. Stem Cells 2016, 34, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Fu, P.; Cong, R.; Wu, H.; Pei, M. Strategies to Minimize Hypertrophy in Cartilage Engineering and Regeneration. Genes Dis. 2015, 2, 76–95. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Y.; Wang, A.; Yu, C.; Luo, Y.; Liu, R.; Zhao, Y.; Xiao, J. Effect of CD44 on Differentiation of Human Amniotic Mesenchymal Stem Cells into Chondrocytes via Smad and ERK Signaling Pathways. Mol. Med. Rep. 2020, 21, 2357–2366. [Google Scholar] [CrossRef] [PubMed]
- Almalki, S.G.; Agrawal, D.K. Key Transcription Factors in the Differentiation of Mesenchymal Stem Cells. Differentiation 2016, 92, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Preitschopf, A.; Zwickl, H.; Li, K.; Lubec, G.; Joo, G.; Rosner, M.; Hengstschläger, M.; Mikula, M. Chondrogenic Differentiation of Amniotic Fluid Stem Cells and Their Potential for Regenerative Therapy. Stem Cell Rev. Rep. 2012, 8, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- De Kroon, L.M.G.; Narcisi, R.; Van Den Akker, G.G.H.; Vitters, E.L.; Blaney Davidson, E.N.; Van Osch, G.J.V.M.; Van Der Kraan, P.M. SMAD3 and SMAD4 Have a More Dominant Role than SMAD2 in TGFβ-Induced Chondrogenic Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells. Sci. Rep. 2017, 7, 43164. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Wang, A.-T.; Zhang, Q.-F.; Liu, R.-M.; Xiao, J.-H. RASL11B Gene Enhances Hyaluronic Acid-Mediated Chondrogenic Differentiation in Human Amniotic Mesenchymal Stem Cells via the Activation of Sox9/ERK/Smad Signals. Exp. Biol. Med. 2020, 245, 1708–1721. [Google Scholar] [CrossRef]
- Hellingman, C.A.; Davidson, E.N.B.; Koevoet, W.; Vitters, E.L.; Van Den Berg, W.B.; Van Osch, G.J.V.M.; Van Der Kraan, P.M. Smad Signaling Determines Chondrogenic Differentiation of Bone-Marrow-Derived Mesenchymal Stem Cells: Inhibition of Smad1/5/8P Prevents Terminal Differentiation and Calcification. Tissue Eng. Part A 2011, 17, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.; Yang, T.; Chen, Y.; Munivez, E.; Bertin, T.; Zabel, B.; Lee, B. Interaction of TGFβ and BMP Signaling Pathways during Chondrogenesis. PLoS ONE 2011, 6, e16421. [Google Scholar] [CrossRef]
- Shintani, N.; Siebenrock, K.A.; Hunziker, E.B. TGF-SS1 Enhances the BMP-2-Induced Chondrogenesis of Bovine Synovial Explants and Arrests Downstream Differentiation at an Early Stage of Hypertrophy. PLoS ONE 2013, 8, e53086. [Google Scholar] [CrossRef] [PubMed]
- Bradley, E.W.; Drissi, M.H. WNT5A Regulates Chondrocyte Differentiation through Differential Use of the CaN/NFAT and IKK/NF-κB Pathways. Mol. Endocrinol. 2010, 24, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Letamendia, A.; Labbé, E.; Attisano, L. Transcriptional Regulation by Smads: Crosstalk between the TGF-Beta and Wnt Pathways. J. Bone Jt. Surg. Am. 2001, 83 (Suppl. S1), S31–S39. [Google Scholar] [CrossRef]
