Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus

 , , , and
, , , and
Abstract
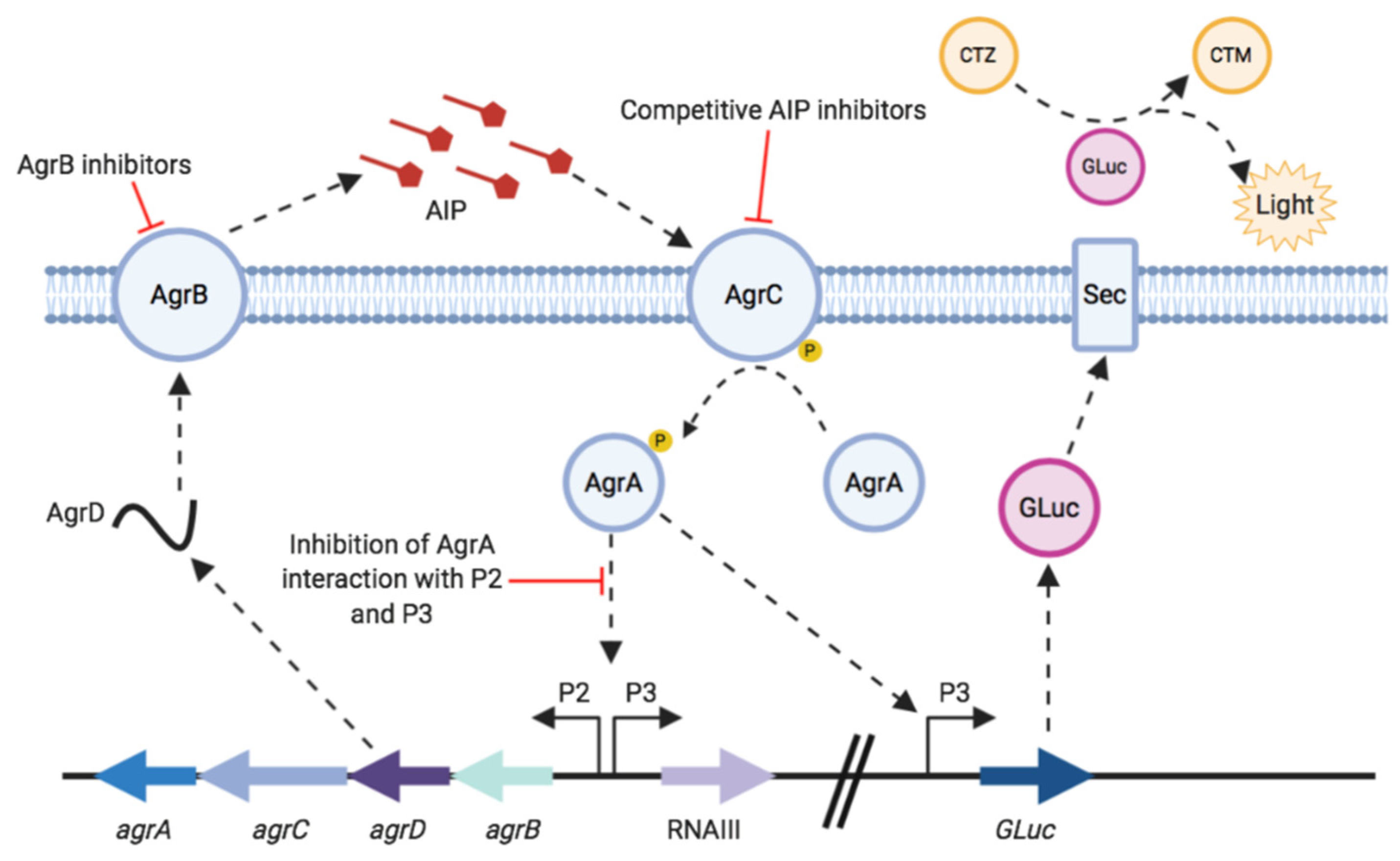
:1. Introduction
2. Materials and Methods
2.1. Construction of agrP3 Reporter S. aureus Strains
2.2. Measuring agrP3 Promoter Activity Using GAUSSIA Luciferase Bioluminescence
3. Results
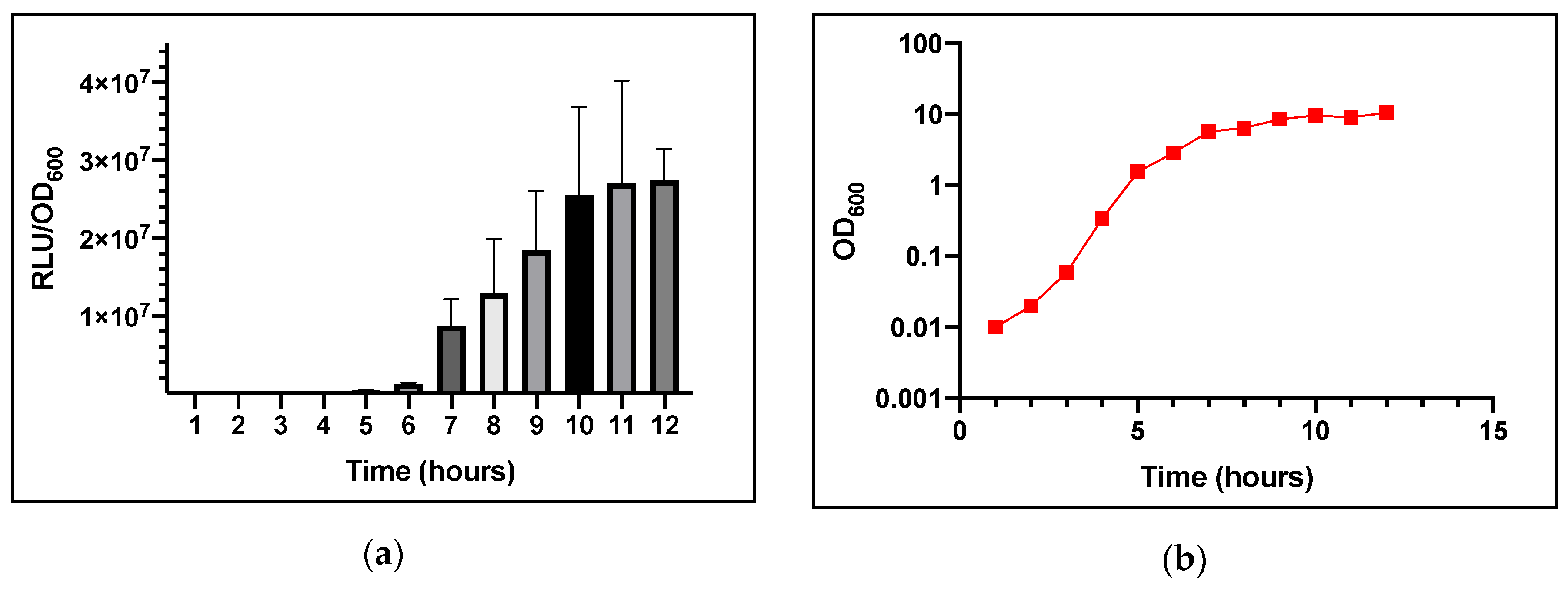
3.1. Induction of agr with Respect to Growth Phase
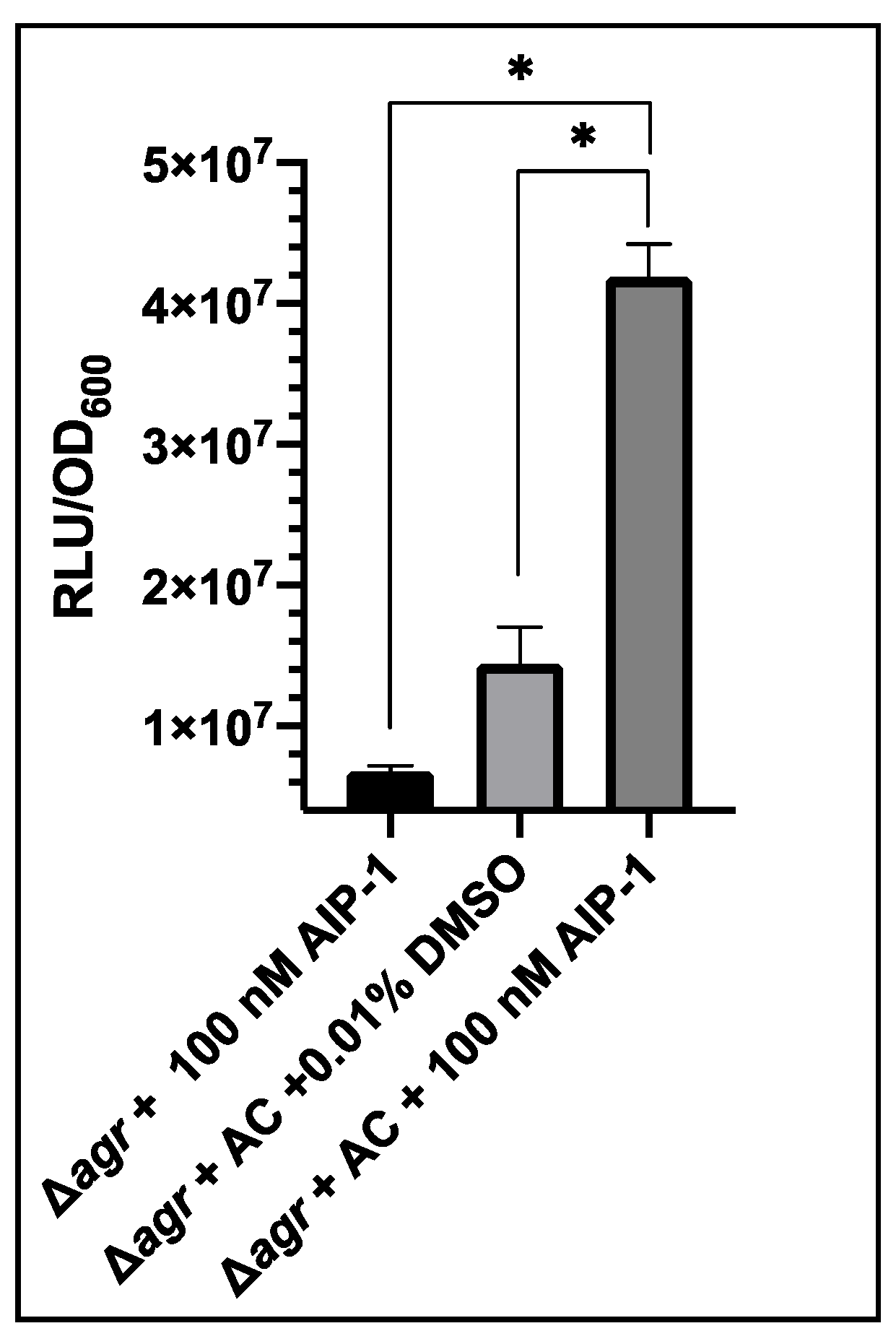
3.2. Activation of USA300 Δagr P3-GLuc pSKermP2 agrAC with Synthetic AIP-1
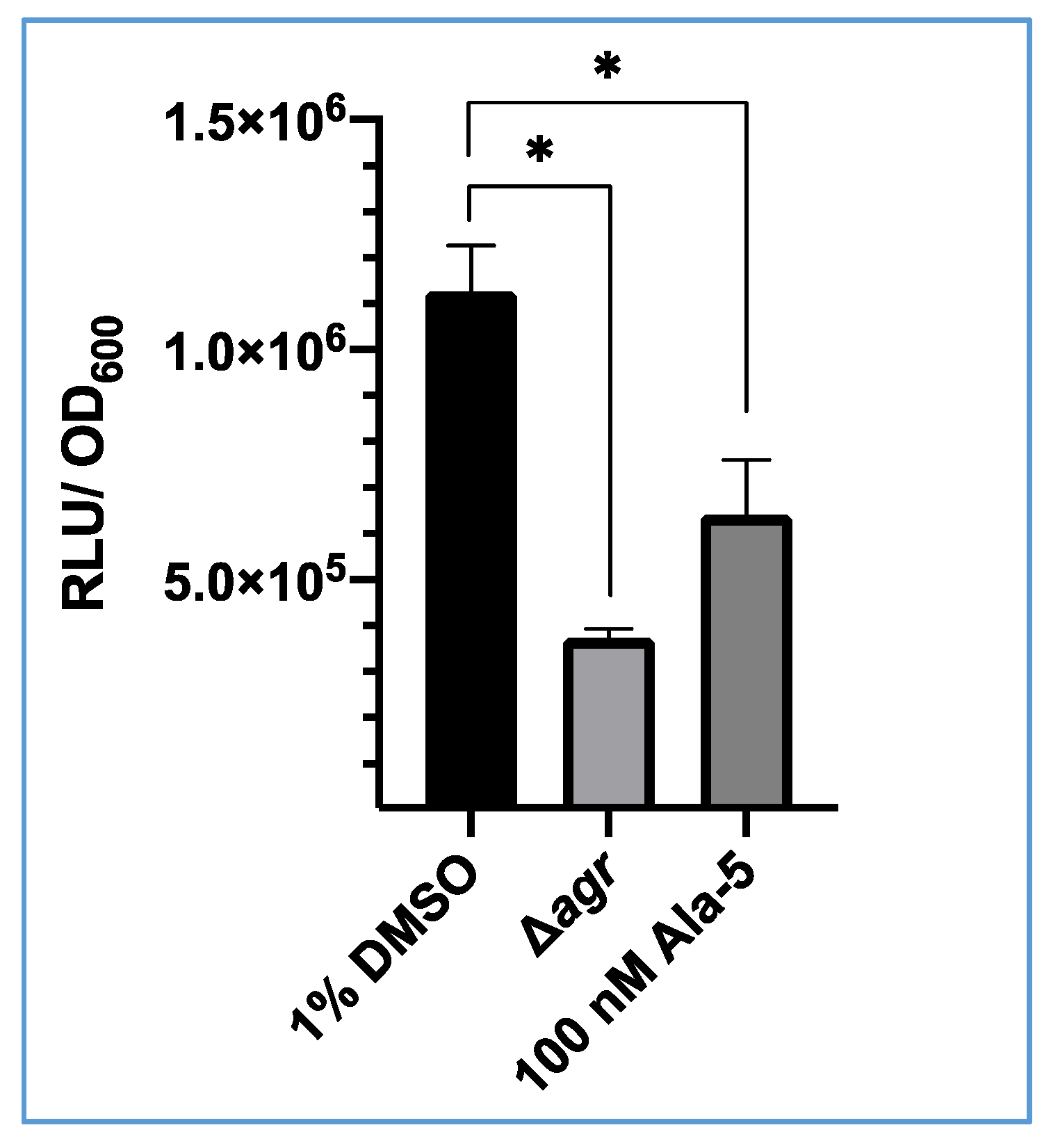
3.3. Inhibition of S. aureus USA300 agrP3 by an AgrC Antagonist (Ala5)AIP-1
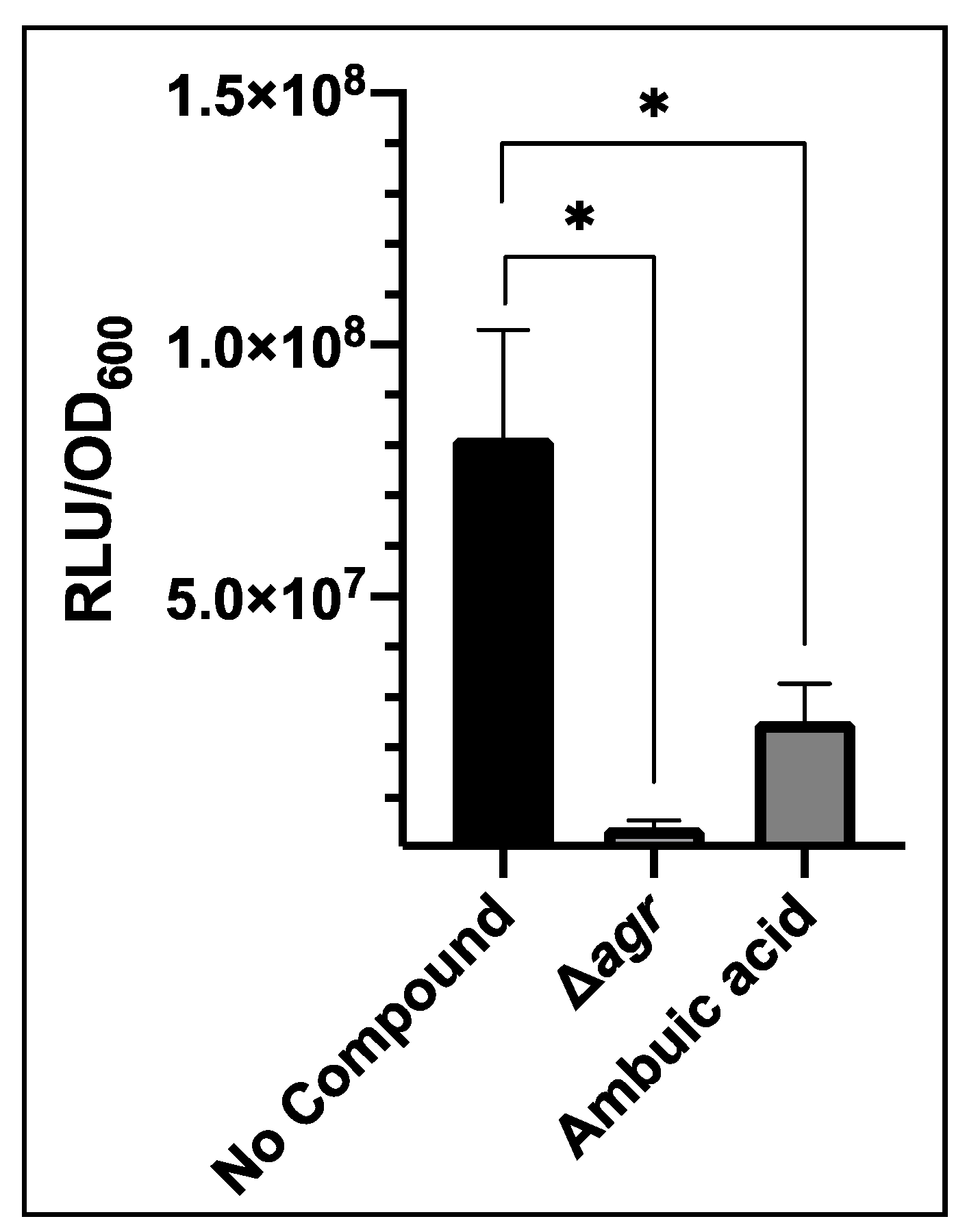
3.4. Evaluation of an AgrB Inhibitor with USA300 agrP3-GLuc
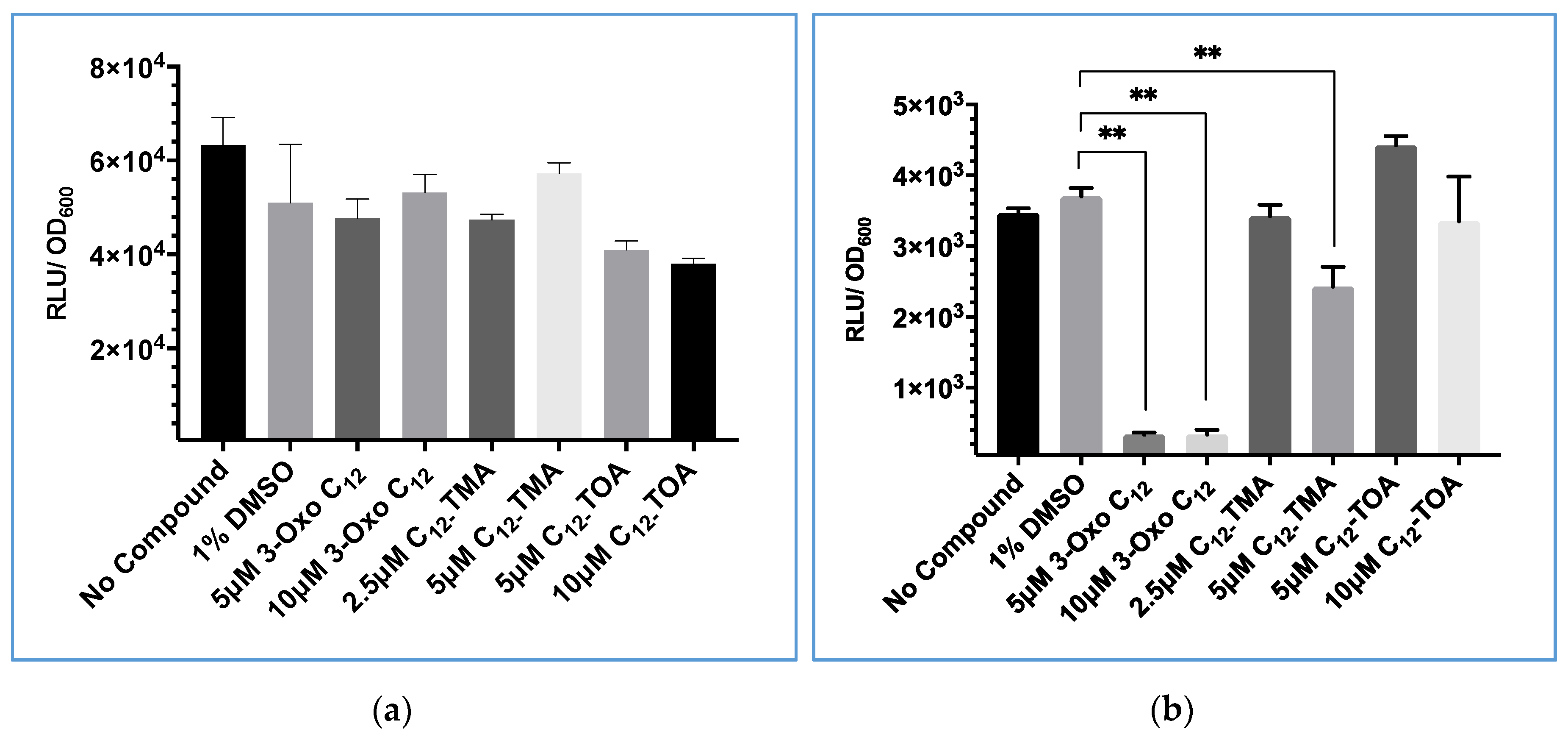
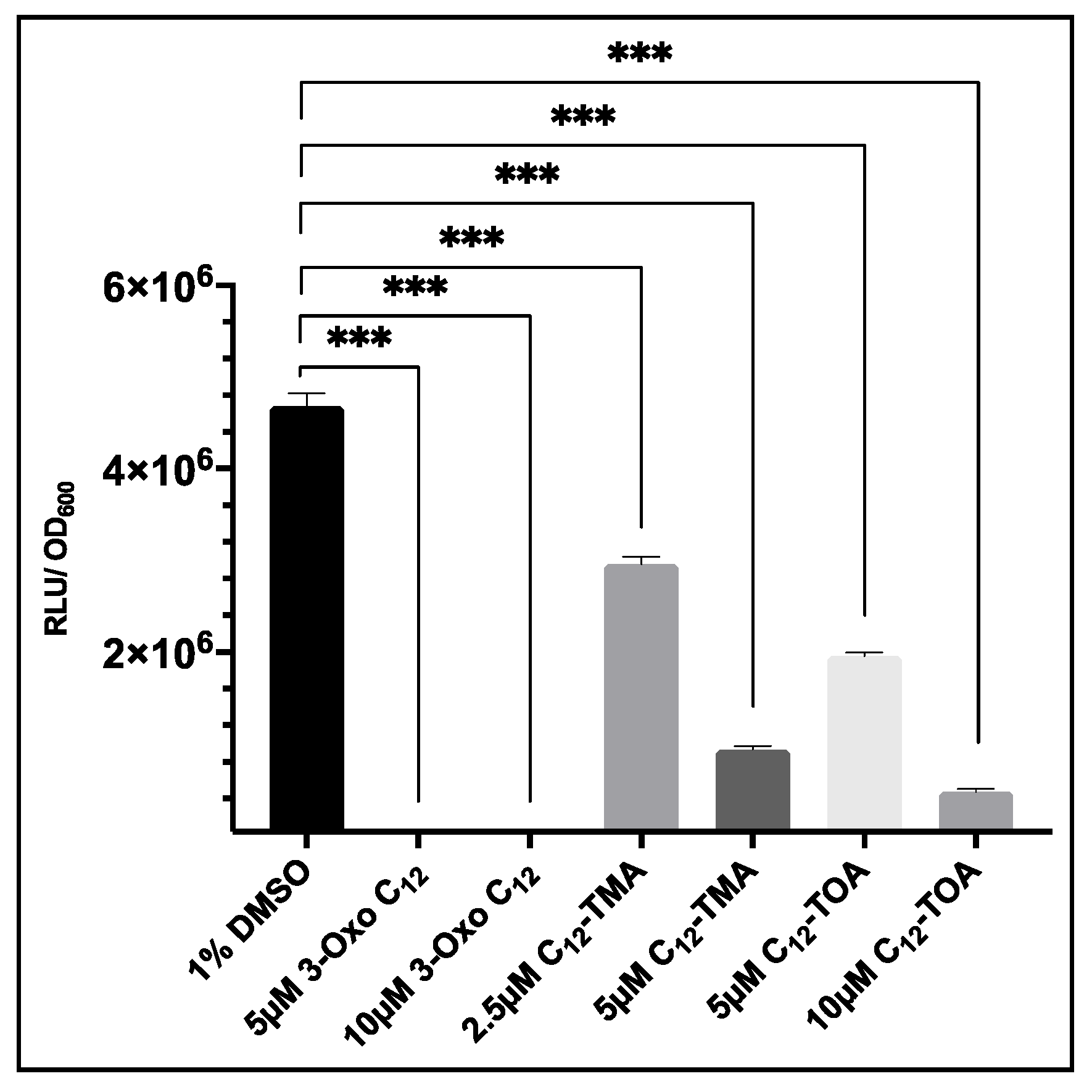
3.5. 3-Oxo-C12-HSL Has Differential Effects on GLuc and Lux Bioluminescence
3.6. Evaluation of USA300 agrP3 GLuc as a Screen for agr Inhibitors Related to 3-oxo-C12-HSL
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184–193. [Google Scholar] [CrossRef]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.H.; Khan, B.A.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-independent target gene control by the agr quorum-sensing system: Insight into the evolution of virulence regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boles, B.R.; Horswill, A.R. Agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Quorum-sensing control in Staphylococci—A target for antimicrobial drug therapy? FEMS Microbiol. Lett. 2004, 241, 135–141. [Google Scholar] [CrossRef]
