Use of Cyanobacterial Luminescent Bioreporters to Report on the Environmental Impact of Metallic Nanoparticles



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Biological Materials and Culture Conditions
2.2. Chemicals
2.3. Characterization of NPs
2.4. Bioluminescence Assays
2.5. Spiking Experiments: Performance of the ROS-Detecting Bioreporters in Environmental Matrices Artificially Contaminated with Metallic NPs
2.6. Statistical Analysis
3. Results
3.1. Metallic NPs Physicochemical Characterization
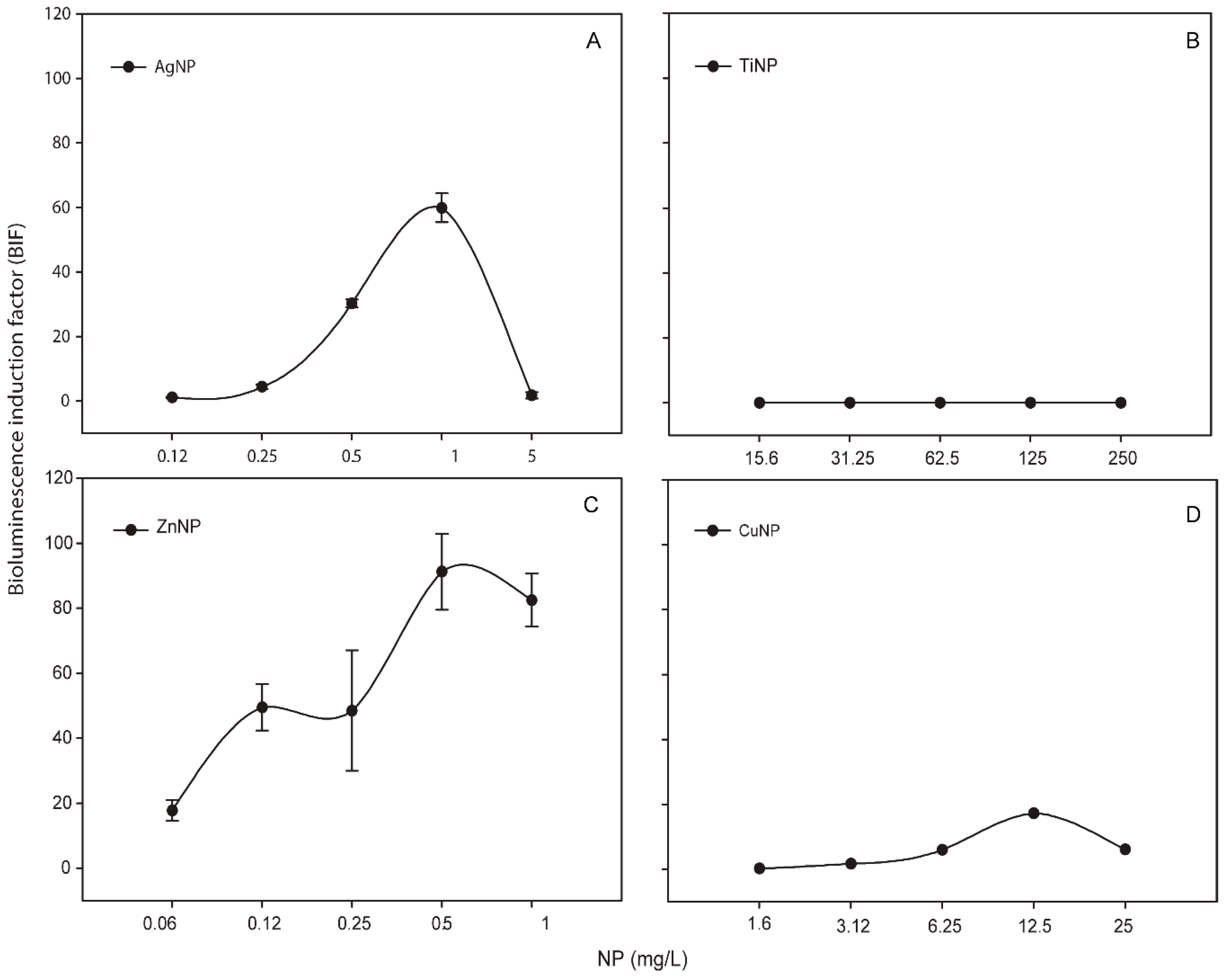
3.2. Response of Nostoc sp. CPB4337 to Metallic NPs: Toxicity
3.3. Response of Synechococcus Elongatus sp. PCC7942 pBG2120 to Released Free-Ions from Metallic NPs
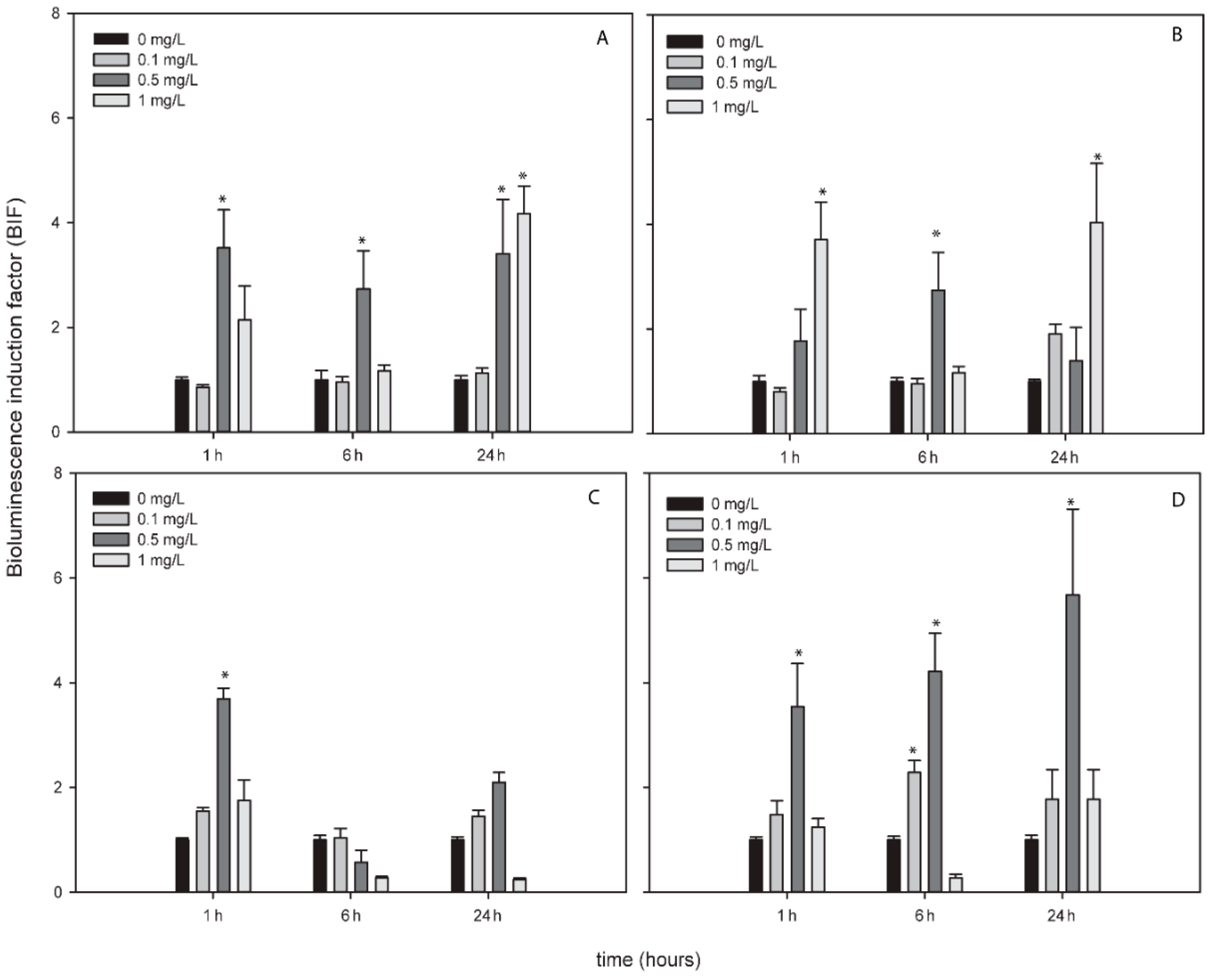
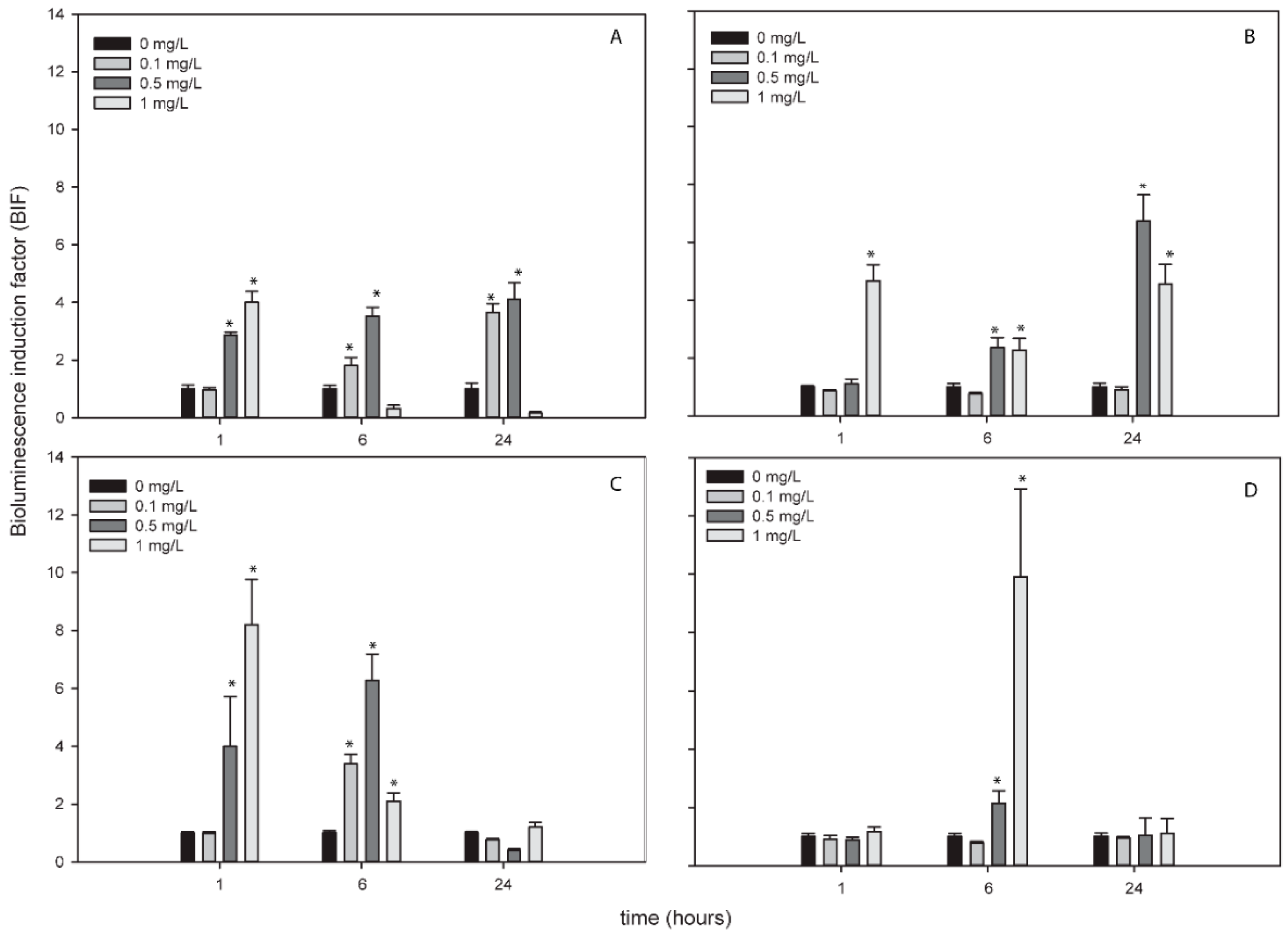
3.4. Response of ROS-Detecting Nostoc sp. PCC7120 Bioreporters to Metallic NPs
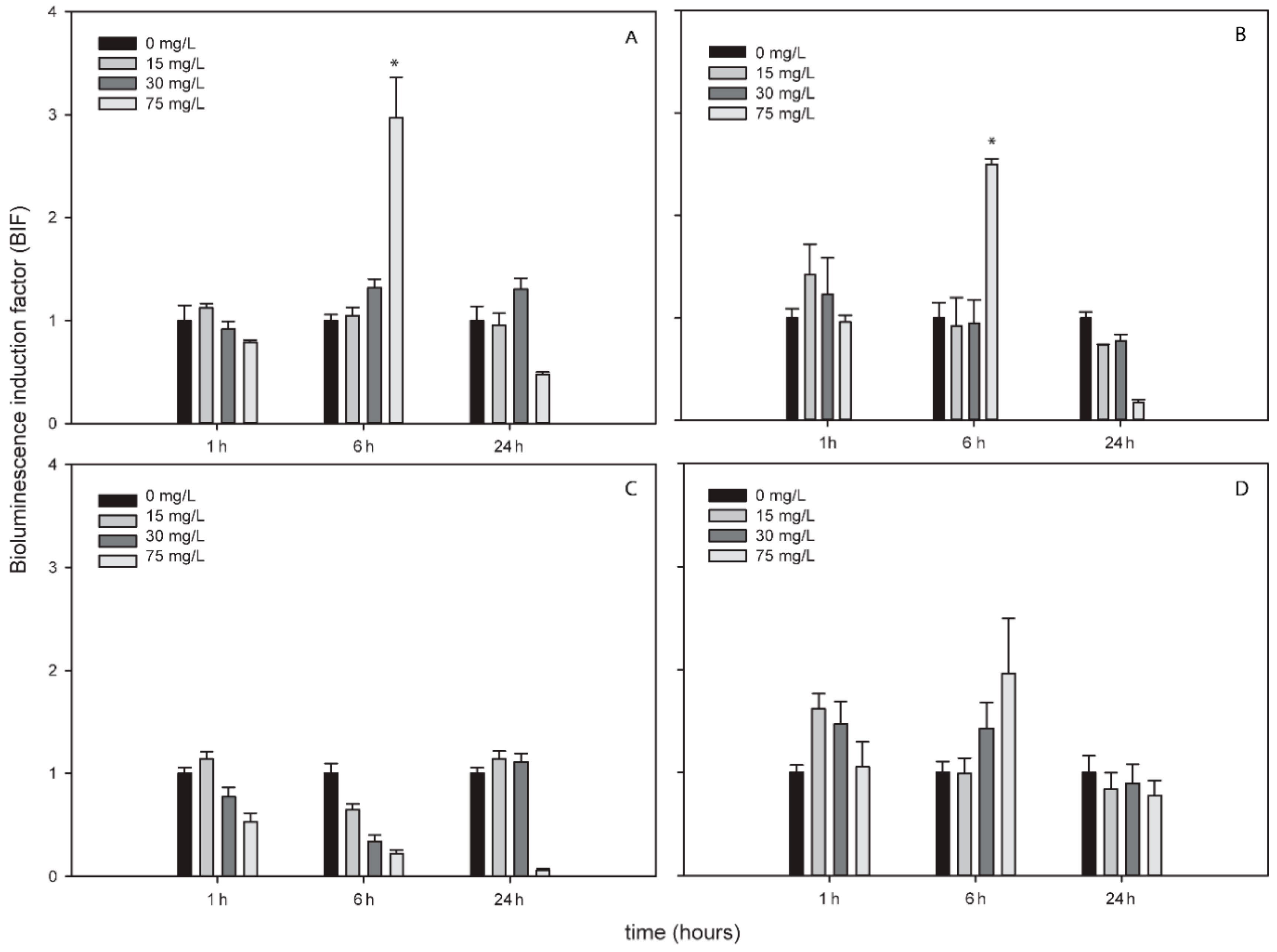
Response of ROS-Detecting Nostoc sp. PCC7120 Bioreporters to Metallic NPs Added to Environmental Water Samples (Spiking Experiments)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z. Heavy metals as essential nutrients. In Heavy Metal Stress in Plants; Springer: Berlin/Heidelberg, Germany, 2004; pp. 271–294. [Google Scholar]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metal toxicity and the environment. In Molecular, Clinical and Environmental Toxicology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 133–164. [Google Scholar]
- Rodea-Palomares, I.; Fernández-Piñas, F.; González-García, C.; Leganés, F. Use of lux-marked cyanobacterial bioreporters for assessment of individual and combined toxicities of metals in aqueous samples. Handb. Cyanobact. Biochem. Biotechnol. Appl. 2009, 283–304. [Google Scholar]
- Durán, N.; Durán, M.; de Jesus, M.B.; Seabra, A.B.; Fávaro, W.J.; Nakazato, G. Silver nanoparticles: A new view on mechanistic aspects on antimicrobial activity. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Abdal Dayem, A.; Hossain, M.; Lee, S.; Kim, K.; Saha, S.; Yang, G.-M.; Choi, H.; Cho, S.-G. The role of reactive oxygen species (ROS) in the biological activities of metallic nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Anreddy, R.N.R.; Yellu, N.R.; Devarakonda, K.R. Oxidative biomarkers to assess the nanoparticle-induced oxidative stress. In Oxidative Stress and Nanotechnology; Springer: Berlin/Heidelberg, Germany, 2013; pp. 205–219. [Google Scholar]
- Anjum, N.A.; Gill, S.S.; Duarte, A.C.; Pereira, E. Oxidative Stress Biomarkers and Antioxidant Defense in Plants Exposed to Metallic Nanoparticles. In Nanomaterials and Plant Potential; Springer: Berlin/Heidelberg, Germany, 2019; pp. 427–439. [Google Scholar]
