
Impedance of the Grape Berry Cuticle as a Novel Phenotypic Trait to Estimate Resistance to Botrytis Cinerea

Abstract
:1. Introduction



2. Experimental Section
2.1. Plant Material and Sampling
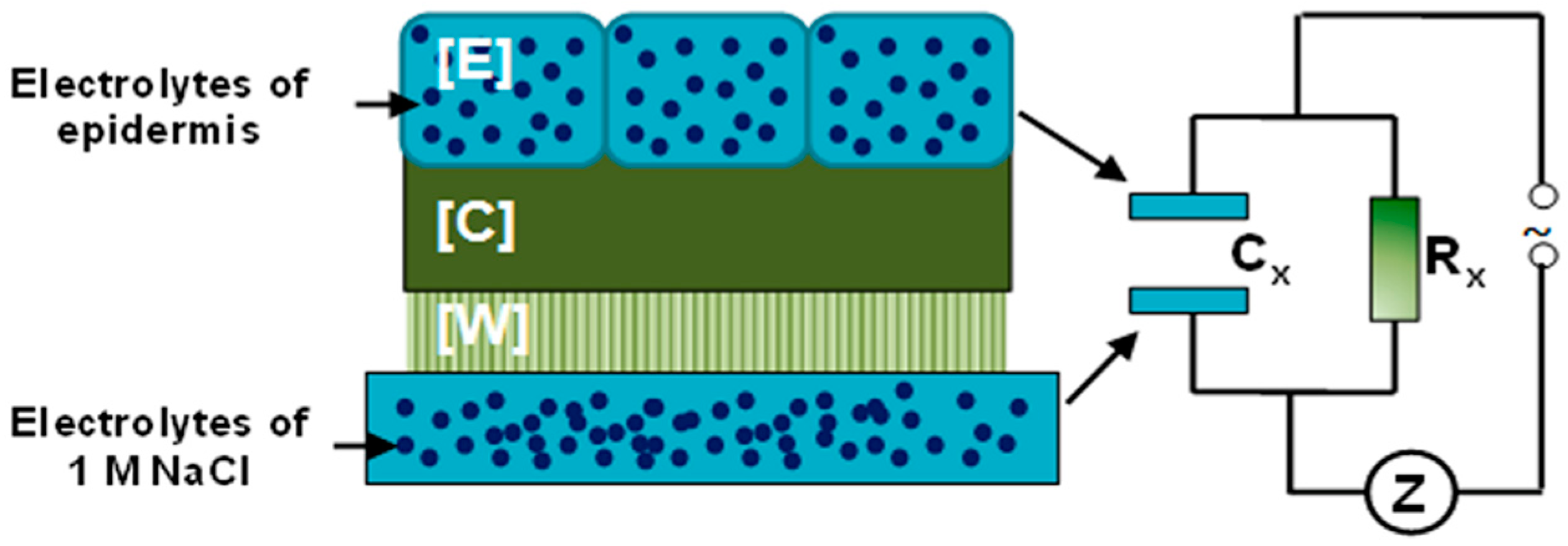
2.2. Construction of the I-Sensor

2.3. Impedance Measurements
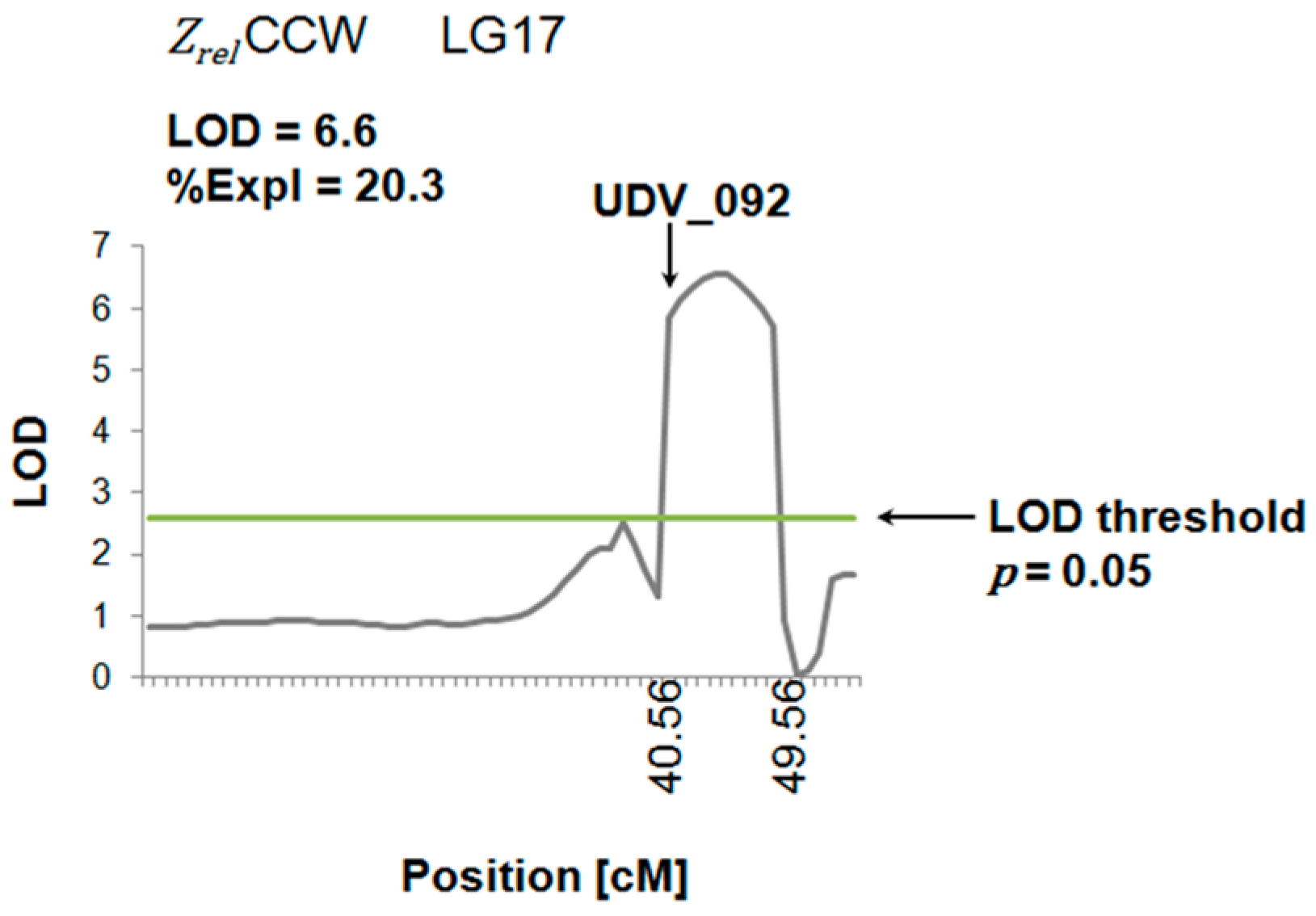
2.4. QTL Analysis
2.5. Reference Evaluations

2.6. Statistical Analysis
3. Results and Discussion
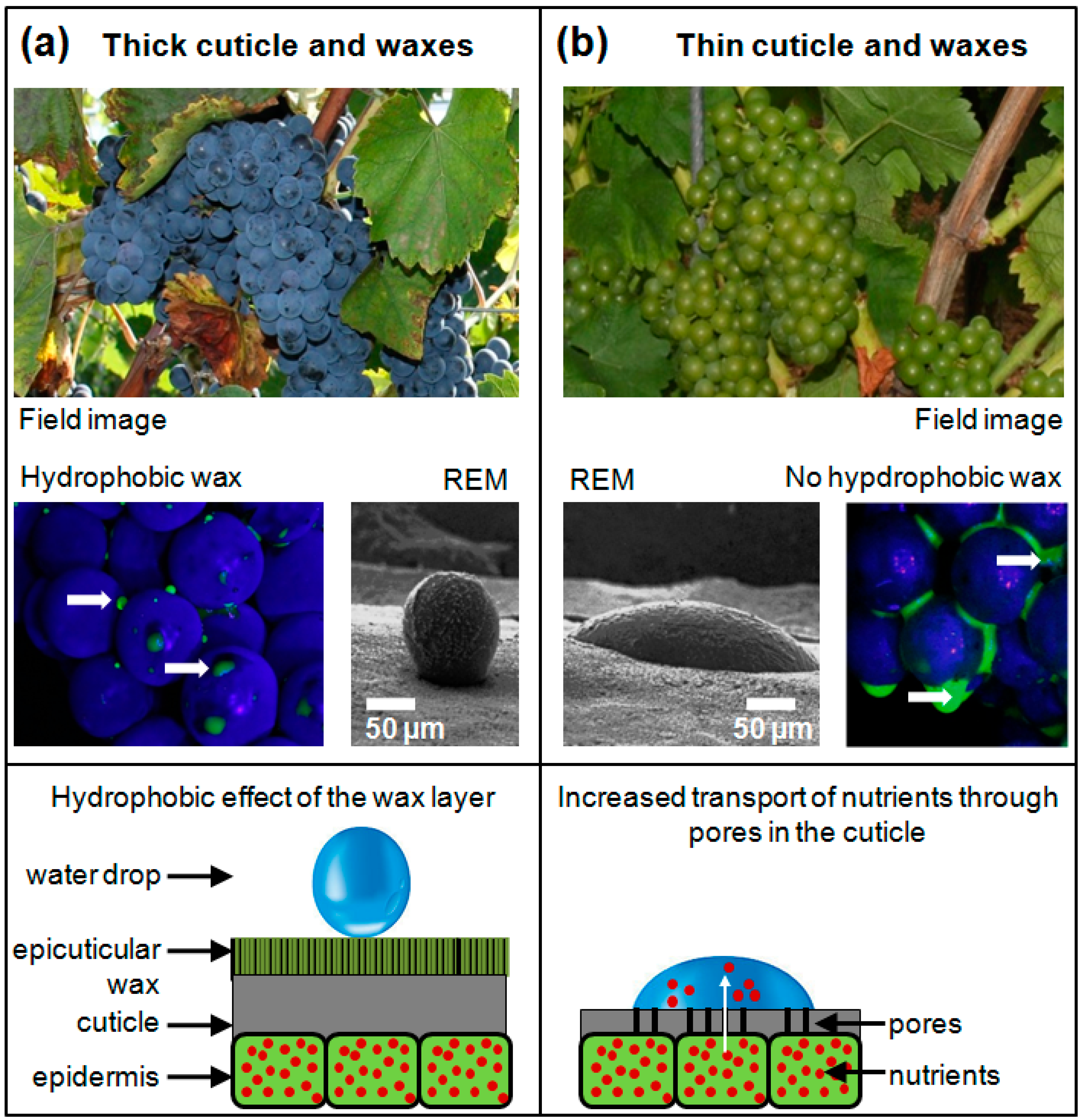
3.1. Validation the Functionality of the I-Sensor
