
Characterization of N-Acylhomoserine Lactones Produced by Bacteria Isolated from Industrial Cooling Water Systems


Abstract
:1. Introduction
2. Experimental Section
2.1. Isolation of Bacteria from Industrial Cooling Water Systems
2.2. Screening of AHL-Producing Bacteria
2.3. Identification of AHL-Producing Bacteria
2.4. Extraction of AHLs
2.5. AHL Identification by Liquid Chromatography-Mass Spectrometry (LC-MS)
2.6. Nucleotide Sequence Accession Numbers
3. Results and Discussion
3.1. Isolation and Identification of AHL-Producing Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | Related Type Strain | Identity | CV026 | VIR07 |
|---|---|---|---|---|---|
| D10 | Factory B | Aeromonas hydrophila | 100% | ++ | + |
| F6 | Factory C | Aeromonas hydrophila | 100% | ++ | + |
| F13 | Factory C | Lysobacter brunescens | 96.5% | - | ++ |
| D7 | Factory E | Methylobacterium oryzae | 99.5% | + | + |
| E9 | Factory E | Bosea massiliensis | 100% | ++ | + |
| F9 | Factory E | Methylobacterium oryzae | 99.5% | - | + |
| F12 | Factory E | Bosea massiliensis | 100% | ++ | + |
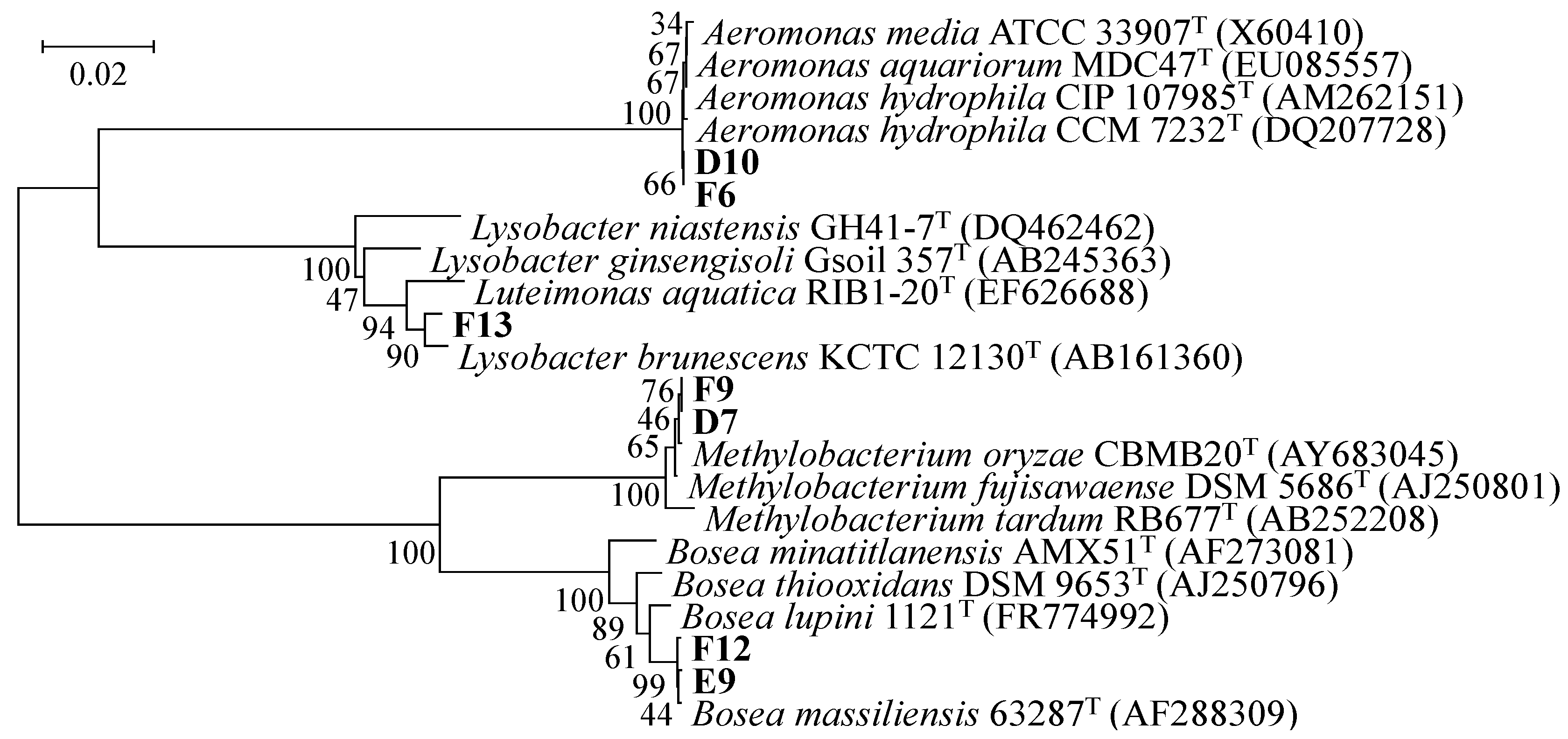
3.2. Phylogenetic Analysis of AHL-Producing Isolates

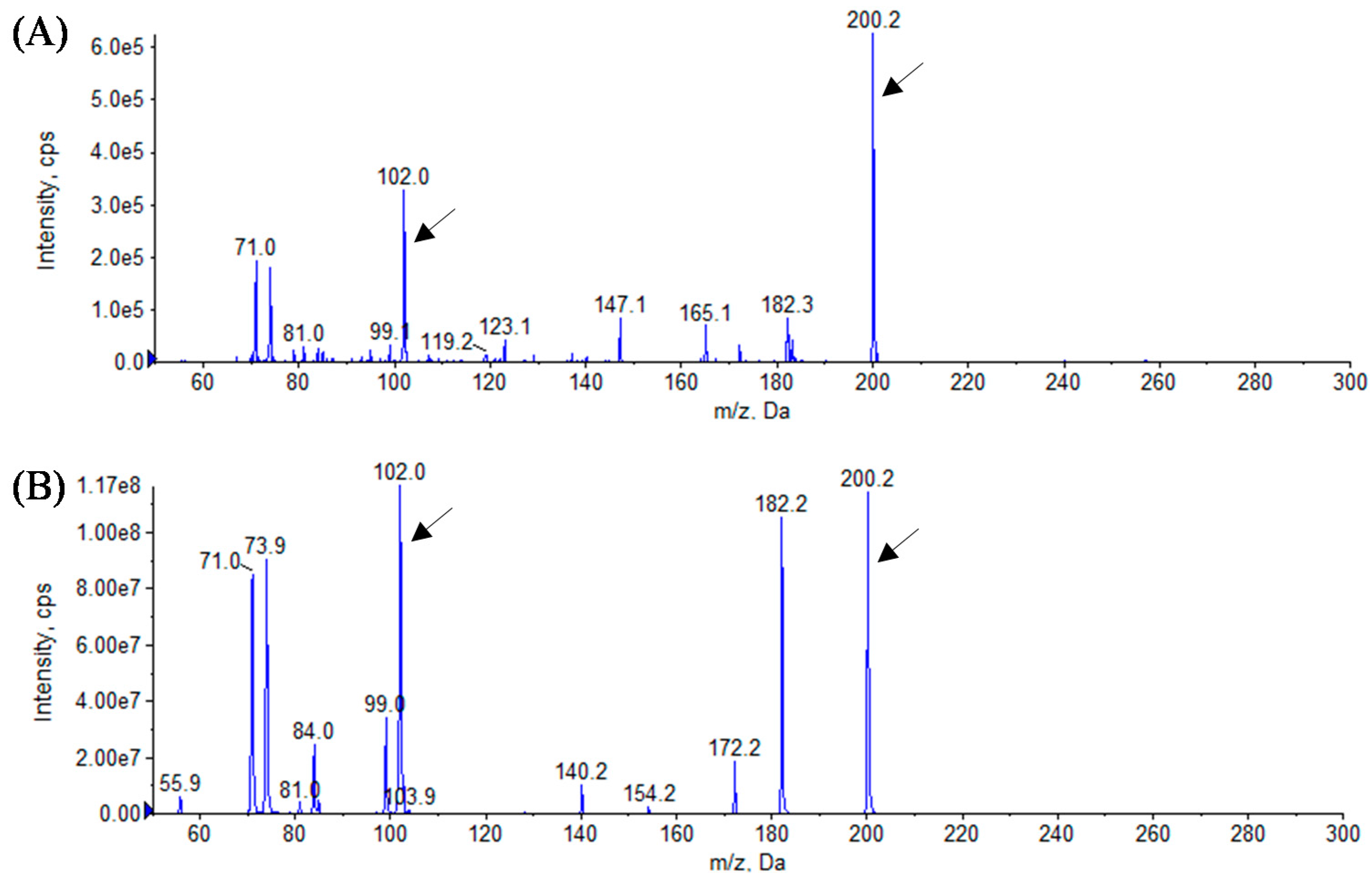
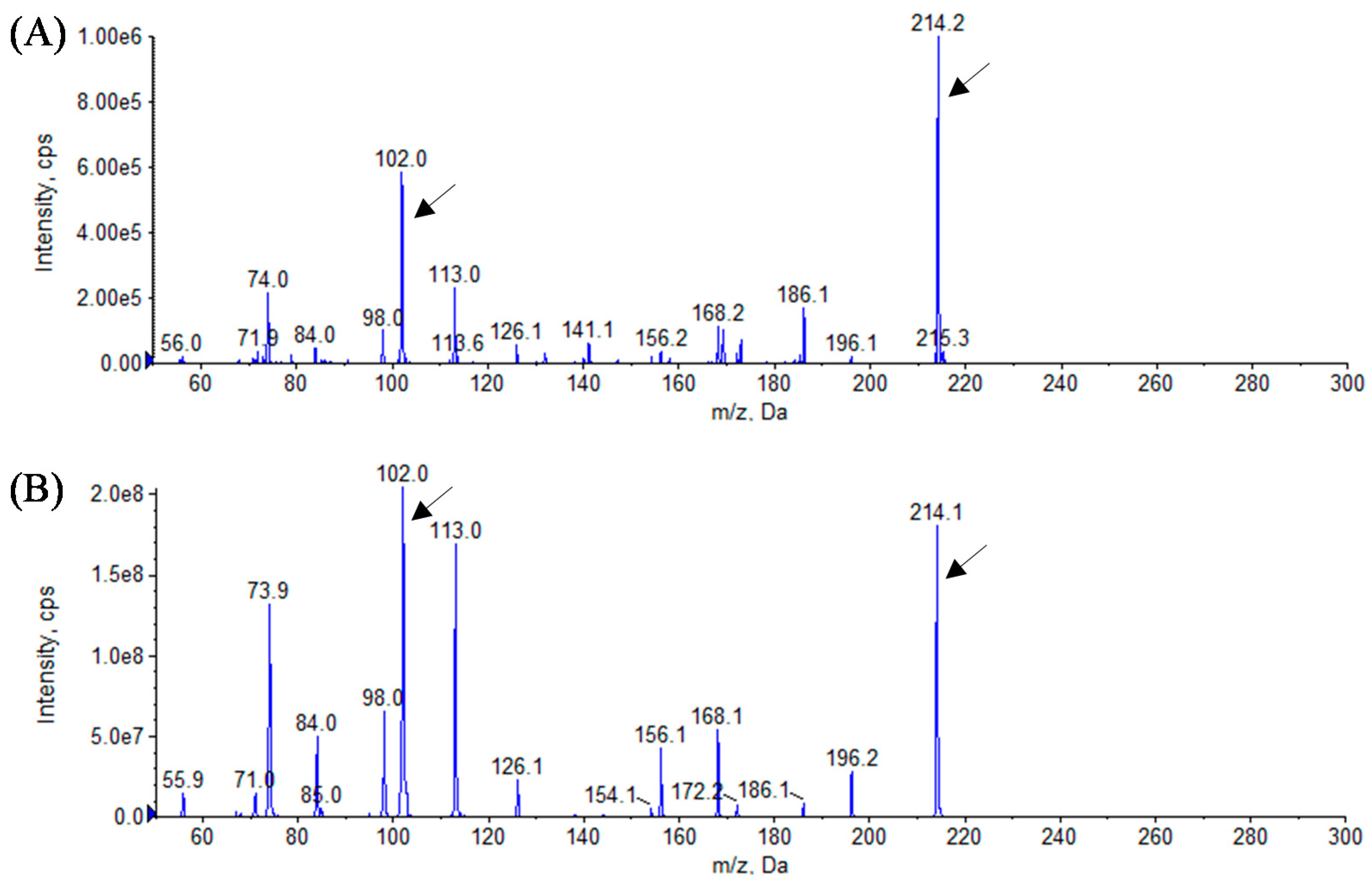
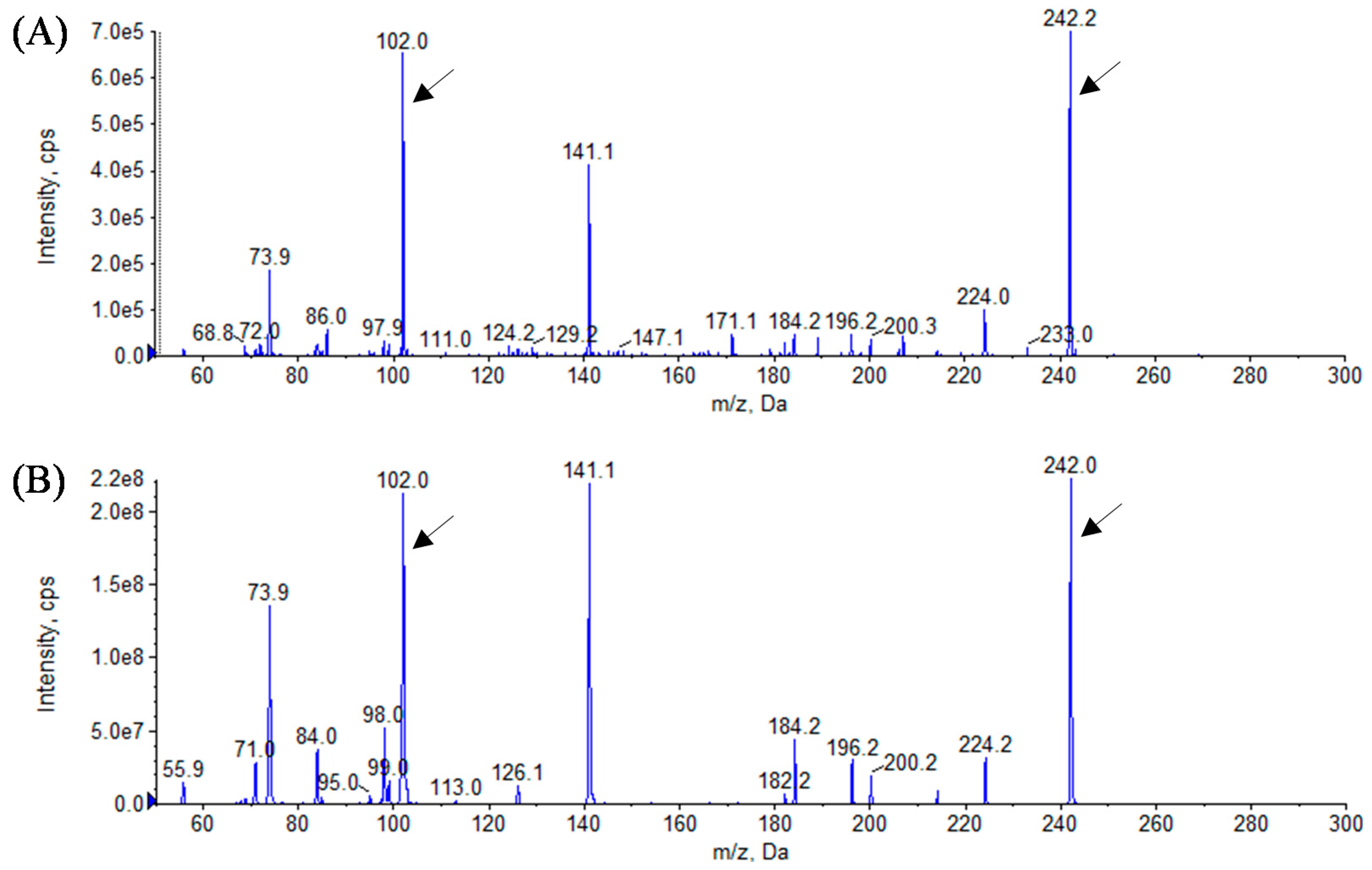
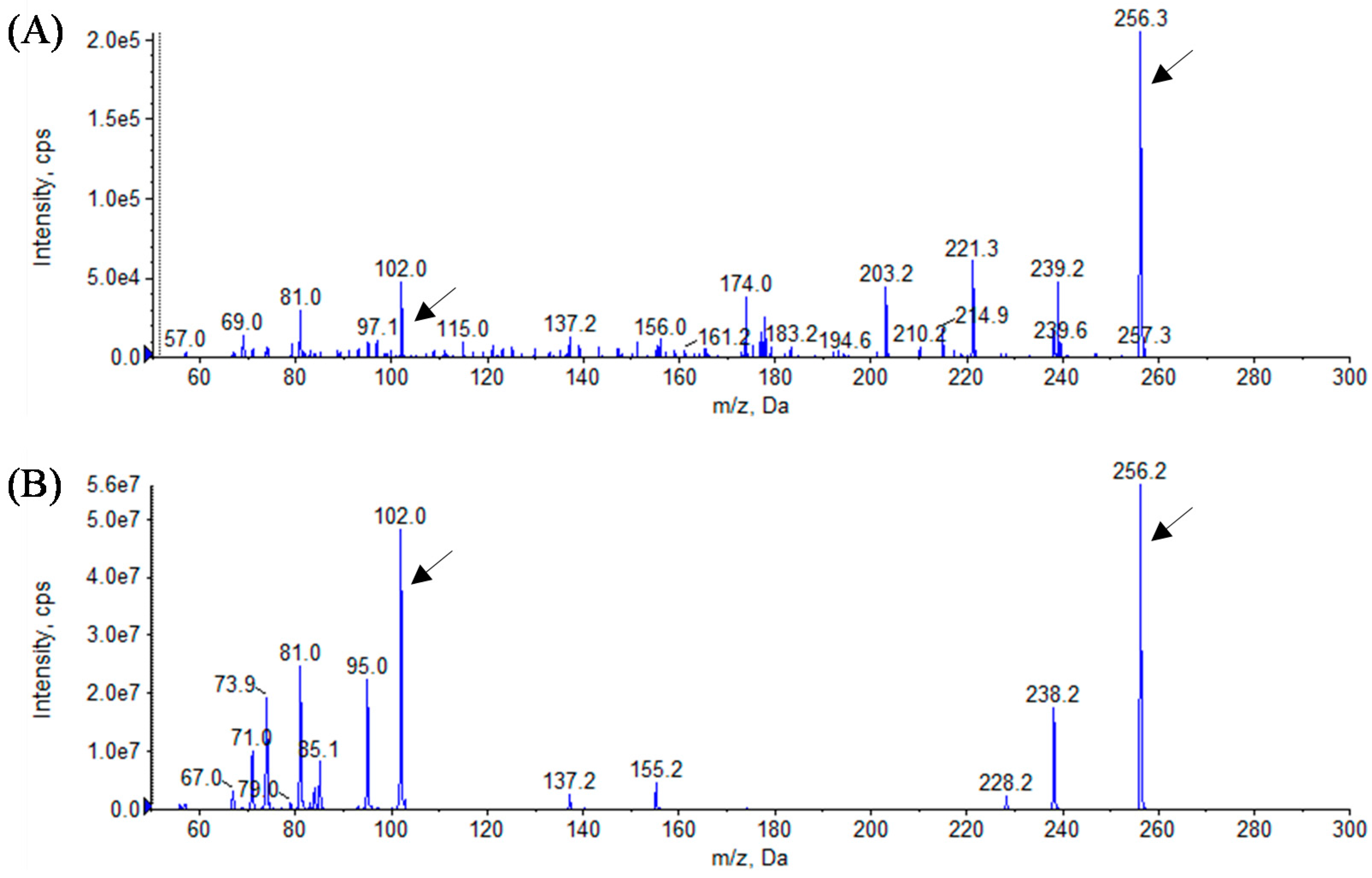
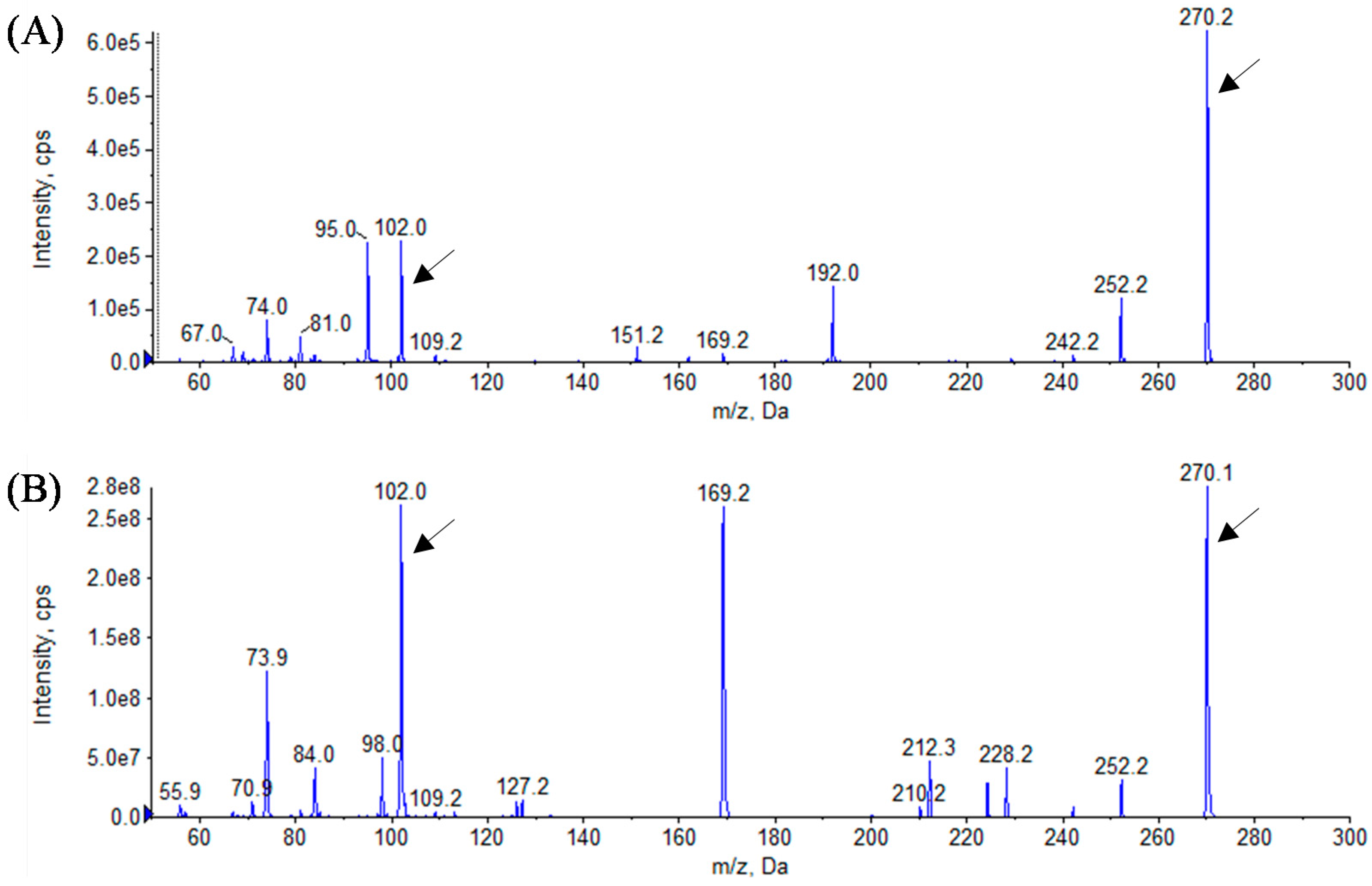
3.3. Analytical Identification of AHL Molecules






4. Conclusions/Outlook
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell. Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Greenberg, E.P. Acyl-homoserine lactone quorum sensing in gram-negative bacteria: A signaling mechanism involved in associations with higher organisms. Proc. Natl. Acad. Sci. USA 2000, 97, 8789–8793. [Google Scholar] [CrossRef] [PubMed]