- Touyz, R.M. Molecular and cellular mechanisms in vascular injury in hypertension: Role of angiotensin II–editorial review. Curr. Opin. Nephrol. Hypertens. 2005, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Latifi, A.; Ruiz, M.; Zhang, C.-C. Oxidative stress in cyanobacteria. FEMS Microbiol. Rev. 2009, 33, 258–278. [Google Scholar] [CrossRef] [Green Version]
- Augusto, O.; Miyamoto, S. Oxygen radicals and related species. Princ. Free Radic. Biomed. 2011, 1, 19–42. [Google Scholar]
- Von Moos, N.; Slaveykova, V.I. Oxidative stress induced by inorganic nanoparticles in bacteria and aquatic microalgae–state of the art and knowledge gaps. Nanotoxicology 2014, 8, 605–630. [Google Scholar] [CrossRef]
- Stone, V.; Donaldson, K. Signs of stress. Nat. Nanotechnol. 2006, 1, 23–24. [Google Scholar] [CrossRef]
- Niazi, J.H.; Gu, M.B. Toxicity of metallic nanoparticles in microorganisms—A review. In Atmospheric and Biological Environmental Monitoring; Springer: Berlin/Heidelberg, Germany, 2009; pp. 193–206. [Google Scholar]
- Zuberek, M.; Grzelak, A. Nanoparticles-caused oxidative imbalance. In Cellular and Molecular Toxicology of Nanoparticles; Springer: Berlin/Heidelberg, Germany, 2018; pp. 85–98. [Google Scholar]
- Handy, R.D.; van den Brink, N.; Chappell, M.; Mühling, M.; Behra, R.; Dušinská, M.; Simpson, P.; Ahtiainen, J.; Jha, A.N.; Seiter, J. Practical considerations for conducting ecotoxicity test methods with manufactured nanomaterials: What have we learnt so far? Ecotoxicology 2012, 21, 933–972. [Google Scholar] [CrossRef]
- Eduok, S.; Martin, B.; Villa, R.; Nocker, A.; Jefferson, B.; Coulon, F. Evaluation of engineered nanoparticle toxic effect on wastewater microorganisms: Current status and challenges. Ecotoxicol. Environ. Saf. 2013, 95, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Duester, L.; Burkhardt, M.; Gutleb, A.C.; Kaegi, R.; Macken, A.; Meermann, B.; von der Kammer, F. Toward a comprehensive and realistic risk evaluation of engineered nanomaterials in the urban water system. Front. Chem. 2014, 2, 39. [Google Scholar] [CrossRef]
- Girotti, S.; Ferri, E.N.; Fumo, M.G.; Maiolini, E. Monitoring of environmental pollutants by bioluminescent bacteria. Anal. Chim. Acta 2008, 608, 2–29. [Google Scholar] [CrossRef]
- Mallevre, F.; Alba, C.; Milne, C.; Gillespie, S.; Fernandes, T.; Aspray, T. Toxicity testing of pristine and aged silver nanoparticles in real wastewaters using bioluminescent Pseudomonas putida. Nanomaterials 2016, 6, 49. [Google Scholar] [CrossRef]
- Holden, P.A.; Nisbet, R.M.; Lenihan, H.S.; Miller, R.J.; Cherr, G.N.; Schimel, J.P.; Gardea-Torresdey, J.L. Ecological nanotoxicology: Integrating nanomaterial hazard considerations across the subcellular, population, community, and ecosystems levels. Acc. Chem. Res. 2012, 46, 813–822. [Google Scholar] [CrossRef]
