
Decoding Brain Responses to Names and Voices across Different Vigilance States

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Stimulation
2.3. EEG Acquisition and Data Pre-Processing
2.4. Decoding and Statistical Analyses
3. Results
3.1. Decoding of the Main Effect of VOICE Familiarity
3.2. Voice Familiarity across the Brain—Topographical Patterns
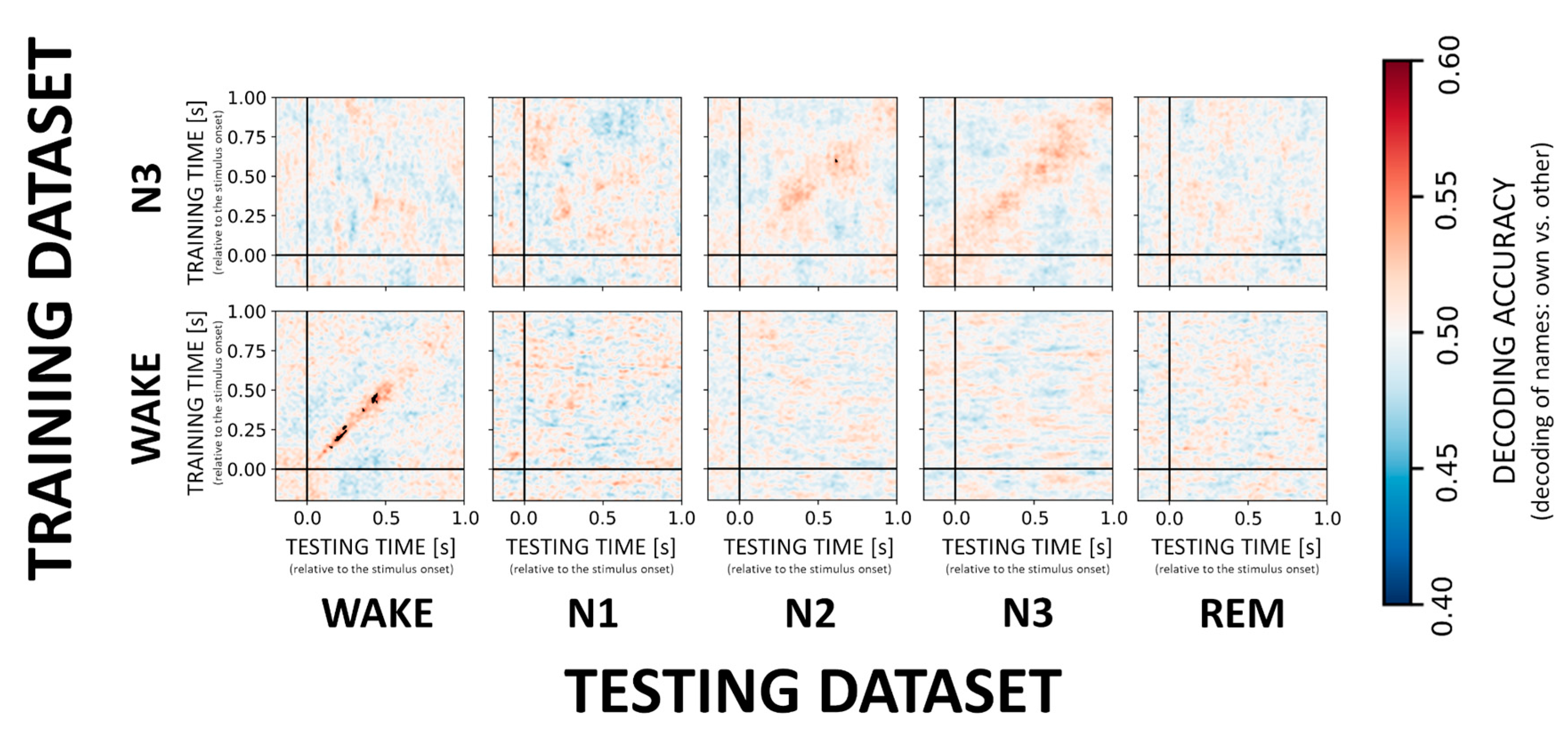
3.3. Decoding of the NAME Effect
3.4. NAME Effect across the Brain—Topographical Patterns
4. Discussion
Supplementary Materials
Author Contributions
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blume, C.; Del Giudice, R.; Wislowska, M.; Heib, D.P.; Schabus, M. Standing sentinel during human sleep: Continued evaluation of environmental stimuli in the absence of consciousness. NeuroImage 2018, 178, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Perrin, F.; Garcia-Larrea, L.; Mauguiere, F.; Bastuji, H. A differential brain response to the subject’s own name persists during sleep. Clin. Neurophysiol. 1999, 110, 2153–2164. [Google Scholar] [CrossRef]
- Blume, C.; del Giudice, R.; Lechinger, J.; Wislowska, M.; Heib, D.P.; Hoedlmoser, K.; Schabus, M. Preferential processing of emotionally and self-relevant stimuli persists in unconscious N2 sleep. Brain Lang. 2017, 167, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouider, S.; Andrillon, T.; Barbosa, L.S.; Goupil, L.; Bekinschtein, T.A. Inducing Task-Relevant Responses to Speech in the Sleeping Brain. Curr. Biol. 2014, 24, 2208–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legendre, G.; Andrillon, T.; Koroma, M.; Kouider, S. Sleepers track informative speech in a multitalker environment. Nat. Hum. Behav. 2019, 3, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.; Sitt, J.D.; King, J.-R.; Elbaz, M.; Azizi, L.; Buiatti, M.; Naccache, L.; van Wassenhove, V.; Dehaene, S. Disruption of hierarchical predictive coding during sleep. Proc. Natl. Acad. Sci. USA 2015, 112, E1353–E1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strauss, M.; Dehaene, S. Detection of arithmetic violations during sleep. Sleep 2019, 42. [Google Scholar] [CrossRef] [PubMed]
- Atienza, M.; Cantero, J.L.; Escera, C. Auditory information processing during human sleep as revealed by event-related brain potentials. Clin. Neurophysiol. 2001, 112, 2031–2045. [Google Scholar] [CrossRef]
- King, J.; Faugeras, F.; Gramfort, A.; Schurger, A.; El Karoui, I.; Sitt, J.; Rohaut, B.; Wacongne, C.; Labyt, E.; Bekinschtein, T.; et al. Single-trial decoding of auditory novelty responses facilitates the detection of residual consciousness. NeuroImage 2013, 83, 726–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, J.-R.; Dehaene, S. Characterizing the dynamics of mental representations: The temporal generalization method. Trends Cogn. Sci. 2014, 18, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boly, M.; Perlbarg, V.; Marrelec, G.; Schabus, M.; Laureys, S.; Doyon, J.; Pélégrini-Issac, M.; Maquet, P.; Benali, H. Hierarchical clustering of brain activity during human nonrapid eye movement sleep. Proc. Natl. Acad. Sci. USA 2012, 109, 5856–5861. [Google Scholar] [CrossRef] [Green Version]