3.2. Novel Phenotypic Trait as Indicator for Resistance of Grapevines to B. cinerea

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zrel | Architecture | Remark | Susceptibility to B. cinerea | Significance |
|---|---|---|---|---|
| CW | Zrel of CW | −0.67 | <0.0001 | |
| C | Zrel C | −0.60 | <0.0001 | |
| W | Zrel of CW - Zrel of C | −0.53 | 0.0004 | |
| Bunch compactness | 0.40 | 0.0096 |
| Bunch Compactness | N | Zrel | Susceptibility to B. cinerea | Significance | Rating of Correlation |
|---|---|---|---|---|---|
| loose | 10 | CW | −0.43 | n.s. | Low |
| C | −0.22 | n.s. | Low | ||
| W | −0.50 | n.s. | Low | ||
| medium | 17 | CW | −0.72 | 0.001 | High |
| C | −0.57 | 0.0173 | Medium | ||
| W | −0.61 | 0.01 | Medium | ||
| compact | 21 | CW | −0.80 | <0.0001 | High |
| C | −0.83 | <0.0001 | High | ||
| W | −0.62 | 0.0027 | Medium |



3.3. Impedance of the Berry Cuticle for QTL Analysis Applications

4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jarvis, W.R. Botryotinia and Botrytis Species: Taxonomy, Physiology, and Pathogenicity; Canadian Department of Agriculture: Ottawa, ON, Canada, 1977; p. 195.