- Zhou, S.; Eid, K.; Glowacki, J. Cooperation Between TGF-β and Wnt Pathways During Chondrocyte and Adipocyte Differentiation of Human Marrow Stromal Cells. J. Bone Miner. Res. 2004, 19, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Pfander, D.; Gelse, K. Hypoxia and Osteoarthritis: How Chondrocytes Survive Hypoxic Environments. Curr. Opin. Rheumatol. 2007, 19, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Robins, J.C.; Akeno, N.; Mukherjee, A.; Dalal, R.R.; Aronow, B.J.; Koopman, P.; Clemens, T.L. Hypoxia Induces Chondrocyte-Specific Gene Expression in Mesenchymal Cells in Association with Transcriptional Activation of Sox9. Bone 2005, 37, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Stokes, D.G.; Liu, G.; Coimbra, I.B.; Piera-Velazquez, S.; Crowl, R.M.; Jiménez, S.A. Assessment of the Gene Expression Profile of Differentiated and Dedifferentiated Human Fetal Chondrocytes by Microarray Analysis. Arthritis Rheum. 2002, 46, 404–419. [Google Scholar] [CrossRef] [PubMed]
- Duval, E.; Baugé, C.; Andriamanalijaona, R.; Bénateau, H.; Leclercq, S.; Dutoit, S.; Poulain, L.; Galéra, P.; Boumédiene, K. Molecular Mechanism of Hypoxia-Induced Chondrogenesis and Its Application in in Vivo Cartilage Tissue Engineering. Biomaterials 2012, 33, 6042–6051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vasheghani, F.; Li, Y.; Blati, M.; Simeone, K.; Fahmi, H.; Lussier, B.; Roughley, P.; Lagares, D.; Pelletier, J.-P.; et al. Cartilage-Specific Deletion of mTOR Upregulates Autophagy and Protects Mice from Osteoarthritis. Ann. Rheum. Dis. 2015, 74, 1432–1440. [Google Scholar] [CrossRef]
- Sun, Y.; Qin, B. Long Noncoding RNA MALAT1 Regulates HDAC4-mediated Proliferation and Apoptosis via Decoying of miR-140-5p in Osteosarcoma Cells. Cancer Med. 2018, 7, 4584–4597. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, Q.; Chen, Z.; Shen, B.; Yang, J.; Kang, P.; Zhou, Z.; Pei, F. MicroRNA-140 Suppresses Human Chondrocytes Hypertrophy by Targeting SMAD1 and Controlling the Bone Morphogenetic Protein Pathway in Osteoarthritis. Am. J. Med. Sci. 2018, 355, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Tardif, G.; Hum, D.; Pelletier, J.-P.; Duval, N.; Martel-Pelletier, J. Regulation of the IGFBP-5 and MMP-13 Genes by the microRNAs miR-140 and miR-27a in Human Osteoarthritic Chondrocytes. BMC Musculoskelet. Disord. 2009, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, S.; Miyaki, S.; Kato, Y.; Yokoyama, S.; Sato, T.; Barrionuevo, F.; Akiyama, H.; Scherer, G.; Takada, S.; Asahara, H. L-Sox5 and Sox6 Proteins Enhance Chondrogenic miR-140 MicroRNA Expression by Strengthening Dimeric Sox9 Activity. J. Biol. Chem. 2012, 287, 22206–22215. [Google Scholar] [CrossRef] [PubMed]