- Sadik, O.; Zhou, A.; Kikandi, S.; Du, N.; Wang, Q.; Varner, K. Sensors as tools for quantitation, nanotoxicity and nanomonitoring assessment of engineered nanomaterials. J. Environ. Monit. 2009, 11, 1782–1800. [Google Scholar] [CrossRef]
- Whitton, B.A.; Potts, M. The Ecology of Cyanobacteria: Their Diversity in Time and Space; Springer Science & Business Media: Berlin, Germany, 2007. [Google Scholar]
- Mur, R.; Skulberg, O.M.; Utkilen, H. Cyanobacteria in the Environment; WHO: Geneva, Switzerland, 1999. [Google Scholar]
- Flores, F.G. The Cyanobacteria: Molecular Biology, Genomics, and Evolution; Horizon Scientific Press: Sevilla, Spain, 2008. [Google Scholar]
- Harms, H.; Wells, M.C.; van der Meer, J.R. Whole-cell living biosensors—Are they ready for environmental application? Appl. Microbiol. Biotechnol. 2006, 70, 273–280. [Google Scholar] [CrossRef]
- Van Der Meer, J.R.; Belkin, S. Where microbiology meets microengineering: Design and applications of reporter bacteria. Nat. Rev. Microbiol. 2010, 8, 511. [Google Scholar] [CrossRef]
- Rodea-Palomares, I.; Gonzalez-Garcia, C.; Leganes, F.; Fernandez-Pinas, F. Effect of pH, EDTA, and anions on heavy metal toxicity toward a bioluminescent cyanobacterial bioreporter. Arch. Environ. Contam. Toxicol. 2009, 57, 477. [Google Scholar] [CrossRef]
- Hurtado-Gallego, J.; Martín-Betancor, K.; Rodea-Palomares, I.; Leganés, F.; Rosal, R.; Fernández-Piñas, F. Two novel cyanobacterial bioluminescent whole-cell bioreporters based on superoxide dismutases MnSod and FeSod to detect superoxide anion. Chemosphere 2018, 201, 772–779. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Gallego, J.; Redondo-López, A.; Leganés, F.; Rosal, R.; Fernández-Piñas, F. Peroxiredoxin (2-cys-prx) and catalase (katA) cyanobacterial-based bioluminescent bioreporters to detect oxidative stress in the aquatic environment. Chemosphere 2019, 236, 124395. [Google Scholar] [CrossRef]
- Martin-Betancor, K.; Rodea-Palomares, I.; Munoz-Martin, M.; Leganes, F.; Fernandez-Pinas, F. Construction of a self-luminescent cyanobacterial bioreporter that detects a broad range of bioavailable heavy metals in aquatic environments. Front. Microbiol. 2015, 6, 186. [Google Scholar] [CrossRef]
- Fernández-Pinas, F.; Wolk, C.P. Expression of luxCD-E in Anabaena sp. can replace the use of exogenous aldehyde for in vivo localization of transcription by luxAB. Gene 1994, 150, 169–174. [Google Scholar] [CrossRef]
