
A Membrane Filter-Assisted Mammalian Cell-Based Biosensor Enabling 3D Culture and Pathogen Detection


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Analytical Components
2.3. Analytical Procedures
3. Results and Discussion
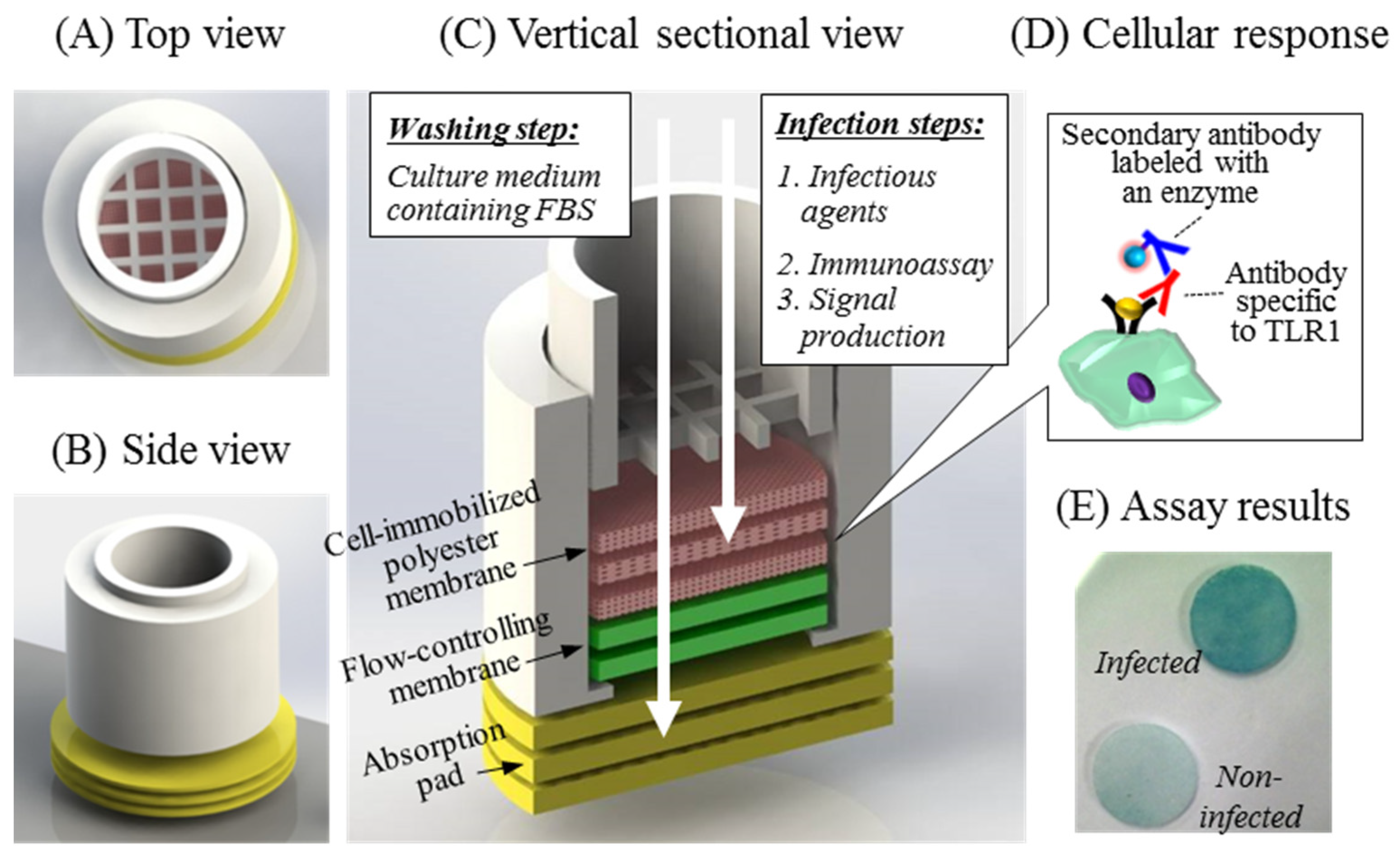
3.1. Analytical Concept
3.2. Fabrication of 3D Cell Culture Membrane
3.3. Characterization of the Cell-Based Biosensor
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Banerjee, P.; Bhunia, A.K. Mammalian cell-based biosensors for pathogens and toxins. Trends Biotechnol. 2009, 27, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xu, G.; Qin, L.; Xu, Y.; Li, Y.; Li, R. Cell-based biosensors and its application in biomedicine. Sens. Actuators B Chem. 2005, 108, 576–584. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, C.; Cai, H.; Hu, N.; Zhou, J.; Wang, P. Cell-Based Biosensors and Their Application in Biomedicine. Chem. Rev. 2014, 114, 6423–6461. [Google Scholar] [CrossRef] [PubMed]
- Bawolak, M.-T.; Gera, L.; Morissette, G.; Bouthillier, J.; Stewart, J.M.; Gobeil, L.-A.; Lodge, R.; Adam, A.; Marceau, F. Fluorescent Ligands of the Bradykinin B1 Receptors: Pharmacologic Characterization and Application to the Study of Agonist-Induced Receptor Translocation and Cell Surface Receptor Expression. J. Pharmacol. Exp. Ther. 2009, 329, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef]
- Cho, I.-H.; Jeon, J.-W.; Paek, S.-H.; Kim, D.-H.; Shin, H.-S.; Ha, U.-H.; Seo, S.-K.; Paek, S.-H. Toll-Like Receptor-Based Immuno-Analysis of Pathogenic Microorganisms. Anal. Chem. 2012, 84, 9713–9720. [Google Scholar] [CrossRef]
- Wolf, M.; Zimmermann, M.; Delamarche, E.; Hunziker, P. Screening cell surface receptors using micromosaic immunoassays. Biomed. Microdevices 2007, 9, 135–141. [Google Scholar] [CrossRef]
- Kohl, T.O.; Ascoli, C.A. Direct and Indirect Cell-Based Enzyme-Linked Immunosorbent Assay. Cold Spring Harb. Protoc. 2017, 2017. [Google Scholar] [CrossRef]
- Lu, X.; Ye, Y.; Zhang, Y.; Sun, X. Current research progress of mammalian cell-based biosensors on the detection of foodborne pathogens and toxins. Crit. Rev. Food Sci. Nutr. 2020. [Google Scholar] [CrossRef]
- Hertzberg, R.P.; Pope, A.J. High-throughput screening: New technology for the 21st century. Curr. Opin. Chem. Biol. 2000, 4, 445–451. [Google Scholar] [CrossRef]
- Slaughter, G.E.; Hobson, R. An impedimetric biosensor based on PC 12 cells for the monitoring of exogenous agents. Biosens. Bioelectron. 2009, 24, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Kloss, D.; Fischer, M.; Rothermel, A.; Simon, J.C.; Robitzki, A.A. Drug testing on 3D in vitro tissues trapped on a microcavity chip. Lab Chip 2008, 8, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Ye, X.; Qin, L.; Xu, Y.; Li, Y.; Li, R.; Wang, P. Cell-based biosensors based on light-addressable potentiometric sensors for single cell monitoring. Biosens. Bioelectron. 2005, 20, 1757–1763. [Google Scholar] [CrossRef]