- Gray, B.; Hall, P.; Gresham, H. Targeting agr- and agr-Like quorum sensing systems for development of common therapeutics to treat multiple gram-positive bacterial infections. Sensors 2013, 13, 5130–5166. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Projan, S.J.; Kornblum, J.; Ross, H.F.; Ji, G.; Kreiswirth, B.; Vandenesch, F.; Moghazeh, S. The agr P2 operon: An autocatalytic sensory transduction system in Staphylococcus aureus. Mol. Gen. Genet. 1995, 248, 446–458. [Google Scholar] [CrossRef]
- Novick, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef]
- Sloan, T.J.; Murray, E.; Yokoyama, M.; Massey, R.C.; Chan, W.C.; Bonev, B.B.; Williams, P. Timing is Everything: Impact of Naturally Occurring Staphylococcus aureus AgrC Cytoplasmic Domain Adaptive Mutations on Autoinduction. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desouky, S.E.; Nishiguchi, K.; Zendo, T.; Igarashi, Y.; Williams, P.; Sonomoto, K.; Nakayama, J. High-throughput screening of inhibitors targeting Agr/Fsr quorum sensing in Staphylococcus aureus and Enterococcus faecalis. Biosci. Biotechnol. Biochem. 2013, 77, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.P.; Williams, P.; Chan, W.C. Attenuating Staphylococcus aureus virulence gene regulation: A medicinal chemistry perspective. J. Med. Chem. 2013, 56, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.S., III; Jin, R.; Novick, R.P. Transient interference with Staphylococcal Quorum Sensing blocks abscess formation. Proc. Natl. Acad. Sci. USA 2005, 102, 1691–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, D.A.; Parlet, C.P.; Crosby, H.A.; Malone, C.L.; Heilmann, K.P.; Horswill, A.R.; Cech, N.B. Signal Biosynthesis Inhibition with Ambuic Acid as a Strategy To Target Antibiotic-Resistant Infections. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Daly, S.M.; Elmore, B.O.; Kavanaugh, J.S.; Triplett, K.D.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Crosby, H.A.; Femling, J.K.; Cech, N.B.; et al. Omega-Hydroxyemodin limits staphylococcus aureus quorum sensing-mediated pathogenesis and inflammation. Antimicrob. Agents Chemother. 2015, 59, 2223–2235. [Google Scholar] [CrossRef] [Green Version]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef]
- Kavanaugh, J.S.; Horswill, A.R. Impact of Environmental Cues on Staphylococcal Quorum Sensing and Biofilm Development. J. Biol. Chem. 2016, 291, 12556–12564. [Google Scholar] [CrossRef] [Green Version]
- Alksne, L.E.; Projan, S.J. Bacterial virulence as a target for antimicrobial chemotherapy. Curr. Opin. Biotechnol. 2000, 11, 625–636. [Google Scholar] [CrossRef]
- Qazi, S.; Middleton, B.; Muharram, S.H.; Cockayne, A.; Hill, P.; O’Shea, P.; Chhabra, S.R.; Cámara, M.; Williams, P. N-acylhomoserine lactones antagonize virulence gene expression and quorum sensing in Staphylococcus aureus. Infect. Immun. 2006, 74, 910–919. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, k.; Vidugiris, G.; Machleidt, T.; et al. Engineered luciferase reporter from a deep sea shrimp utilizing a novel imidazopyrazinone substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Shi, P.; Song, W.; Bi, S. Chemiluminescence and Bioluminescence Imaging for Biosensing and Therapy: In Vitro and In Vivo Perspectives. Theranostics 2019, 9, 4047–4065. [Google Scholar] [CrossRef] [PubMed]