- Gonzalo, S.; Llaneza, V.; Pulido-Reyes, G.; Fernández-Piñas, F.; Bonzongo, J.C.; Leganes, F.; Rosal, R.; García-Calvo, E.; Rodea-Palomares, I. A colloidal singularity reveals the crucial role of colloidal stability for nanomaterials in-vitro toxicity testing: nZVI-microalgae colloidal system as a case study. PLoS ONE 2014, 9, e109645. [Google Scholar] [CrossRef]
- Rosal, R.; Rodea-Palomares, I.; Boltes, K.; Fernández-Piñas, F.; Leganés, F.; Gonzalo, S.; Petre, A. Ecotoxicity assessment of lipid regulators in water and biologically treated wastewater using three aquatic organisms. Environ. Sci. Pollut. Res. 2010, 17, 135–144. [Google Scholar] [CrossRef]
- Rodea-Palomares, I.; Leganés, F.; Rosal, R.; Fernández-Pinas, F. Toxicological interactions of perfluorooctane sulfonic acid (PFOS) and perfluorooctanoic acid (PFOA) with selected pollutants. J. Hazard. Mater. 2012, 201, 209–218. [Google Scholar] [CrossRef]
- González-Pleiter, M.; Gonzalo, S.; Rodea-Palomares, I.; Leganés, F.; Rosal, R.; Boltes, K.; Marco, E.; Fernández-Piñas, F. Toxicity of five antibiotics and their mixtures towards photosynthetic aquatic organisms: Implications for environmental risk assessment. Water Res. 2013, 47, 2050–2064. [Google Scholar] [CrossRef]
- Rodea-Palomares, I.; Boltes, K.; Fernández-Pinas, F.; Leganés, F.; García-Calvo, E.; Santiago, J.; Rosal, R. Physicochemical characterization and ecotoxicological assessment of CeO2 nanoparticles using two aquatic microorganisms. Toxicol. Sci. 2010, 119, 135–145. [Google Scholar] [CrossRef]
- Rodea-Palomares, I.; Gonzalo, S.; Santiago-Morales, J.; Leganés, F.; García-Calvo, E.; Rosal, R.; Fernández-Pinas, F. An insight into the mechanisms of nanoceria toxicity in aquatic photosynthetic organisms. Aquat. Toxicol. 2012, 122, 133–143. [Google Scholar] [CrossRef]
- Martín-de-Lucía, I.; Campos-Mañas, M.C.; Agüera, A.; Rodea-Palomares, I.; Pulido-Reyes, G.; Leganés, F.; Fernández-Piñas, F.; Rosal, R. Reverse Trojan-horse effect decreased wastewater toxicity in the presence of inorganic nanoparticles. Environ. Sci. Nano 2017, 4, 1273–1282. [Google Scholar] [CrossRef] [Green Version]
- Kahru, A.; Dubourguier, H.-C. From ecotoxicology to nanoecotoxicology. Toxicology 2010, 269, 105–119. [Google Scholar] [CrossRef]
- Kahru, A.; Ivask, A.; Blinova, I.; Kasemets, K.; Dubourguier, H.C. Bioavailability and ecotoxicology of nanoparticles. Toxicol. Lett. 2008, S20. [Google Scholar] [CrossRef]
- Douterelo, I.; Perona, E.; Mateo, P. Use of cyanobacteria to assess water quality in running waters. Environ. Pollut. 2004, 127, 377–384. [Google Scholar] [CrossRef]
- Barrán-Berdón, A.L.; Rodea-Palomares, I.; Leganés, F.; Fernández-Piñas, F. Free Ca2+ as an early intracellular biomarker of exposure of cyanobacteria to environmental pollution. Anal. Bioanal. Chem. 2011, 400, 1015–1029. [Google Scholar] [CrossRef]