- Schuster, M.; Sexton, D.J.; Diggle, S.P.; Greenberg, E.P. Acyl-homoserine lactone quorum sensing: From evolution to application. Annu. Rev. Microbiol. 2013, 67, 43–63. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Dessaux, Y.; Oger, P. Quorum sensing and quorum quenching: the yin and yang of bacterial communication. ChemBioChem 2009, 10, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, M.; Xiao, H.; Wu, W.; Xie, M.; Sun, M.; Zhu, C.; Li, P. Bacterial community structure in cooling water and biofilm in an industrial recirculating cooling water system. Water Sci Technol. 2013, 68, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Lade, H.; Paul, D.; Kweon, J.H. Quorum quenching mediated approaches for control of membrane biofouling. Int. J. Biol. Sci. 2014, 10, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Yeon, K.M.; Cheong, W.S.; Oh, H.S.; Lee, W.N.; Hwang, B.K.; Lee, C.H.; Beyenal, H.; Lewandowski, Z. Quorum sensing: A new biofouling control paradigm in a membrane bioreactor for advanced wastewater treatment. Environ. Sci. Technol. 2009, 43, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Case, R.J.; Labbate, M.; Kjelleberg, S. AHL-driven quorum-sensing circuits: their frequency and function among the Proteobacteria. ISME J. 2008, 2, 345–349. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Cámara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Brcroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, T.; Kato, M.; Fukamachi, K.; Kato, N.; Ikeda, T. N-acylhomoserine lactone regulates violacein production in Chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol. Lett. 2008, 279, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, S.; Morohoshi, T.; Kurabeishi, A.; Shinozaki, M.; Fujita, H.; Sawada, I.; Ikeda, T. Production and degradation of N-acylhomoserine lactone quorum sensing signal molecules in bacteria isolated from activated sludge. Biosci. Biotechnol. Biochem. 2013, 77, 2436–2440. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam, S.A.; McGarrell, D.M.; Garrity, G.M.; Tiedje, J.M. The Ribosomal Database Project (RDP-II): Sequences and tools for high-throughput rRNA analysis. Nucleic Acids Res. 2005, 33, D294–D296. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, S.R.; Harty, C.; Hooi, D.S.; Daykin, M.; Williams, P.; Telford, G.; Pritchard, D.I.; Bycroft, B.W. Synthetic analogues of the bacterial signal (quorum sensing) molecule N-(3-oxododecanoyl)-l-homoserine lactone as immune modulators. J. Med. Chem. 2003, 46, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Valle, A.; Bailey, M.J.; Whiteley, A.S.; Manefield, M. N-acyl-l-homoserine lactones (AHLs) affect microbial community composition and function in activated sludge. Environ. Microbiol. 2004, 6, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Furuhata, K.; Kato, Y.; Goto, K.; Hara, M.; Yoshida, S.; Fukuyama, M. Isolation and identification of methylobacterium species from the tap water in hospitals in Japan and their antibiotic susceptibility. Microbiol. Immunol. 2006, 50, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Poonguzhali, S.; Madhaiyan, M.; Sa, T. Production of acyl-homoserine lactone quorum-sensing signals is widespread in gram-negative Methylobacterium. J. Microbiol. Biotechnol. 2007, 17, 226–233. [Google Scholar] [PubMed]
- Yano, T.; Kubota, H.; Hanai, J.; Hitomi, J.; Tokuda, H. Stress tolerance of Methylobacterium biofilms in bathrooms. Microbes Environ. 2013, 28, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Oger, P.M.; Mansouri, H.; Nesme, X.; Dessaux, Y. Engineering root exudation of Lotus toward the production of two novel carbon compounds leads to the selection of distinct microbial populations in the rhizosphere. Microb. Ecol. 2014, 47, 96–103. [Google Scholar] [CrossRef] [PubMed]