- Dudek, K.A.; Lafont, J.E.; Martinez-Sanchez, A.; Murphy, C.L. Type II Collagen Expression Is Regulated by Tissue-Specific miR-675 in Human Articular Chondrocytes. J. Biol. Chem. 2010, 285, 24381–24387. [Google Scholar] [CrossRef]
- Martinez-Sanchez, A.; Dudek, K.A.; Murphy, C.L. Regulation of Human Chondrocyte Function through Direct Inhibition of Cartilage Master Regulator SOX9 by MicroRNA-145 (miRNA-145). J. Biol. Chem. 2012, 287, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Horita, M.; Hsu, S.-N.; Raper, A.; Farquharson, C.; Stephen, L.A. miR-29b Inhibits TGF-Β1-Induced Cell Proliferation in Articular Chondrocytes. Biochem. Biophys. Rep. 2022, 29, 101216. [Google Scholar] [CrossRef] [PubMed]
- Le, L.T.T.; Swingler, T.E.; Crowe, N.; Vincent, T.L.; Barter, M.J.; Donell, S.T.; Delany, A.M.; Dalmay, T.; Young, D.A.; Clark, I.M. The microRNA-29 Family in Cartilage Homeostasis and Osteoarthritis. J. Mol. Med. 2016, 94, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Manuelpillai, U.; Moodley, Y.; Borlongan, C.V.; Parolini, O. Amniotic Membrane and Amniotic Cells: Potential Therapeutic Tools to Combat Tissue Inflammation and Fibrosis? Placenta 2011, 32, S320–S325. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, Y.; Yang, J.; Huang, J.; Luo, C.; Zhang, J.; Yan, W.; Ao, Y. Bone Morphogenetic Protein 9 Enhances Osteogenic and Angiogenic Responses of Human Amniotic Mesenchymal Stem Cells Cocultured with Umbilical Vein Endothelial Cells through the PI3K/AKT/m-TOR Signaling Pathway. Aging 2021, 13, 24829–24849. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Lehmann, T.; Cai, H.; Stolz, D.B.; Strom, S.C. Stem Cell Characteristics of Amniotic Epithelial Cells. Stem Cells 2005, 23, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Bühring, H.-J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise Review: Isolation and Characterization of Cells from Human Term Placenta: Outcome of the First International Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Soncini, M.; Evangelista, M.; Schmidt, D. Amniotic Membrane and Amniotic Fluid-Derived Cells: Potential Tools for Regenerative Medicine? Regen. Med. 2009, 4, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Tamagawa, T.; Ishiwata, I.; Saito, S. Establishment and Characterization of a Pluripotent Stem Cell Line Derived from Human Amniotic Membranes and Initiation of Germ Layers in Vitro. Hum. Cell 2008, 17, 125–130. [Google Scholar] [CrossRef]
- Mattioli, M.; Gloria, A.; Turriani, M.; Mauro, A.; Curini, V.; Russo, V.; Tetè, S.; Marchisio, M.; Pierdomenico, L.; Berardinelli, P.; et al. Stemness Characteristics and Osteogenic Potential of Sheep Amniotic Epithelial Cells. Cell Biol. Int. 2012, 36, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Corradetti, B.; Bizzaro, D.; Magatti, M.; Ressel, L.; Tassan, S.; Parolini, O.; Cremonesi, F. Characterization and Potential Applications of Progenitor-like Cells Isolated from Horse Amniotic Membrane: Characterization and Application of Horse Amnion-Derived Progenitor-like Cells. J. Tissue Eng. Regen. Med. 2012, 6, 622–635. [Google Scholar] [CrossRef]