- Rosal, R.; Rodríguez, A.; Perdigón-Melón, J.A.; Petre, A.; García-Calvo, E.; Gómez, M.J.; Agüera, A.; Fernández-Alba, A.R. Occurrence of emerging pollutants in urban wastewater and their removal through biological treatment followed by ozonation. Water Res. 2010, 44, 578–588. [Google Scholar] [CrossRef]
- Ritz, C.; Streibig, J.C. Bioassay analysis using R. J. Stat. Softw 2005, 12, 1–22. [Google Scholar] [CrossRef]
- Pulido-Reyes, G.; Leganes, F.; Fernández-Piñas, F.; Rosal, R. Bio-nano interface and environment: A critical review. Environ. Toxicol. Chem. 2017, 36, 3181–3193. [Google Scholar] [CrossRef]
- Mazumder, B.; Ray, S.; Pal, P.; Pathak, Y. Nanotechnology: Therapeutic, Nutraceutical, and Cosmetic Advances; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Hurtado-Gallego, J.; Pulido-Reyes, G.; González-Pleiter, M.; Fernández-Piñas, F. Luminescent Microbial Bioassays and Microalgal Biosensors as Tools for Environmental Toxicity Evaluation. In Handbook of Cell Biosensors; Thouand, G., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–58. [Google Scholar]
- Pulido-Reyes, G.; Martín, E.; Coronado, J.G.; Leganes, F.; Rosal, R.; Fernández-Piñas, F. Physicochemical and biological interactions between cerium oxide nanoparticles and a 1,8-naphthalimide derivative. J. Photochem. Photobiol. B Biol. 2017, 172, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Bundschuh, M.; Seitz, F.; Rosenfeldt, R.R.; Schulz, R. Effects of nanoparticles in fresh waters: Risks, mechanisms and interactions. Freshw. Biol. 2016, 61, 2185–2196. [Google Scholar] [CrossRef]
- McShan, D.; Ray, P.C.; Yu, H. Molecular toxicity mechanism of nanosilver. J. Food Drug Anal. 2014, 22, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Ivask, A.; Bondarenko, O.; Jepihhina, N.; Kahru, A. Profiling of the reactive oxygen species-related ecotoxicity of CuO, ZnO, TiO2, silver and fullerene nanoparticles using a set of recombinant luminescent Escherichia coli strains: Differentiating the impact of particles and solubilised metals. Anal. Bioanal. Chem. 2010, 398, 701–716. [Google Scholar] [CrossRef]
- Hwang, E.T.; Lee, J.H.; Chae, Y.J.; Kim, Y.S.; Kim, B.C.; Sang, B.I.; Gu, M.B. Analysis of the toxic mode of action of silver nanoparticles using stress-specific bioluminescent bacteria. Small 2008, 4, 746–750. [Google Scholar] [CrossRef]
- Völker, C.; Oetken, M.; Oehlmann, J. The biological effects and possible modes of action of nanosilver. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 223, pp. 81–106. [Google Scholar]
- Nair, P.M.G.; Park, S.Y.; Choi, J. Evaluation of the effect of silver nanoparticles and silver ions using stress responsive gene expression in Chironomus riparius. Chemosphere 2013, 92, 592–599. [Google Scholar] [CrossRef]