- Molitor, D.; Behr, M.; Fischer, S.; Hoffmann, L.; Evers, D. Timing of cluster-zone leaf removal and its impact on canopy morphology, cluster structure and bunch rot susceptibility of grapes. J. Int. Sci. Vigne Vin. 2011, 45, 149–159. [Google Scholar]
- Broome, J.; English, J.; Marois, J.; Latorre, B.; Aviles, J. Development of an infection model for botrytis bunch rot of grapes based on wetness duration and temperature. Phytopathology 1995, 85, 97–102. [Google Scholar] [CrossRef]
- Ellison, P.; Ash, G.; McDonald, C. An expert system for the management of botrytis cinerea in australian vineyards. I. Development. Agric. Syst. 1998, 56, 185–207. [Google Scholar] [CrossRef]
- Hed, B.; Ngugi, H.K.; Travis, J.W. Relationship between cluster compactness and bunch rot in vignoles grapes. Plant Dis. 2009, 93, 1195–1201. [Google Scholar] [CrossRef]
- Vail, M.; Marois, J. Grape cluster architecture and the susceptibility of berries to botrytis cinerea. Phytopathology 1991, 81, 188–191. [Google Scholar] [CrossRef]
- Vail, M.; Wolpert, J.; Gubler, W.; Rademacher, M. Effect of cluster tightness on botrytis bunch rot in six chardonnay clones. Plant Dis. 1998, 82, 107–109. [Google Scholar] [CrossRef]
- Molitor, D.; Behr, M.; Hoffmann, L.; Evers, D. Impact of grape cluster division on cluster morphology and bunch rot epidemic. Am. J. Enol. Vitic. 2012, 63, 508–514. [Google Scholar] [CrossRef]
- Nair, N.G.; Allen, R.N. Infection of grape flowers and berries by botrytis cinerea as a function of time and temperature. Mycol. Res. 1993, 97, 1012–1014. [Google Scholar] [CrossRef]
- Deytieux-Belleau, C.; Geny, L.; Roudet, J.; Mayet, V.; Donèche, B.; Fermaud, M. Grape berry skin features related to ontogenic resistance to botrytis cinerea. Eur. J. Plant Pathol. 2009, 125, 551–563. [Google Scholar] [CrossRef]
- Gabler, F.M.; Smilanick, J.L.; Mansour, M.; Ramming, D.W.; Mackey, B.E. Correlations of morphological, anatomical, and chemical features of grape berries with resistance to botrytis cinerea. Phytopathology 2003, 93, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, M.; Kassemeyer, H.H.; Hahn, M. Age-dependent grey mould susceptibility and tissue-specific defence gene activation of grapevine berry skins after infection by botrytis cinerea. J. Phytopathol. 2007, 155, 258–263. [Google Scholar] [CrossRef]
- Nanni, V.; Schumacher, J.; Giacomelli, L.; Brazzale, D.; Sbolci, L.; Moser, C.; Tudzynski, P.; Baraldi, E. Vvamp2, a grapevine flower-specific defensin capable of inhibiting botrytis cinerea growth: Insights into its mode of action. Plant Pathol. 2014, 63, 899–910. [Google Scholar] [CrossRef]
- Commenil, P.; Brunet, L.; Audran, J.-C. The development of the grape berry cuticle in relation to susceptibility to bunch rot disease. J. Exp. Bot. 1997, 48, 1599–1607. [Google Scholar] [CrossRef]
- Becker, T.; Knoche, M. Deposition, strain, and microcracking of the cuticle in developing ‘riesling’ grape berries. Vitis 2012, 51, 1–6. [Google Scholar]
- Becker, T.; Knoche, M. Water induces microcracks in the grape berry cuticle. Vitis 2012, 51, 141–142. [Google Scholar]
