
Evaluation of a Binary Classification Approach to Detect Herbage Scarcity Based on Behavioral Responses of Grazing Dairy Cows



Abstract
:
1. Introduction
2. Materials and Methods
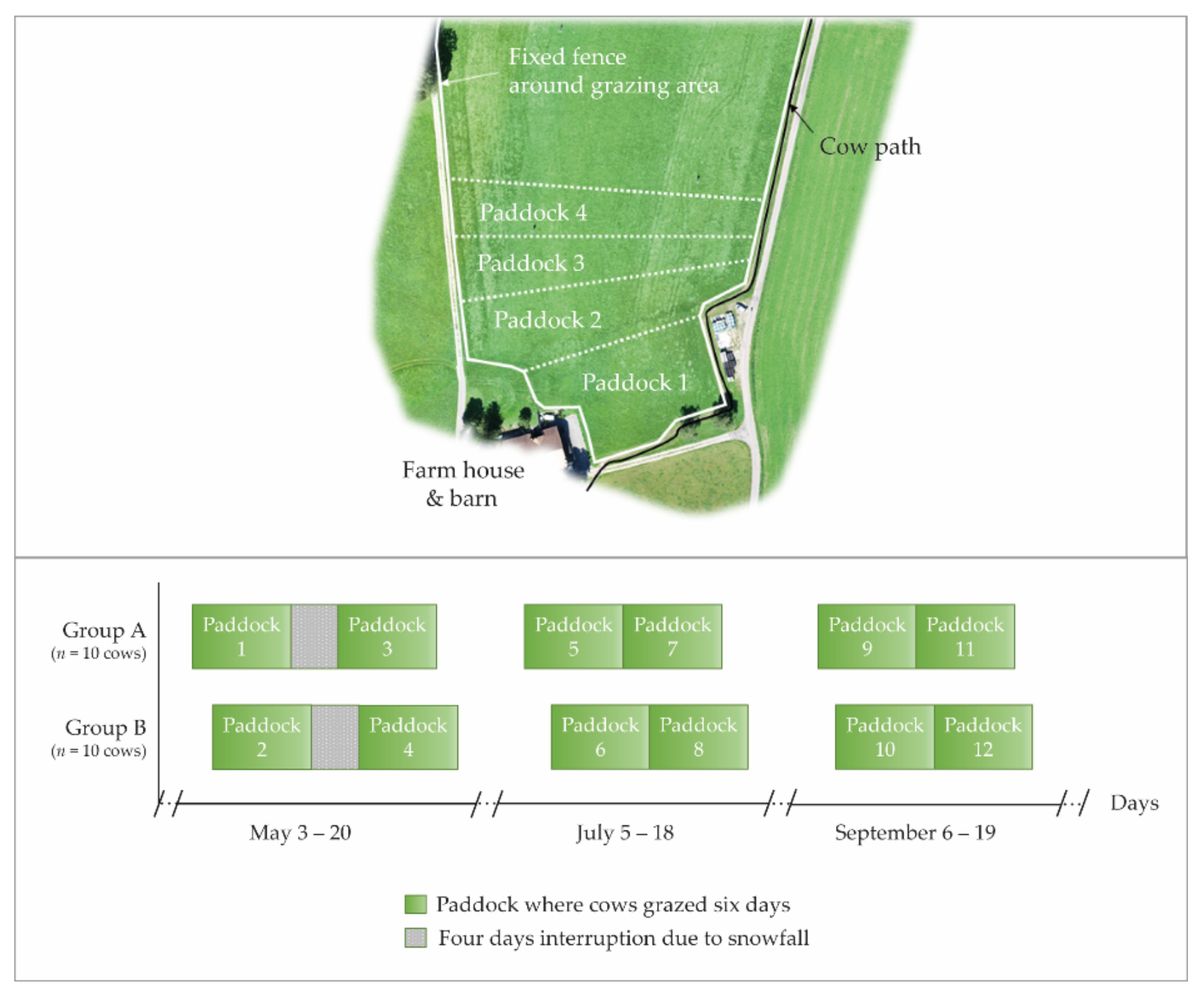
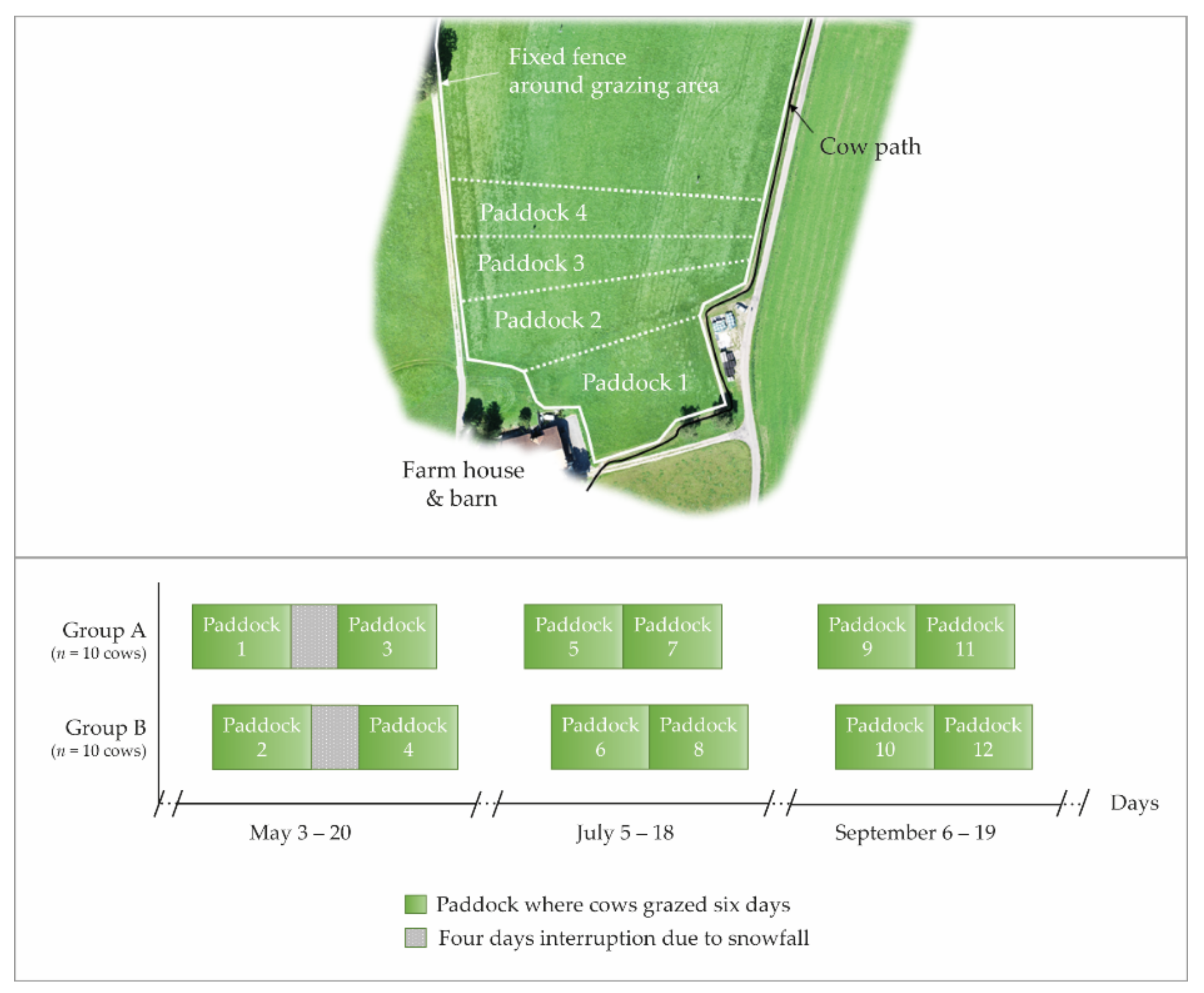
2.1. Grazing Site
2.2. Animals and Housing
2.3. Experimental Setup
2.4. Feed Allocation
2.4.1. Herbage Allowance