- Tlili, A.; Cornut, J.; Behra, R.; Gil-Allué, C.; Gessner, M.O. Harmful effects of silver nanoparticles on a complex detrital model system. Nanotoxicology 2016, 10, 728–735. [Google Scholar] [CrossRef]
- Shah, S.N.A.; Shah, Z.; Hussain, M.; Khan, M. Hazardous effects of titanium dioxide nanoparticles in ecosystem. Bioinorg. Chem. Appl. 2017, 2017, 4101735. [Google Scholar] [CrossRef]
- Ramsden, C.; Henry, T.; Handy, R. Sub-lethal effects of titanium dioxide nanoparticles on the physiology and reproduction of zebrafish. Aquat. Toxicol. 2013, 126, 404–413. [Google Scholar] [CrossRef]
- Ates, M.; Demir, V.; Adiguzel, R.; Arslan, Z. Bioaccumulation, subacute toxicity, and tissue distribution of engineered titanium dioxide nanoparticles in goldfish (Carassius auratus). J. Nanomater. 2013, 2013, 9. [Google Scholar] [CrossRef]
- Iswarya, V.; Bhuvaneshwari, M.; Alex, S.A.; Iyer, S.; Chaudhuri, G.; Chandrasekaran, P.T.; Bhalerao, G.M.; Chakravarty, S.; Raichur, A.M.; Chandrasekaran, N. Combined toxicity of two crystalline phases (anatase and rutile) of Titania nanoparticles towards freshwater microalgae: Chlorella sp. Aquat. Toxicol. 2015, 161, 154–169. [Google Scholar] [CrossRef]
- Sharma, V.K. Aggregation and toxicity of titanium dioxide nanoparticles in aquatic environment—A review. J. Environ. Sci. Health Part A 2009, 44, 1485–1495. [Google Scholar] [CrossRef]
- Maness, P.-C.; Smolinski, S.; Blake, D.M.; Huang, Z.; Wolfrum, E.J.; Jacoby, W.A. Bactericidal activity of photocatalytic TiO2 reaction: Toward an understanding of its killing mechanism. Appl. Environ. Microbiol. 1999, 65, 4094–4098. [Google Scholar]
- Zhao, X.; Wang, S.; Wu, Y.; You, H.; Lv, L. Acute ZnO nanoparticles exposure induces developmental toxicity, oxidative stress and DNA damage in embryo-larval zebrafish. Aquat. Toxicol. 2013, 136, 49–59. [Google Scholar] [CrossRef]
- Huang, C.-C.; Aronstam, R.S.; Chen, D.-R.; Huang, Y.-W. Oxidative stress, calcium homeostasis, and altered gene expression in human lung epithelial cells exposed to ZnO nanoparticles. Toxicol. Vitr. 2010, 24, 45–55. [Google Scholar] [CrossRef]
- Applerot, G.; Lellouche, J.; Lipovsky, A.; Nitzan, Y.; Lubart, R.; Gedanken, A.; Banin, E. Understanding the antibacterial mechanism of CuO nanoparticles: Revealing the route of induced oxidative stress. Small 2012, 8, 3326–3337. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, S.; Wang, Z.; Ye, N.; Fang, H.; Wang, D. TiO2, SiO2 and ZrO2 nanoparticles synergistically provoke cellular oxidative damage in freshwater microalgae. Nanomaterials 2018, 8, 95. [Google Scholar] [CrossRef]