- Gao, G.; Qian, J.; Fang, D.; Yu, Y.; Zhi, J. Development of a mediated whole cell-based electrochemical biosensor for joint toxicity assessment of multi-pollutants using a mixed microbial consortium. Anal. Chim. Acta 2016, 924, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Pollack, R.E.; Green, H.; Todaro, G.J. Growth control in cultured cells: Selection of sublines with increased sensitivity to contact inhibition and decreased tumor-producing ability. Proc. Natl. Acad. Sci. USA 1968, 60, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Todaro, G.J.; Lazar, G.K.; Green, H. The initiation of cell division in a contact-inhibited mammalian cell line. J. Cell. Comp. Physiol. 1965, 66, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Grinnell, F. Fibroblast biology in three-dimensional collagen matrices. Trends Cell Biol. 2003, 13, 264–269. [Google Scholar] [CrossRef]
- Grinnell, F.; Ho, C.-H.; Tamariz, E.; Lee, D.J.; Skuta, G. Dendritic Fibroblasts in Three-dimensional Collagen Matrices. Mol. Biol. Cell 2003, 14, 384–395. [Google Scholar] [CrossRef] [Green Version]
- Khetan, S.; Burdick, J.A. Patterning network structure to spatially control cellular remodeling and stem cell fate within 3-dimensional hydrogels. Biomaterials 2010, 31, 8228–8234. [Google Scholar] [CrossRef]
- Lee, S.-H.; Moon, J.J.; West, J.L. Three-dimensional micropatterning of bioactive hydrogels via two-photon laser scanning photolithography for guided 3D cell migration. Biomaterials 2008, 29, 2962–2968. [Google Scholar] [CrossRef] [Green Version]
- Huxley-Jones, J.; Robertson, D.L.; Boot-Handford, R.P. On the origins of the extracellular matrix in vertebrates. Matrix Biol. 2007, 26, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension—how 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125, 3015–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M. Degradation of Polyesters in Medical Applications; IntechOpen: London, UK, 2012. [Google Scholar]
- Khlebtsov, B.N.; Tumskiy, R.S.; Burov, A.M.; Pylaev, T.E.; Khlebtsov, N.G. Quantifying the Numbers of Gold Nanoparticles in the Test Zone of Lateral Flow Immunoassay Strips. ACS Appl. Nano Mater. 2019, 2, 5020–5028. [Google Scholar] [CrossRef] [Green Version]
- Koczula, K.M.; Gallotta, A. Lateral flow assays. Essays Biochem. 2016, 60, 111–120. [Google Scholar] [CrossRef]