- Rutigliano, L.; Corradetti, B.; Valentini, L.; Bizzaro, D.; Meucci, A.; Cremonesi, F.; Lange-Consiglio, A. Molecular Characterization and in Vitro Differentiation of Feline Progenitor-like Amniotic Epithelial Cells. Stem Cell Res. Ther. 2013, 4, 133. [Google Scholar] [CrossRef] [PubMed]
- Muttini, A.; Barboni, B.; Valbonetti, L.; Russo, V.; Maffulli, N. Amniotic Epithelial Stem Cells: Salient Features and Possible Therapeutic Role. Sports Med. Arthrosc. Rev. 2018, 26, 70–74. [Google Scholar] [CrossRef]
- Kim, J.; Kang, H.M.; Kim, H.; Kim, M.R.; Kwon, H.C.; Gye, M.C.; Kang, S.G.; Yang, H.S.; You, J. Ex Vivo Characteristics of Human Amniotic Membrane-Derived Stem Cells. Cloning Stem Cells 2007, 9, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Longo, U.; Vaquero, J.; Forriol, F.; Loppini, M.; Khan, W.; Denaro, V. Amniotic Membrane Transplant for Articular Cartilage Repair: An Experimental Study in Sheep. Curr. Stem Cell Res. Ther. 2014, 10, 77–83. [Google Scholar] [CrossRef]
- Veronesi, F.; Berni, M.; Marchiori, G.; Cassiolas, G.; Muttini, A.; Barboni, B.; Martini, L.; Fini, M.; Lopomo, N.F.; Marcacci, M.; et al. Evaluation of Cartilage Biomechanics and Knee Joint Microenvironment after Different Cell-Based Treatments in a Sheep Model of Early Osteoarthritis. Int. Orthop. (SICOT) 2021, 45, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Fini, M.; Martini, L.; Berardinelli, P.; Russo, V.; Filardo, G.; Di Matteo, B.; Marcacci, M.; Kon, E. In Vivo Model of Osteoarthritis to Compare Allogenic Amniotic Epithelial Stem Cells and Autologous Adipose Derived Cells. Biology 2022, 11, 681. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.Z.; Park, S.R.; Choi, B.H.; Lee, K.-Y.; Kang, C.K.; Min, B.-H. Human Amniotic Membrane as a Delivery Matrix for Articular Cartilage Repair. Tissue Eng. 2007, 13, 693–702. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Liu, Z.; Zhang, J.; Shen, M.; Li, Y.; Jin, Y.; Liu, Y. Human Amniotic Mesenchymal Stem Cell Sheets Encapsulating Cartilage Particles Facilitate Repair of Rabbit Osteochondral Defects. Am. J. Sports Med. 2020, 48, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Shim, M.-S.; Shim, S.H.; Yang, H.N.; Jeon, S.Y.; Woo, D.G.; Lee, D.R.; Yoon, T.K.; Park, K.-H. Chondrogenic Potential of Stem Cells Derived from Amniotic Fluid, Adipose Tissue, or Bone Marrow Encapsulated in Fibrin Gels Containing TGF-Β3. Biomaterials 2011, 32, 8139–8149. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yu, G.; Cao, C.; Pang, J.; Chen, X. Bone Morphogenetic Protein-7 Promotes Chondrogenesis in Human Amniotic Epithelial Cells. Int. Orthop. (SICOT) 2011, 35, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Nogami, M.; Tsuno, H.; Koike, C.; Okabe, M.; Yoshida, T.; Seki, S.; Matsui, Y.; Kimura, T.; Nikaido, T. Isolation and Characterization of Human Amniotic Mesenchymal Stem Cells and Their Chondrogenic Differentiation. Transplantation 2012, 93, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Saghati, S.; Rahbarghazi, R.; Baradar Khoshfetrat, A.; Moharamzadeh, K.; Tayefi Nasrabadi, H.; Roshangar, L. Phenolated Alginate-Collagen Hydrogel Induced Chondrogenic Capacity of Human Amniotic Mesenchymal Stem Cells. J. Biomater. Appl. 2021, 36, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Ng, F.; Boucher, S.; Koh, S.; Sastry, K.S.R.; Chase, L.; Lakshmipathy, U.; Choong, C.; Yang, Z.; Vemuri, M.C.; Rao, M.S.; et al. PDGF, TGF-β, and FGF Signaling Is Important for Differentiation and Growth of Mesenchymal Stem Cells (MSCs): Transcriptional Profiling Can Identify Markers and Signaling Pathways Important in Differentiation of MSCs into Adipogenic, Chondrogenic, and Osteogenic Lineages. Blood 2008, 112, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Boeuf, S.; Richter, W. Chondrogenesis of Mesenchymal Stem Cells: Role of Tissue Source and Inducing Factors. Stem Cell Res. Ther. 2010, 1, 31. [Google Scholar] [CrossRef] [PubMed]
- Topoluk, N.; Hawkins, R.; Tokish, J.; Mercuri, J. Amniotic Mesenchymal Stromal Cells Exhibit Preferential Osteogenic and Chondrogenic Differentiation and Enhanced Matrix Production Compared with Adipose Mesenchymal Stromal Cells. Am. J. Sports Med. 2017, 45, 2637–2646. [Google Scholar] [CrossRef] [PubMed]
- Mauro, A.; Sanyal, H.; Canciello, A.; Berardinelli, P.; Russo, V.; Bernabò, N.; Valbonetti, L.; Barboni, B. In Vitro Effect of Estradiol and Progesterone on Ovine Amniotic Epithelial Cells. Stem Cells Int. 2019, 2019, 8034578. [Google Scholar] [CrossRef]
- Wei, J.P.; Nawata, M.; Wakitani, S.; Kametani, K.; Ota, M.; Toda, A.; Konishi, I.; Ebara, S.; Nikaido, T. Human Amniotic Mesenchymal Cells Differentiate into Chondrocytes. Cloning Stem Cells 2009, 11, 19–26. [Google Scholar] [CrossRef]
- Wang, Y.-Q.; Wang, N.-X.; Luo, Y.; Yu, C.-Y.; Xiao, J.-H. Ganoderal A Effectively Induces Osteogenic Differentiation of Human Amniotic Mesenchymal Stem Cells via Cross-Talk between Wnt/β-Catenin and BMP/SMAD Signaling Pathways. Biomed. Pharmacother. 2020, 123, 109807. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, A.-T.; Xiao, J.-H. CD44 Mediates Hyaluronan to Promote the Differentiation of Human Amniotic Mesenchymal Stem Cells into Chondrocytes. Biotechnol. Lett. 2023, 45, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Gunti, S.; Hoke, A.T.K.; Vu, K.P.; London, N.R. Organoid and Spheroid Tumor Models: Techniques and Applications. Cancers 2021, 13, 874. [Google Scholar] [CrossRef] [PubMed]