- England, C.G.; Ehlerding, E.B.; Cai, W. NanoLuc: A Small Luciferase Is Brightening Up the Field of Bioluminescence. Bioconjug. Chem. 2016, 27, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Maguire, C.A.; Deliolanis, N.C.; Pike, L.; Niers, J.M.; Tjon-Kon-Fat, L.A.; Sena-Esteves, M.; Tannous, B.A. Gaussia luciferase variant for high-throughput functional screening applications. Anal. Chem. 2009, 81, 7102–7106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathnayaka, T.; Tawa, M.; Sohya, S.; Yohda, M.; Kuroda, Y. Biophysical characterization of highly active recombinant Gaussia luciferase expressed in Escherichia coli. Biochim. Biophys. Acta 2010, 1804, 1902–1907. [Google Scholar] [CrossRef]
- Wu, N.; Rathnayaka, T.; Kuroda, Y. Bacterial expression and re-engineering of Gaussia princeps luciferase and its use as a reporter protein. Biochim. Biophys. Acta 2015, 1854 Pt A, 1392–1399. [Google Scholar] [CrossRef]
- Wiles, S.; Ferguson, K.; Stefanidou, M.; Young, D.B.; Robertson, B.D. Alternative luciferase for monitoring bacterial cells under adverse conditions. Appl. Environ. Microbiol. 2005, 71, 3427–3432. [Google Scholar] [CrossRef] [Green Version]
- Wille, T.; Blank, K.; Schmidt, C.; Vogt, V.; Gerlach, R.G. Gaussia princeps luciferase as a reporter for transcriptional activity, protein secretion, and protein-protein interactions in Salmonella enterica serovar typhimurium. Appl. Environ. Microbiol. 2012, 78, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Tannous, B.A. Gaussia luciferase reporter assay for monitoring biological processes in culture and in vivo. Nat. Protoc. 2009, 4, 582–591. [Google Scholar] [CrossRef]