- Moghadas, H.; Saidi, M.S.; Kashaninejad, N.; Nguyen, N.-T. A high-performance polydimethylsiloxane electrospun membrane for cell culture in lab-on-a-chip. Biomicrofluidics 2018, 12, 024117. [Google Scholar] [CrossRef]
- Jiang, D.; Ge, P.; Wang, L.; Jiang, H.; Yang, M.; Yuan, L.; Ge, Q.; Fang, W.; Ju, X. A novel electrochemical mast cell-based paper biosensor for the rapid detection of milk allergen casein. Biosens. Bioelectron. 2019, 130, 299–306. [Google Scholar] [CrossRef]
- Ng, K.; Gao, B.; Yong, K.W.; Li, Y.; Shi, M.; Zhao, X.; Li, Z.; Zhang, X.; Pingguan-Murphy, B.; Yang, H.; et al. Paper-based cell culture platform and its emerging biomedical applications. Mater. Today 2017, 20, 32–44. [Google Scholar] [CrossRef]
- Shin, H.S.; Ha, U.H. Up-regulation of bradykinin B2 receptor by Pseudomonas aeruginosa via the NF-κB pathway. Curr. Microbiol. 2011, 63, 138–144. [Google Scholar] [CrossRef]
- Zor, T.; Selinger, Z. Linearization of the Bradford Protein Assay Increases Its Sensitivity: Theoretical and Experimental Studies. Anal. Biochem. 1996, 236, 302–308. [Google Scholar] [CrossRef] [Green Version]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In Vitro Cultivation of Human Tumors: Establishment of Cell Lines Derived From a Series of Solid Tumors2. J. Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef]
- Jeon, J.-W.; Ha, U.-H.; Paek, S.-H. In Vitro Inflammation Inhibition Model Based on Semi-Continuous Toll-Like Receptor Biosensing. PLoS ONE 2014, 9, e105212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assoian, R.K. Anchorage-dependent Cell Cycle Progression. J. Cell Biol. 1997, 136, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Huang, J.; Yang, X.; Liu, B.; Zhang, W.; Huang, L.; Deng, F.; Ma, J.; Bai, Y.; Lu, R.; et al. Serum Starvation Induced Cell Cycle Synchronization Facilitates Human Somatic Cells Reprogramming. PLoS ONE 2012, 7, e28203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.H.; Nguyen, N.-T.; Chua, Y.C.; Kang, T.G. Oxygen plasma treatment for reducing hydrophobicity of a sealed polydimethylsiloxane microchannel. Biomicrofluidics 2010, 4, 032204. [Google Scholar] [CrossRef] [Green Version]
- Eddington, D.T.; Puccinelli, J.P.; Beebe, D.J. Thermal aging and reduced hydrophobic recovery of polydimethylsiloxane. Sens. Actuators B Chem. 2006, 114, 170–172. [Google Scholar] [CrossRef]
- Xiao, D.; Zhang, H.; Wirth, M. Chemical Modification of the Surface of Poly(dimethylsiloxane) by Atom-Transfer Radical Polymerization of Acrylamide. Langmuir 2002, 18, 9971–9976. [Google Scholar] [CrossRef]
- Vroman, L. Effect of absorbed proteins on the wettability of hydrophilic and hydrophobic solids. Nature 1962, 196, 476–477. [Google Scholar] [CrossRef]