- Winter, A.; Breit, S.; Parsch, D.; Benz, K.; Steck, E.; Hauner, H.; Weber, R.M.; Ewerbeck, V.; Richter, W. Cartilage-like Gene Expression in Differentiated Human Stem Cell Spheroids: A Comparison of Bone Marrow-Derived and Adipose Tissue-Derived Stromal Cells. Arthritis Rheum. 2003, 48, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Yoshida, T.; Okabe, M.; Toda, A.; Higuchi, O.; Nogami, M.; Yoneda, N.; Zhou, K.; Kyo, S.; Kiyono, T.; et al. Establishment of Immortalized Human Amniotic Mesenchymal Stem Cells. Cell Transplant. 2013, 22, 267–278. [Google Scholar] [CrossRef]
- Muiños-López, E.; Hermida-Gómez, T.; Fuentes-Boquete, I.; De Toro-Santos, J.; Blanco, F.J.; Díaz-Prado, S.M. Human Amniotic Mesenchymal Stromal Cells as Favorable Source for Cartilage Repair. Tissue Eng. Part A 2017, 23, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.R.; Reis, R.L.; Oliveira, J.M. Layered Scaffolds for Osteochondral Tissue Engineering. In Osteochondral Tissue Engineering; Oliveira, J.M., Pina, S., Reis, R.L., San Roman, J., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2018; Volume 1058, pp. 193–218. ISBN 978-3-319-76710-9. [Google Scholar]
- Kolambkar, Y.M.; Peister, A.; Soker, S.; Atala, A.; Guldberg, R.E. Chondrogenic Differentiation of Amniotic Fluid-Derived Stem Cells. J. Mol. Hist. 2007, 38, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Condello, V.; Filardo, G.; Madonna, V.; Andriolo, L.; Screpis, D.; Bonomo, M.; Zappia, M.; Dei Giudici, L.; Zorzi, C. Use of a Biomimetic Scaffold for the Treatment of Osteochondral Lesions in Early Osteoarthritis. BioMed Res. Int. 2018, 2018, 7937089. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, Z.; Wu, H.; Li, W.; Zheng, L.; Zhao, J. Collagen-Alginate as Bioink for Three-Dimensional (3D) Cell Printing Based Cartilage Tissue Engineering. Mater. Sci. Eng. C 2018, 83, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Naseer, N.; Bashir, S.; Latief, N.; Latif, F.; Khan, S.N.; Riazuddin, S. Human Amniotic Membrane as Differentiating Matrix for in Vitro Chondrogenesis. Regen. Med. 2018, 13, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Lu, M.; Wang, T.; Dian, D.; Zhong, Y.; Aleahmad, M. Human Amniotic Membrane as a Delivery Vehicle for Stem Cell-Based Therapies. Life Sci. 2021, 272, 119157. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Prado, S.; Rendal-Vázquez, M.E.; Muiños-López, E.; Hermida-Gómez, T.; Rodríguez-Cabarcos, M.; Fuentes-Boquete, I.; De Toro, F.J.; Blanco, F.J. Potential Use of the Human Amniotic Membrane as a Scaffold in Human Articular Cartilage Repair. Cell Tissue Bank. 2010, 11, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.-L.; Sulaiman, S.; Pingguan-Murphy, B.; Selvaratnam, L.; Tai, C.