- Wurdinger, T.; Badr, C.; Pike, L.; de Kleine, R.; Weissleder, R.; Breakefield, X.O.; Tannous, B.A. A secreted luciferase for ex vivo monitoring of in vivo processes. Nat. Methods 2008, 5, 171–173. [Google Scholar] [CrossRef] [Green Version]
- Perehinec, T.M.; Qazi, S.N.; Gaddipati, S.R.; Salisbury, V.; Rees, C.E.; Hill, P.J. Construction and evaluation of multisite recombinatorial (Gateway) cloning vectors for Gram-positive bacteria. BMC Mol. Biol. 2007, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novick, R. Properties of a cryptic high-frequency transducing phage in Staphylococcus aureus. Virology 1967, 33, 155–166. [Google Scholar] [CrossRef]
- Lei, M.G.; Cue, D.; Alba, J.; Junecko, J.; Graham, J.W.; Lee, C.Y. A single copy integration vector that integrates at an engineered site on the Staphylococcus aureus chromosome. BMC Res. Notes 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fey, P.D.; Endres, J.L.; Yajjala, V.K.; Widhelm, T.J.; Boissy, R.J.; Bose, J.L.; Bayles, K.W. A genetic resource for rapid and comprehensive phenotype screening of nonessential Staphylococcus aureus genes. Mbio 2013, 4, e00537-12. [Google Scholar] [CrossRef] [Green Version]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: Establishing a national database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef] [Green Version]
- Jensen, R.O.; Winzer, K.; Clarke, S.R.; Chan, W.C.; Williams, P. Differential recognition of Staphylococcus aureus quorum-sensing signals depends on both extracellular loops 1 and 2 of the transmembrane sensor AgrC. J. Mol. Biol. 2008, 381, 300–309. [Google Scholar] [CrossRef]