- Xia, B.; Chen, B.; Sun, X.; Qu, K.; Ma, F.; Du, M. Interaction of TiO2 nanoparticles with the marine microalga Nitzschia closterium: Growth inhibition, oxidative stress and internalization. Sci. Total Environ. 2015, 508, 525–533. [Google Scholar] [CrossRef]
- Wang, F.; Guan, W.; Xu, L.; Ding, Z.; Ma, H.; Ma, A.; Terry, N. Effects of Nanoparticles on Algae: Adsorption, Distribution, Ecotoxicity and Fate. Appl. Sci. 2019, 9, 1534. [Google Scholar] [CrossRef]
- Levard, C.; Hotze, E.M.; Lowry, G.V.; Brown Jr, G.E. Environmental transformations of silver nanoparticles: Impact on stability and toxicity. Environ. Sci. Technol. 2012, 46, 6900–6914. [Google Scholar] [CrossRef]
- Lee, S.; Kim, K.; Shon, H.; Kim, S.D.; Cho, J. Biotoxicity of nanoparticles: Effect of natural organic matter. J. Nanopart. Res. 2011, 13, 3051–3061. [Google Scholar] [CrossRef]
- Blinova, I.; Ivask, A.; Heinlaan, M.; Mortimer, M.; Kahru, A. Ecotoxicity of nanoparticles of CuO and ZnO in natural water. Environ. Pollut. 2010, 158, 41–47. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene System | Characteristics and Culture Conditions | References | |
|---|---|---|---|
| Nostoc sp. CPB4337 | luxCDABE genes in the chromosome | Toxicity bioreporter based on Nostoc sp. PCC7120. NmR in AA/8+N growth medium | [32] |
| Synechococcus elongatus sp. PCC7942 pBG2120 | Plasmid pBG2120: Psmt::luxCDABE smt encodes the transcriptional SmtB and the metallothionein SmtA | Bioavailable heavy metal bioreporter based on Synechococcus elongatus sp. PCC7942 expressing the plasmid pBG2120. CmR in BG11 growth medium | [31] |
| Nostoc sp. PCC7120 pBG2154 | Plasmid pBG2154; PsodA::luxCDABE sodA encodes a Mn-superoxide dismutase | Specific superoxide anion bioreporter based on Nostoc sp. PCC7120 expressing the plasmid pBG2154. SpR in AA/8+N growth medium | [29] |
| Nostoc sp. PCC7120 pBG2165 | Plasmid pBG2165; PsodB::luxCDABE sodB encodes a Fe-superoxide dismutase enzyme | Specific superoxide anion bioreporter based on Nostoc sp. PCC7120 expressing the plasmid pBG2165. SpR in AA/8+N growth medium | [29] |
| Nostoc sp. PCC7120 pBG2172 | Plasmid pBG2172; P2-cys-prx:: luxCDABE 2-cys-prx encodes a peroxiredoxin enzyme | Superoxide anion and H2O2 bioreporter based on Nostoc sp. PCC 7120 expressing the plasmid pBG2172. SpR in AA/8+N growth medium | [30] |
| Nostoc sp. PCC7120 pBG2173 | Plasmid pBG2173; PkatA:: luxCDABE katA encodes a Mn-catalase enzyme | Specific superoxide anion bioreporter based on Nostoc sp. PCC 7120 expressing the plasmid pBG2173. SpR in AA/8+N | [30] |
| Size (nm) | |||||
|---|---|---|---|---|---|
| Ultrapure Water (pH 6.5) | AA/8+N (pH 7) | BG11 (pH 7.6) | Glx1 (pH 6.9) | WWTP (pH 7.5) | |