- D'Angelo-Picard, C.; Faure, D.; Penot, I.; Dessaux, Y. Diversity of N-acyl homoserine lactone-producing and -degrading bacteria in soil and tobacco rhizosphere. Environ. Microbiol. 2005, 7, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Wang, Y.; Liu, Y.; Xu, F.; He, Y.W.; Du, L.; Venturi, V.; Fan, J.; Hu, B.; Liu, F. Lysobacter enzymogenes uses two distinct cell-cell signaling systems for differential regulation of secondary-metabolite biosynthesis and colony morphology. Appl. Environ. Microbiol. 2013, 79, 6604–6616. [Google Scholar] [CrossRef] [PubMed]
- Nadell, C.D.; Xavier, J.B.; Levin, S.A.; Foster, K.R. The evolution of quorum sensing in bacterial biofilms. PLoS Biol. 2008, 6. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.; Yeon, K.M.; Shim, J.; Kim, S.R.; Lee, C.H.; Lee, J.; Kim, J. Effective antifouling using quorum-quenching acylase stabilized in magnetically-separable mesoporous silica. Biomacromolecules 2014, 15, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okutsu, N.; Morohoshi, T.; Xie, X.; Kato, N.; Ikeda, T. Characterization of N-Acylhomoserine Lactones Produced by Bacteria Isolated from Industrial Cooling Water Systems. Sensors 2016, 16, 44. https://doi.org/10.3390/s16010044
Okutsu N, Morohoshi T, Xie X, Kato N, Ikeda T. Characterization of N-Acylhomoserine Lactones Produced by Bacteria Isolated from Industrial Cooling Water Systems. Sensors. 2016; 16(1):44. https://doi.org/10.3390/s16010044
Chicago/Turabian StyleOkutsu, Noriya, Tomohiro Morohoshi, Xiaonan Xie, Norihiro Kato, and Tsukasa Ikeda. 2016. "Characterization of N-Acylhomoserine Lactones Produced by Bacteria Isolated from Industrial Cooling Water Systems" Sensors 16, no. 1: 44. https://doi.org/10.3390/s16010044
APA StyleOkutsu, N., Morohoshi, T., Xie, X., Kato, N., & Ikeda, T. (2016). Characterization of N-Acylhomoserine Lactones Produced by Bacteria Isolated from Industrial Cooling Water Systems. Sensors, 16(1), 44. https://doi.org/10.3390/s16010044