- Monk, I.R.; Shah, I.M.; Xu, M.; Tan, M.W.; Foster, T.J. Transforming the untransformable: Application of direct transformation to manipulate genetically Staphylococcus aureus and Staphylococcus epidermidis. MBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Wider, D.; Picard, D. Secreted dual reporter assay with Gaussia luciferase and the red fluorescent protein mCherry. PLoS ONE 2017, 12, e0189403. [Google Scholar] [CrossRef] [Green Version]
- McDowell, P.; Affas, Z.; Reynolds, C.; Holden, M.T.; Wood, S.J.; Saint, S.; Cockayne, A.; Hill, P.J.; Dodd, C.E.; Bycroft, B.W.; et al. Structure, activity and evolution of the group I thiolactone peptide quorum-sensing system of Staphylococcus aureus. Mol. Microbiol. 2001, 41, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Murray, E.J.; Crowley, R.C.; Truman, A.; Clarke, S.R.; Cottam, J.A.; Jadhav, G.P.; Steele, V.R.; O’Shea, P.; Lindholm, C.; Cockayne, A.; et al. Targeting Staphylococcus aureus quorum sensing with nonpeptidic small molecule inhibitors. J. Med. Chem. 2014, 57, 2813–2819. [Google Scholar] [CrossRef]
- Baldry, M.; Kitir, B.; Frokiaer, H.; Christensen, S.B.; Taverne, N.; Meijerink, M.; Franzyk, H.; Olsen, C.A.; Wells, J.M.; Ingmer, H. The agr Inhibitors Solonamide B and Analogues Alter Immune Responses to Staphylococccus aureus but Do Not Exhibit Adverse Effects on Immune Cell Functions. PLoS ONE 2016, 11, e0145618. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.; Todd, D.A.; Velazquez, J.V.; Cech, N.B.; Sonenshein, A.L. CodY-mediated regulation of the Staphylococcus aureus Agr system integrates nutritional and population density signals. J. Bacteriol. 