- Yadav, G.D.; Thathagar, M.B. Esterification of maleic acid with ethanol over cation-exchange resin catalysts. React. Funct. Polym. 2002, 52, 99–110. [Google Scholar] [CrossRef]
- Ito, Y. Surface micropatterning to regulate cell functions. Biomaterials 1999, 20, 2333–2342. [Google Scholar] [CrossRef]
- Da Cunha, C.B.; Lopes, P.P.; Mayer, F.D.; Hoffmann, R. Assessment of Chemical and Mechanical Properties of Polymers Aiming to Replace the Stainless Steel in Distillation Column. Mater. Res. 2018, 21. [Google Scholar] [CrossRef]
- Marongiu, A.; Faravelli, T.; Bozzano, G.; Dente, M.; Ranzi, E. Thermal degradation of poly(vinyl chloride). J. Anal. Appl. Pyrolysis 2003, 70, 519–553. [Google Scholar] [CrossRef]
- Seo, S.-M.; Kim, S.-W.; Park, J.-N.; Cho, J.-H.; Kim, H.-S.; Paek, S.-H. A fluorescent immunosensor for high-sensitivity cardiac troponin I using a spatially-controlled polymeric, nano-scale tracer to prevent quenching. Biosens. Bioelectron. 2016, 83, 19–26. [Google Scholar] [CrossRef]
- Jiang, R.-D.; Shen, H.; Piao, Y.-J. The morphometrical analysis on the ultrastructure of A549 cells. Rom. J. Morphol. Embryol. 2010, 51, 663–667. [Google Scholar]
- Raspotnig, G.; Fauler, G.; Jantscher, A.; Windischhofer, W.; Schachl, K.; Leis, H.J. Colorimetric Determination of Cell Numbers by Janus Green Staining. Anal. Biochem. 1999, 275, 74–83. [Google Scholar] [CrossRef]
- Hennessy, E.J.; Parker, A.E.; O’Neill, L.A.J. Targeting Toll-like receptors: Emerging therapeutics? Nat. Rev. Drug Discov. 2010, 9, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Okano, T.; Yamada, N.; Okuhara, M.; Sakai, H.; Sakurai, Y. Mechanism of cell detachment from temperature-modulated, hydrophilic-hydrophobic polymer surfaces. Biomaterials 1995, 16, 297–303. [Google Scholar] [CrossRef]
- Weiss, L.; Blumenson, L.E. Dynamic adhesion and separation of cellsin vitro II. Interactions of cells with hydrophilic and hydrophobic surfaces. J. Cell. Physiol. 1967, 70, 23–32. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, I.-H.; Jeon, J.-W.; Choi, M.-J.; Cho, H.-M.; Lee, J.-S.; Kim, D.-H. A Membrane Filter-Assisted Mammalian Cell-Based Biosensor Enabling 3D Culture and Pathogen Detection. Sensors 2021, 21, 3042. https://doi.org/10.3390/s21093042
Cho I-H, Jeon J-W, Choi M-J, Cho H-M, Lee J-S, Kim D-H. A Membrane Filter-Assisted Mammalian Cell-Based Biosensor Enabling 3D Culture and Pathogen Detection. Sensors. 2021; 21(9):3042. https://doi.org/10.3390/s21093042
Chicago/Turabian StyleCho, Il-Hoon, Jin-Woo Jeon, Min-Ji Choi, Hyun-Mo Cho, Jong-Sung Lee, and Dong-Hyung Kim. 2021. "A Membrane Filter-Assisted Mammalian Cell-Based Biosensor Enabling 3D Culture and Pathogen Detection" Sensors 21, no. 9: 3042. https://doi.org/10.3390/s21093042