Exploiting the Autofluorescent Properties of Photosynthetic Pigments for Analysis of Pigmentation and Morphology in Live Fremyella diplosiphon Cells

Abstract
:1. Introduction
2. Experimental Section
2.1. Strains and culture growth conditions
2.2. Slide preparation
2.3. Confocal imaging parameters
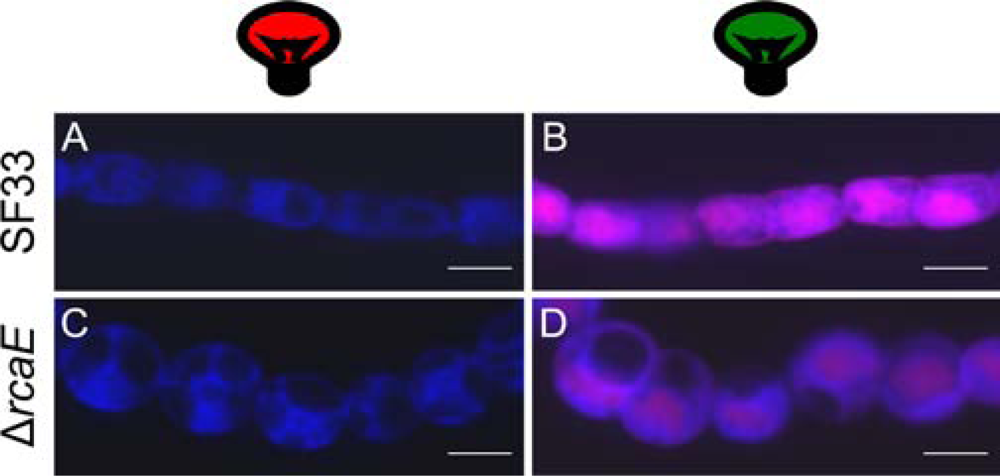
3. Results and Discussion
3.1. Determining imaging parameters for F. diplosiphon PBP autofluorescence
3.2. Imaging different F. diplosiphon strains with optimal settings for detecting PBP autofluorescence
4. Conclusions
Acknowledgments
References
- Bennett, A; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol 1973, 58, 419–435. [Google Scholar]
- Bordowitz, JR; Montgomery, BL. Photoregulation of cellular morphology during complementary chromatic adaptation requires sensor-kinase-class protein RcaE in Fremyella diplosiphon. J. Bacteriol 2008, 190, 4069–4074. [Google Scholar]
- Kehoe, DM; Grossman, AR. Similarity of a chromatic adaptation sensor to phytochrome and ethylene receptors. Science 1996, 273, 1409–1412. [Google Scholar]
- Terauchi, K; Montgomery, BL; Grossman, AR; Lagarias, JC; Kehoe, DM. RcaE is a complementary chromatic adaptation photoreceptor required for green and red light responsiveness. Mol. Microbiol 2004, 51, 567–577. [Google Scholar]
- Grossman, AR. A molecular understanding of complementary chromatic adaptation. Photosynth. Res 2003, 76, 207–215. [Google Scholar]
- Tandeau de Marsac, N; Houmard, J. Adaptation of cyanobacteria to environmental stimuli: New steps towards molecular mechanisms. FEMS Microbiol. Lett 1993, 104, 119–189. [Google Scholar]
- Wolf, E; Schüßler, A. Phycobiliprotein fluorescence of Nostoc punctiforme changes during the life cycle and chromatic adaptation: Characterization by spectral confocal laser scanning microscopy and spectral unmixing. Plant Cell Environ 2005, 28, 480–491. [Google Scholar]
- Hernández Mariné, M; Clavero, E; Roldán, M. Microscopy methods applied to research on cyanobacteria. Limnetica 2004, 23, 179–186. [Google Scholar]
- Mavrakis, M; Pourquié, O; Lecuit, T. Lighting up developmental mechanisms: How fluorescence imaging heralded a new era. Development 2010, 137, 373–387. [Google Scholar]
- Lewis, PJ. Bacterial subcellular architecture: recent advances and future prospects. Mol. Microbiol 2004, 54, 1135–1150. [Google Scholar]
- Pattanaik, B; Montgomery, BL. FdTonB is involved in the photoregulation of cellular morphology during complementary chromatic adaptation in Fremyella diplosiphon. Microbiol 2010, 156, 731–741. [Google Scholar]
- Vermaas, WFJ; Timlin, JA; Jones, HDT; Sinclair, MB; Nieman, LT; Hamad, SW; Melgaard, DK; Haaland, DM. In vivo hyperspectral confocal fluorescence imaging to determine pigment localization and distribution in cyanobacterial cells. Proc. Natl. Acad. Sci. USA 2008, 105, 4050–4055. [Google Scholar]
- Sinha, RP; Richter, P; Faddoul, J; Braun, M; Häder, DP. Effects of UV and visible light on cyanobacteria at the cellular level. Photochem. Photobiol. Sci 2002, 1, 553–559. [Google Scholar]
- Orcutt, K; Ren, S; Gundersen, K. Detecting proteins in highly autofluorescent cells using quantum dot antibody conjugates. Sensors 2009, 9, 7540–7549. [Google Scholar]