2014, 196, 1184–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Blinn, C.; McLeod, S.M.; Wiseman, J.W.; Newman, J.V.; Fisher, S.L.; Walkup, G.K. Secreted Gaussia princeps luciferase as a reporter of Escherichia coli replication in a mouse tissue cage model of infection. PLoS ONE 2014, 9, e90382. [Google Scholar] [CrossRef] [PubMed]
- Mayville, P.; Ji, G.; Beavis, R.; Yang, H.; Goger, M.; Novick, R.P.; Muir, T.W. Structure-activity analysis of synthetic autoinducing thiolactone peptides from Staphylococcus aureus responsible for virulence. Proc. Natl. Acad. Sci. USA 1999, 96, 1218–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference |
|---|---|---|
| S. aureus 8325-4 | Derivative of S. aureus NCTC 8325 | [32] |
| S. aureus RN4220 attB2 | Cloning intermediate | [33] |
| S. aureus USA300 JE2 | Plasmid-cured derivative of the CA-MRSA strain USA300 LAC | [34,35] |
| S. aureus RN4220 pXyl/tet: SpA leader: GLuc | Constitutively expressing SpA leader:GLuc | This study |
| S. aureus RN4220 Pxyl/tet:::luxABCDE | Constitutively bioluminescent strain | [20] |
| S. aureus USA300 agrP3 Gluc | USA300 with chromosomal integration of SpA leader and GLuc under the control of agrP3 | This study |
| S. aureus USA300 Δagr P3 Gluc | USA300 Δagr with chromosomal integration of SpA leader and GLuc under the control of agrP3. | This study |
| S. aureus ROJ143 | ROJ143 carrying plasmid pSKermP2 agrC1 agrA | [36] |
| S. aureus USA300 Δagr P3 Gluc pSKermP2 agrC1 agrA | USA300 Δagr P3 Gluc carrying pSKermP2 agrC1 agrA | This study |
| E. coli DC10B | Cloning intermediate dc | [37] |
| Primer | Sequence | Function | Reference |
|---|---|---|---|
| CT12 (F) | CTAGTAGGAGGAAAAACATATGATGACTTTACA | Amplification of SpA:GLuc | This study |