-C.; Kamarul, T. Human Amnion as a Novel Cell Delivery Vehicle for Chondrogenic Mesenchymal Stem Cells. Cell Tissue Bank. 2011, 12, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Lindenmair, A.; Nürnberger, S.; Stadler, G.; Meinl, A.; Hackl, C.; Eibl, J.; Gabriel, C.; Hennerbichler, S.; Redl, H.; Wolbank, S. Intact Human Amniotic Membrane Differentiated towards the Chondrogenic Lineage. Cell Tissue Bank. 2014, 15, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Rastegar Adib, F.; Bagheri, F.; Sharifi, A.M. Osteochondral Regeneration in Rabbit Using Xenograft Decellularized ECM in Combination with Different Biological Products; Platelet-rich Fibrin, Amniotic Membrane Extract, and Mesenchymal Stromal Cells. J. Biomed. Mater. Res. 2022, 110, 2089–2099. [Google Scholar] [CrossRef] [PubMed]
- Abraham, D.M.; Herman, C.; Witek, L.; Cronstein, B.N.; Flores, R.L.; Coelho, P.G. Self-assembling Human Skeletal Organoids for Disease Modeling and Drug Testing. J. Biomed. Mater. Res. 2022, 110, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Hemeryck, L.; Hermans, F.; Chappell, J.; Kobayashi, H.; Lambrechts, D.; Lambrichts, I.; Bronckaers, A.; Vankelecom, H. Organoids from Human Tooth Showing Epithelial Stemness Phenotype and Differentiation Potential. Cell. Mol. Life Sci. 2022, 79, 153. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.A.; Shang, J.; Xiang, S.; Li, E.N.; Yagi, H.; Riewruja, K.; Lin, H.; Tuan, R.S. Articular Tissue-Mimicking Organoids Derived from Mesenchymal Stem Cells and Induced Pluripotent Stem Cells. Organoids 2022, 1, 135–148. [Google Scholar] [CrossRef]
- Crispim, J.F.; Ito, K. De Novo Neo-Hyaline-Cartilage from Bovine Organoids in Viscoelastic Hydrogels. Acta Biomater. 2021, 128, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Clavell, R.; Martín De Llano, J.J.; Milián, L.; Oliver, M.; Mata, M.; Carda, C.; Sancho-Tello, M. Chondrogenic Potential of Human Dental Pulp Stem Cells Cultured as Microtissues. Stem Cells Int. 2021, 2021, 7843798. [Google Scholar] [CrossRef]
- O’Connor, S.K.; Katz, D.B.; Oswald, S.J.; Groneck, L.; Guilak, F. Formation of Osteochondral Organoids from Murine Induced Pluripotent Stem Cells. Tissue Eng. Part A 2021, 27, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.S.; Mithoefer, K. Scientific Developments and Clinical Applications Utilizing Chondrons and Chondrocytes with Matrix for Cartilage Repair. Cartilage 2021, 13, 1195S–1205S. [Google Scholar] [CrossRef] [PubMed]
- Jessop, Z.M.; Al-Sabah, A.; Francis, W.R.; Whitaker, I.S. Transforming Healthcare through Regenerative Medicine. BMC Med. 2016, 14, 115. [Google Scholar] [CrossRef] [PubMed]
- Badylak, S.; Rosenthal, N. Regenerative Medicine: Are We There Yet? NPJ Regen. Med. 2017, 2, 2. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
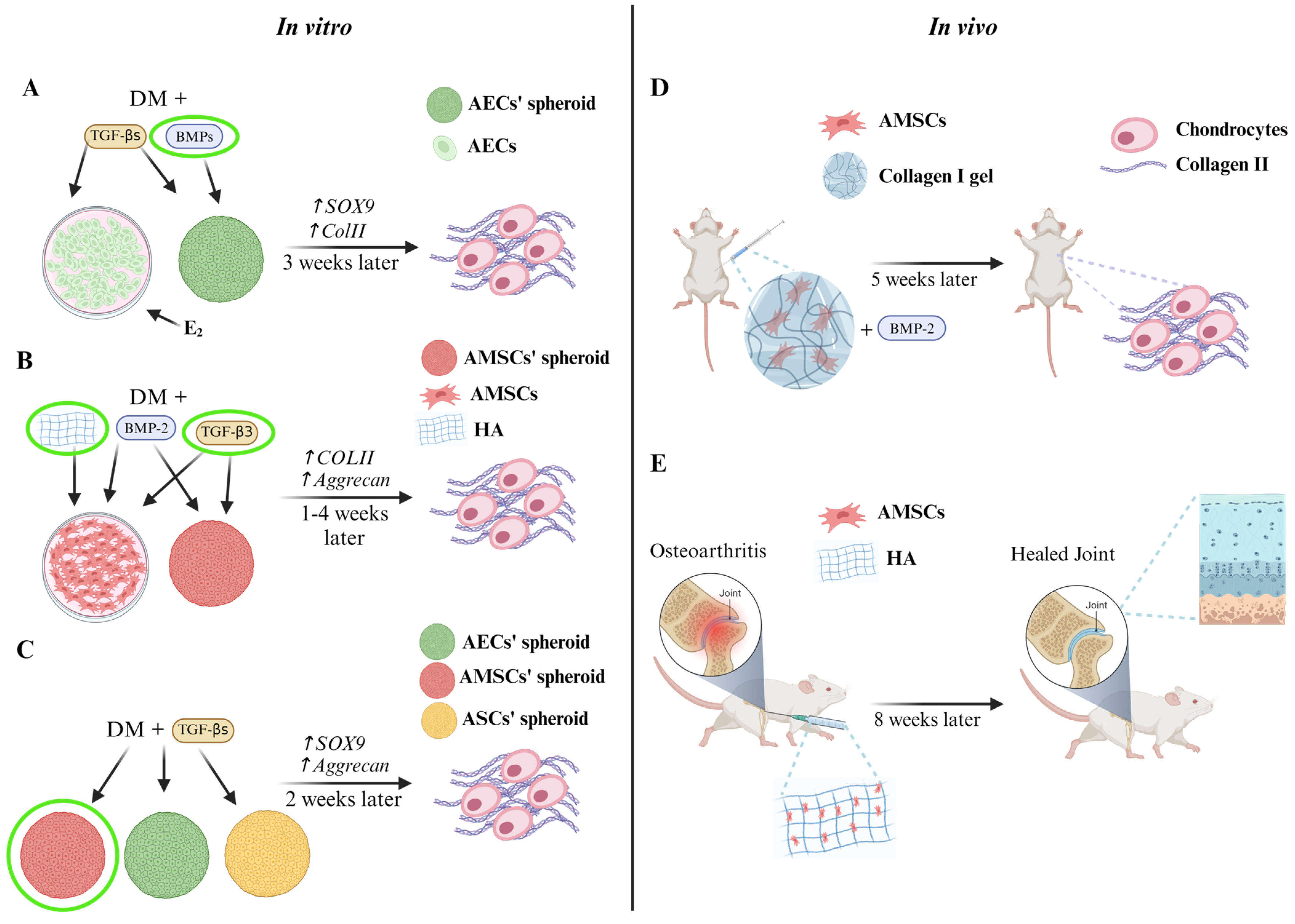{kind=link}
| KO Models | Target | Effects | References |
|---|---|---|---|
| mEmbryo chimeras from SOX9-/- embryonic stem cells | SOX9 |