- Rechtschaffen, A.; Hauri, P.; Zeitlin, M. Auditory Awakening Thresholds in Rem and Nrem Sleep Stages. Percept. Mot. Ski. 1966, 22, 927–942. [Google Scholar] [CrossRef] [PubMed]
- Andrillon, T.; Poulsen, A.T.; Hansen, L.K.; Léger, D.; Kouider, S. Neural Markers of Responsiveness to the Environment in Human Sleep. J. Neurosci. 2016, 36, 6583–6596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruby, P.; Caclin, A.; Boulet, S.; Delpuech, C.; Morlet, D. Odd Sound Processing in the Sleeping Brain. J. Cogn. Neurosci. 2008, 20, 296–311. [Google Scholar] [CrossRef] [PubMed]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2010, 2011, 1–9. [Google Scholar] [CrossRef]
- Anderer, P.; Gruber, G.; Paraptics, S.; Woertz, M.; Miazhynskaia, T.; Klösch, G.; Danker-Hopfe, H. An E-health solution for automatic sleep classification according to Rechtschaffen and Kales: Validation study of the Somnolyzer 24 x 7 utilizing the Siesta database. Neuropsychobiology 2005, 51, 115–133. [Google Scholar] [CrossRef]
- Iber, C.; Ancoli-Israel, S.; Chesson, A.; Quan, S. American Academy of Sleep Medicine. In The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications; American Academy of Sleep Medicine: Westchester, IL, USA, 2007. [Google Scholar]
- Gramfort, A.; Luessi, M.; Larson, E.; Engemann, D.A.; Strohmeier, D.; Brodbeck, C.; Parkkonen, L.; Hämäläinen, M.S. MNE software for processing MEG and EEG data. NeuroImage 2014, 86, 446–460. [Google Scholar] [CrossRef] [Green Version]
- Varoquaux, G.; Raamana, P.R.; Engemann, D.A.; Hoyos-Idrobo, A.; Schwartz, Y.; Thirion, B. Assessing and tuning brain decoders: Cross-validation, caveats, and guidelines. NeuroImage 2017, 145, 166–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haufe, S.; Meinecke, F.; Görgen, K.; Dähne, S.; Haynes, J.-D.; Blankertz, B.; Bießmann, F. On the interpretation of weight vectors of linear models in multivariate neuroimaging. NeuroImage 2014, 87, 96–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wielek, T.; Blume, C.; Wislowska, M.; del Giudice, R.; Schabus, M. Decoding Brain Responses to Names and Voices across Different Vigilance States. Sensors 2021, 21, 3393. https://doi.org/10.3390/s21103393
Wielek T, Blume C, Wislowska M, del Giudice R, Schabus M. Decoding Brain Responses to Names and Voices across Different Vigilance States. Sensors. 2021; 21(10):3393. https://doi.org/10.3390/s21103393
Chicago/Turabian StyleWielek, Tomasz, Christine Blume, Malgorzata Wislowska, Renata del Giudice, and Manuel Schabus. 2021. "Decoding Brain Responses to Names and Voices across Different Vigilance States" Sensors 21, no. 10: 3393. https://doi.org/10.3390/s21103393
APA StyleWielek, T., Blume, C., Wislowska, M., del Giudice, R., & Schabus, M. (2021). Decoding Brain Responses to Names and Voices across Different Vigilance States. Sensors, 21(10), 3393. https://doi.org/10.3390/s21103393