- Schreiber, L. Transport barriers made of cutin, suberin and associated waxes. Trends Plant Sci. 2010, 15, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, E.; Heredia-Guerrero, J.A.; Heredia, A. The biophysical design of plant cuticles: An overview. New Phytol. 2011, 189, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Benavente, J.; Ramos-Barrado, J.R.; Heredia, A. A study of the electrical behaviour of isolated tomato cuticular membranes and cutin by impedance spectroscopy measurements. Colloids Surf. A Physicochem. Eng. Asp. 1998, 140, 333–338. [Google Scholar] [CrossRef]
- Schreiber, L. Effect of temperature on cuticular transpiration of isolated cuticular membranes and leaf discs. J. Exp. Bot. 2001, 52, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, L.; Skrabs, M.; Hartmann, K.; Diamantopoulos, P.; Simanova, E.; Santrucek, J. Effect of humidity on cuticular water permeability of isolated cuticular membranes and leaf disks. Planta 2001, 214, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Zyprian, E.; Eibach, R.; Töpfer, R. Eine neue genetische karte der weinrebe aus der kreuzung ‘‘GF.GA-47–42’’ x ‘‘villard blanc’’. Deutsches Weinbau Jahrbuch. 2006, 57, 151–158. [Google Scholar]
- Fechter, I.; Hausmann, L.; Zyprian, E.; Daum, M.; Holtgräwe, D.; Weisshaar, B.; Töpfer, R. Qtl analysis of flowering time and ripening traits suggests an impact of a genomic region on linkage group 1 in vitis. Theor. Appl. Genet. 2014, 127, 1857–1872. [Google Scholar] [CrossRef] [PubMed]
- Zyprian, E.; Ochßner, I.; Schwander, F.; Simon, S.; Bonow-Rex, M.; Moreno-Sanz, P.; Grando, M.S.; Wiedemann-Merdinoglu, S.; Merdinoglu, D.; Eibach, R.; et al. Quantitative trait loci affecting resistance traits and ripening of grapevines in a genetic map based on single nucleotide polymorphisms and microsatellites. manuscript in preparation.
- Lorenz, D.H.; Eichhorn, K.W.; Bleiholder, H.; Klose, R.; Meier, U.; Weber, E. Growth stages of the grapevine: Phenological growth stages of the grapevine (vitis vinifera l. Ssp. Vinifera)-codes and descriptions according to the extended bbch scale. Aust. J. Grape Wine Res. 1995, 1, 100–103. [Google Scholar] [CrossRef]
- OIV. OIV publications: OIV descriptor list for grape varieties and Vitis species (2nd ed.). Available online: http://www.Oiv.int (accessed on 26 May 2015).
- Bühl, A. Spss 16: Einführung in die Moderne Datenanalyse; Pearson Deutschland GmbH: Hallbergmoos, Germany, 2008; Volume 7332. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herzog, K.; Wind, R.; Töpfer, R. Impedance of the Grape Berry Cuticle as a Novel Phenotypic Trait to Estimate Resistance to Botrytis Cinerea. Sensors 2015, 15, 12498-12512. https://doi.org/10.3390/s150612498
Herzog K, Wind R, Töpfer R. Impedance of the Grape Berry Cuticle as a Novel Phenotypic Trait to Estimate Resistance to Botrytis Cinerea. Sensors. 2015; 15(6):12498-12512. https://doi.org/10.3390/s150612498
Chicago/Turabian StyleHerzog, Katja, Rolf Wind, and Reinhard Töpfer. 2015. "Impedance of the Grape Berry Cuticle as a Novel Phenotypic Trait to Estimate Resistance to Botrytis Cinerea" Sensors 15, no. 6: 12498-12512. https://doi.org/10.3390/s150612498
APA StyleHerzog, K., Wind, R., & Töpfer, R. (2015). Impedance of the Grape Berry Cuticle as a Novel Phenotypic Trait to Estimate Resistance to Botrytis Cinerea. Sensors, 15(6), 12498-12512. https://doi.org/10.3390/s150612498