| Without NPs | - | 711.0 ± 181.18 171.8 ± 36.4 | 772.1 ± 122.6 | 227.9 ± 49.6 | 148.4 ± 10.2 |
| AgNPs | 55.7 ± 20.5 9.2 ± 3.3 | 635.9 ± 154.9 46.6 ± 10.4 8.5 ± 1.9 | 306.8 ± 39.1 28.96 ± 3.0 | 76.7 ± 38.1 10.3 ± 4.4 | 169.4 ± 110.0 16.5 ± 4.8 |
| TiNPs | 2333.0 ± 377.5 | 1197.0 ± 87.4 | 5433.0 ± 283.0 | 4213.0 ± 909.9 | 1099.0 ± 114.0 |
| ZnNPs | 264.8 ± 55 | 637.0 ± 120.3 | 1193.0 ± 87.5 | - | 509.3 ± 118.3 |
| CuNPs | 242.0 ± 120.2 | 204.6 ± 73.9 | 921.2±112.8 | 256.2 ± 91.1 | 249.1 ± 99.7 |
| ζ-potential (mV) | |||||
| Without NPs | - | −27.4 ± 1.0 | −11.51 ± 2.8 | −9.8 ± 1.4 | −9.9 ± 2.2 |
| AgNPs | −10.9 ± 2.4 | −24.2 ± 1.7 | −21.0 ± 11.8 | −4.7 ± 0.3 | −11.7 ± 11.5 |
| TiNPs | −17.1 ± 2.1 | −23.6 ± 1.0 | −18.2 ± 2.0 | −17.2 ± 0.8 | −12.5 ± 0.7 |
| ZnNPs | −17.4 ± 0.6 | −27.5 ± 1.3 | −19.0 ± 2.1 | −13.7 ± 3.3 | −13.3 ± 1.4 |
| CuNPs | −24.1 ± 0.2 | −28.7 ± 0.9 | −26.1 ± 1.0 | −22.4 ± 0.4 | −14.5 ± 0.1 |
| Time (h) | Metallic NPs | “R” Model Fitted | IC10 (mg/L) | IC50 (mg/L) | IC90 (mg/L) |
|---|---|---|---|---|---|
| 1 | Ag | LL.4 | 0.13 ± 0.04 | 0.23 ± 0.04 | 0.43 ± 0.17 |
| Ti | - | - | - | - | |
| Zn | LL2.3 | - | 2.04 ± 0.8 | 17.9 ± 5.5 | |
| Cu | W1.3 | 17.01 ± 3 | 32.8 ± 2.1 | 50.01 ± 4.23 | |
| 6 | Ag | W1.3 | 0.02 ± 0.02 | 0.14 ± 0.03 | 1.92 ± 0.6 |
| Ti | W1.4 | 0.55 ± 0.4 | 8.94 ± 2.0 | 52.77 ± 25.0 | |
| Zn | LL2.3 | - | 1.23 ± 0.1 | 2.96 ± 0.2 | |
| Cu | W2.3 | 14.06 ± 3.2 | 18.58 ± 2.26 | 28.77 ± 2.04 | |
| 24 | Ag | W1.3 | 0.4 ± 0.04 | 0.71 ± 0.03 | 1.03 ± 0.05 |
| Ti | LL.3 | 3.92 ± 0.77 | 15.23 ± 1.18 | 59.2 ± 8.05 | |
| Zn | W1.4 | 0.07 ± 0.04 | 0.38 ± 0.07 | 1.06 ± 0.33 | |
| Cu | W2.3 | 9.17 ± 0.41 | 12.45 ± 0.78 | 20.10 ± 3.69 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hurtado-Gallego, J.; Leganés, F.; Rosal, R.; Fernández-Piñas, F. Use of Cyanobacterial Luminescent Bioreporters to Report on the Environmental Impact of Metallic Nanoparticles. Sensors 2019, 19, 3597. https://doi.org/10.3390/s19163597
Hurtado-Gallego J, Leganés F, Rosal R, Fernández-Piñas F. Use of Cyanobacterial Luminescent Bioreporters to Report on the Environmental Impact of Metallic Nanoparticles. Sensors. 2019; 19(16):3597. https://doi.org/10.3390/s19163597
Chicago/Turabian StyleHurtado-Gallego, Jara, Francisco Leganés, Roberto Rosal, and Francisca Fernández-Piñas. 2019. "Use of Cyanobacterial Luminescent Bioreporters to Report on the Environmental Impact of Metallic Nanoparticles" Sensors 19, no. 16: 3597. https://doi.org/10.3390/s19163597
APA StyleHurtado-Gallego, J., Leganés, F., Rosal, R., & Fernández-Piñas, F. (2019). Use of Cyanobacterial Luminescent Bioreporters to Report on the Environmental Impact of Metallic Nanoparticles. Sensors, 19(16), 3597. https://doi.org/10.3390/s19163597