| CT13 (R) | ATTTGTCGACCTCAGGAGAGCGTTCACC | ||
| CT14 (F) | AGTGAATTCCCGGGGATCCGACACGTCGACCCTCACTG | Amplification of agrP3 | This study |
| CT15 (R) | CTCCTACTAGCCATCACATCTCTGTGATCTAG | ||
| CT16 (F) | GATGTGATGGCTAGTAGGAGGAAAAACATATGATG | Amplification of SpA:GLuc | This study |
| CT17 (R) | TCGATAAGCTTGGCTGCAGGATTTGTCGACCTCAGGAG | ||
| OU9R10 | CATACTACATATCAACGAAATCAG | Forward primer at RN4220 attB2 integration site | [33] |
| SCV4 | ACCCAGTTTGTAATTCCAGGAG | Reverse primer at 5′ end of pLL102 to RN4220 attB2 | [33] |
| SCV8 | GCACATAATTGCTCACAGCCA | Forward primer at 3′ end of pLL102 to RN4220 attB2 | [33] |
| OU9R7 | ATGGGTGGTAAAACACAAATTTC | Reverse primer at RN4220 attB2 integration site | [33] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blower, I.; Tong, C.; Sun, X.; Murray, E.; Luckett, J.; Chan, W.; Williams, P.; Hill, P. Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus. Sensors 2020, 20, 4305. https://doi.org/10.3390/s20154305
Blower I, Tong C, Sun X, Murray E, Luckett J, Chan W, Williams P, Hill P. Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus. Sensors. 2020; 20(15):4305. https://doi.org/10.3390/s20154305
Chicago/Turabian StyleBlower, Isobel, Carmen Tong, Xiaohui Sun, Ewan Murray, Jeni Luckett, Weng Chan, Paul Williams, and Philip Hill. 2020. "Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus" Sensors 20, no. 15: 4305. https://doi.org/10.3390/s20154305
APA StyleBlower, I., Tong, C., Sun, X., Murray, E., Luckett, J., Chan, W., Williams, P., & Hill, P. (2020). Gaussia Luciferase as a Reporter for Quorum Sensing in Staphylococcus aureus. Sensors, 20(15), 4305. https://doi.org/10.3390/s20154305