- Neunuebel, MR; Golden, JW. The Anabaena sp. strain PCC 7120 gene all2874 encodes a diguanylate cyclase and is required for normal heterocyst development under high-light growth conditions. J. Bacteriol 2008, 190, 6829–6836. [Google Scholar]
- Zhang, W; Du, Y; Khudyakov, I; Fan, Q; Gao, H; Ning, D; Wolk, CP; Xu, X. A gene cluster that regulates both heterocyst differentiation and pattern formation in Anabaena sp. strain PCC 7120. Mol. Microbiol 2007, 66, 1429–1443. [Google Scholar]
- Merino-Puerto, V; Mariscal, V; Mullineaux, CW; Herrero, A; Flores, E. Fra proteins influencing filament integrity, diazotrophy and localization of septal protein SepJ in the heterocyst-forming cyanobacterium Anabaena sp. Mol. Microbiol 2010, 75, 1159–1170. [Google Scholar]
- Rosinski, J; Hainfeld, JF; Rigbi, M; Siegelman, HW. Phycobilisome ultrastructure and chromatic adaptation in Fremyella diplosiphon. Ann. Bot 1981, 47, 1–12. [Google Scholar]
- Bordowitz, JR; Whitaker, MJ; Montgomery, BL. Independence and interdependence of the photoregulation of pigmentation and development inFremyella diplosiphon. Commun Integr Biol 2010, 3. in press. [Google Scholar]
- Whitaker, MJ; Bordowitz, JR; Montgomery, BL. CpcF-dependent regulation of pigmentation and development in Fremyella diplosiphon. Biochem. Biophys. Res. Commun 2009, 389, 602–606. [Google Scholar]
- Cobley, JG; Zerweck, E; Reyes, R; Mody, A; Seludo-Unson, JR; Jaeger, H; Weerasuriya, S; Navankasattusas, S. Construction of shuttle plasmids which can be efficiently mobilized from Escherichia coli into the chromatically adapting cyanobacterium, Fremyella diplosiphon. Plasmid 1993, 30, 90–105. [Google Scholar]
- Reize, IB; Melkonian, M. A new way to investigate living flagellated/ciliated cells in the light microscope: Immobilization of cells in agarose. Bot. Acta 1989, 102, 145–151. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Excitation laser (nm) | Scan range (nm) | λmax RL (nm) | λmax GL (nm) |
|---|---|---|---|
| 488 | ∼500–750 | ∼670 | ∼580 & 650 |
| 543 | ∼550–750 | ∼660 | ∼580 & 660 |
| 633 | ∼640–750 | ∼660–670 | ∼660–670 |
© 2010 by the authors licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bordowitz, J.R.; Montgomery, B.L. Exploiting the Autofluorescent Properties of Photosynthetic Pigments for Analysis of Pigmentation and Morphology in Live Fremyella diplosiphon Cells. Sensors 2010, 10, 6969-6979. https://doi.org/10.3390/s100706969
Bordowitz JR, Montgomery BL. Exploiting the Autofluorescent Properties of Photosynthetic Pigments for Analysis of Pigmentation and Morphology in Live Fremyella diplosiphon Cells. Sensors. 2010; 10(7):6969-6979. https://doi.org/10.3390/s100706969
Chicago/Turabian StyleBordowitz, Juliana R., and Beronda L. Montgomery. 2010. "Exploiting the Autofluorescent Properties of Photosynthetic Pigments for Analysis of Pigmentation and Morphology in Live Fremyella diplosiphon Cells" Sensors 10, no. 7: 6969-6979. https://doi.org/10.3390/s100706969
APA StyleBordowitz, J. R., & Montgomery, B. L. (2010). Exploiting the Autofluorescent Properties of Photosynthetic Pigments for Analysis of Pigmentation and Morphology in Live Fremyella diplosiphon Cells. Sensors, 10(7), 6969-6979. https://doi.org/10.3390/s100706969