| Bi et al., 1999 [30] |
| mSOX5-/- and SOX6-/- | SOX5 and SOX6 (downstream to SOX9) |
| Smits et al., 2001 [31] |
| mConditionally SOX9-/- | SOX trio genes | Before condensation:
| Akiyama et al., 2002 [32] |
| mSMAD3(ex8/ex8) KO | TGF-β/SMAD3 signalling pathway | Alterations during late phases of chondrogenesis:
| Yang et al., 2001 [33] |
| mSMAD3-/- | TGF-β/SMAD3 signalling pathway |
| Li et al., 2005 [34] Kawakatsu et al., 2011 [35] |
| mMT-DNIIR (KO for TGFRII) | TGF-β signalling pathway |
| Serra et al., 1997 [36] |
| mBMP2-/- and BMP4-/- embryos | BMP signalling pathway |
| Winnier et al., 1995 [37] Zhang et Bradley. 1996 [38] |
| mConditionally BMP2-/- and BMP4-/- or BMP2-/- and BMP7-/- | BMP signalling pathway |
| Bandyopadhyay et al., 2006 [39] |
| mSMAD1-/- and SMAD5-/- | BMP/SMAD1/SMAD5 canonical signalling pathway |
| Retting et al., 2009 [40] |
| mBMPR1B-/- and conditionally BMPR1A-/- | BMP signalling pathway | KO for BMPR1B:
| Yoon et al., 2005 [41] Yoon et al., 2006 [42] |
| mNoggin-/- | Noggin-mediated BMP pathway antagonism |
| Brunet et al., 1998 [43] |
| m-β-catenin-/-, LRP5-/-, and LRP6-/- mice | Wnt/β-catenin signalling pathway (canonical pathway) |
| Joeng et al., 2011 [44] |
| m-β-catenin-/- | Wnt/β-catenin signalling pathway (canonical pathway) |
| Candela et al., 2014 [45] |
| mWnt5a-/- | Non-canonical Wnt signalling pathway |
| Yang et al., 2003 [46] |
| mFGFR3-/- | FGF signalling pathway |
| Colvin et al., 1996 [47] Deng et al., 1996 [48] |
| zebrafish FGFR3-/- | FGF signalling pathway |
| Sun et al., 2020 [49] |
| mHIF1A-/- | Hypoxia response pathway |
| Iyer et al., 1998 [50] |
| mConditionally HIF1A-/- | Hypoxia response pathway |
| Schipani et al., 2001 [51] Bouaziz et al., 2016 [52] |
| mHDAC4-/- | Epigenetic modifications |
| Vega et al., 2004 [53] |
| m miR-455-3p-/- | Epigenetic modifications |
| Hu et al., 2019 [54] Wen et al., 2020 [55] |
| m miR-140-/- | Epigenetic modifications |
| Miyaki et al., 2010 [56] Nakamura et al., 2011 [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulcanese, L.; Prencipe, G.; Canciello, A.; Cerveró-Varona, A.; Perugini, M.; Mauro, A.; Russo, V.; Barboni, B. Stem-Cell-Driven Chondrogenesis: Perspectives on Amnion-Derived Cells. Cells 2024, 13, 744. https://doi.org/10.3390/cells13090744
Sulcanese L, Prencipe G, Canciello A, Cerveró-Varona A, Perugini M, Mauro A, Russo V, Barboni B. Stem-Cell-Driven Chondrogenesis: Perspectives on Amnion-Derived Cells. Cells. 2024; 13(9):744. https://doi.org/10.3390/cells13090744
Chicago/Turabian StyleSulcanese, Ludovica, Giuseppe Prencipe, Angelo Canciello, Adrián Cerveró-Varona, Monia Perugini, Annunziata Mauro, Valentina Russo, and Barbara Barboni. 2024. "Stem-Cell-Driven Chondrogenesis: Perspectives on Amnion-Derived Cells" Cells 13, no. 9: 744. https://doi.org/10.3390/cells13090744