2.4.2. Allowance of Roughage and Concentrate Feeds Offered in the Barn
2.5. Pasture Measurements
2.6. Animal-Related Measurements
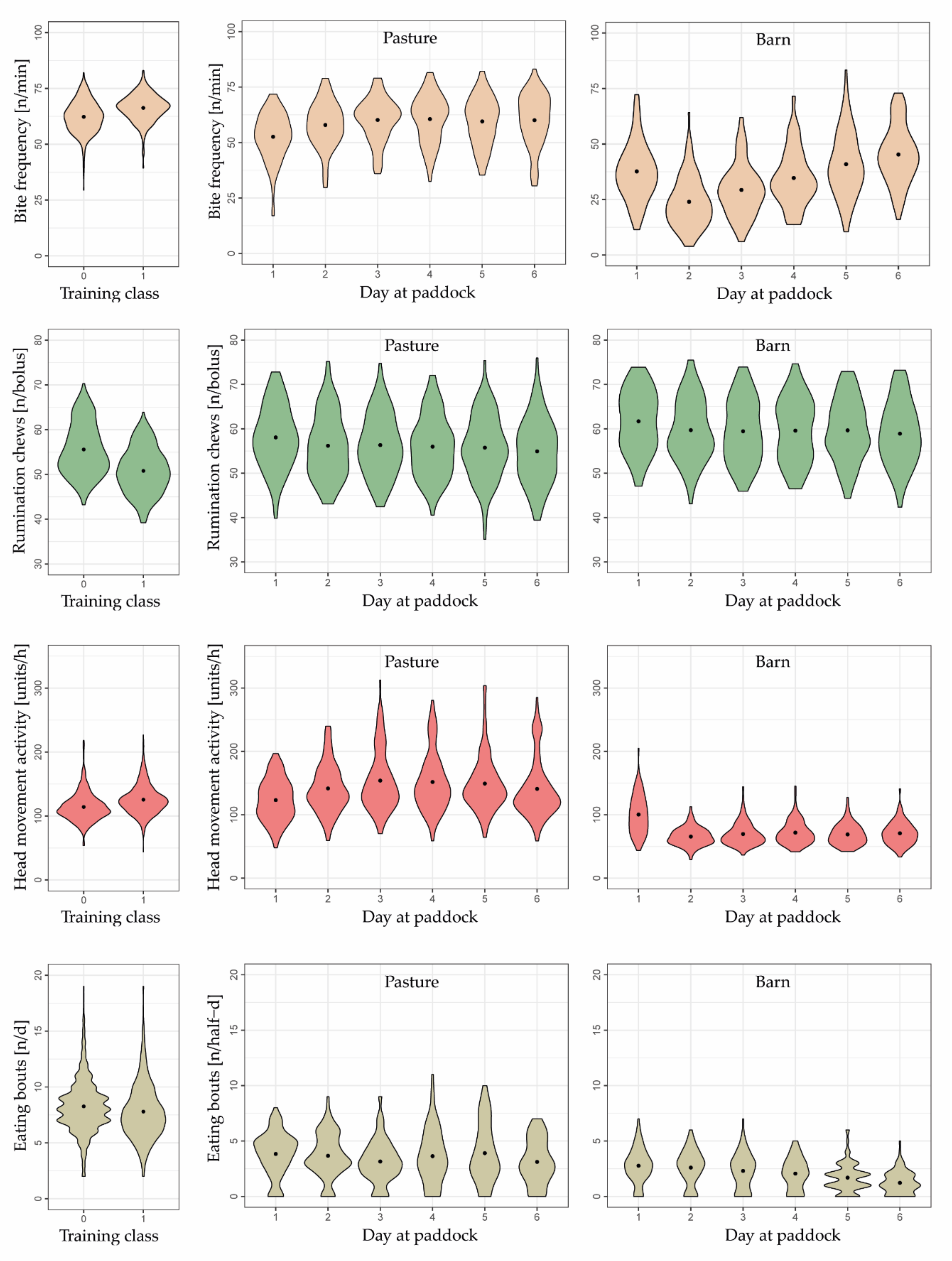
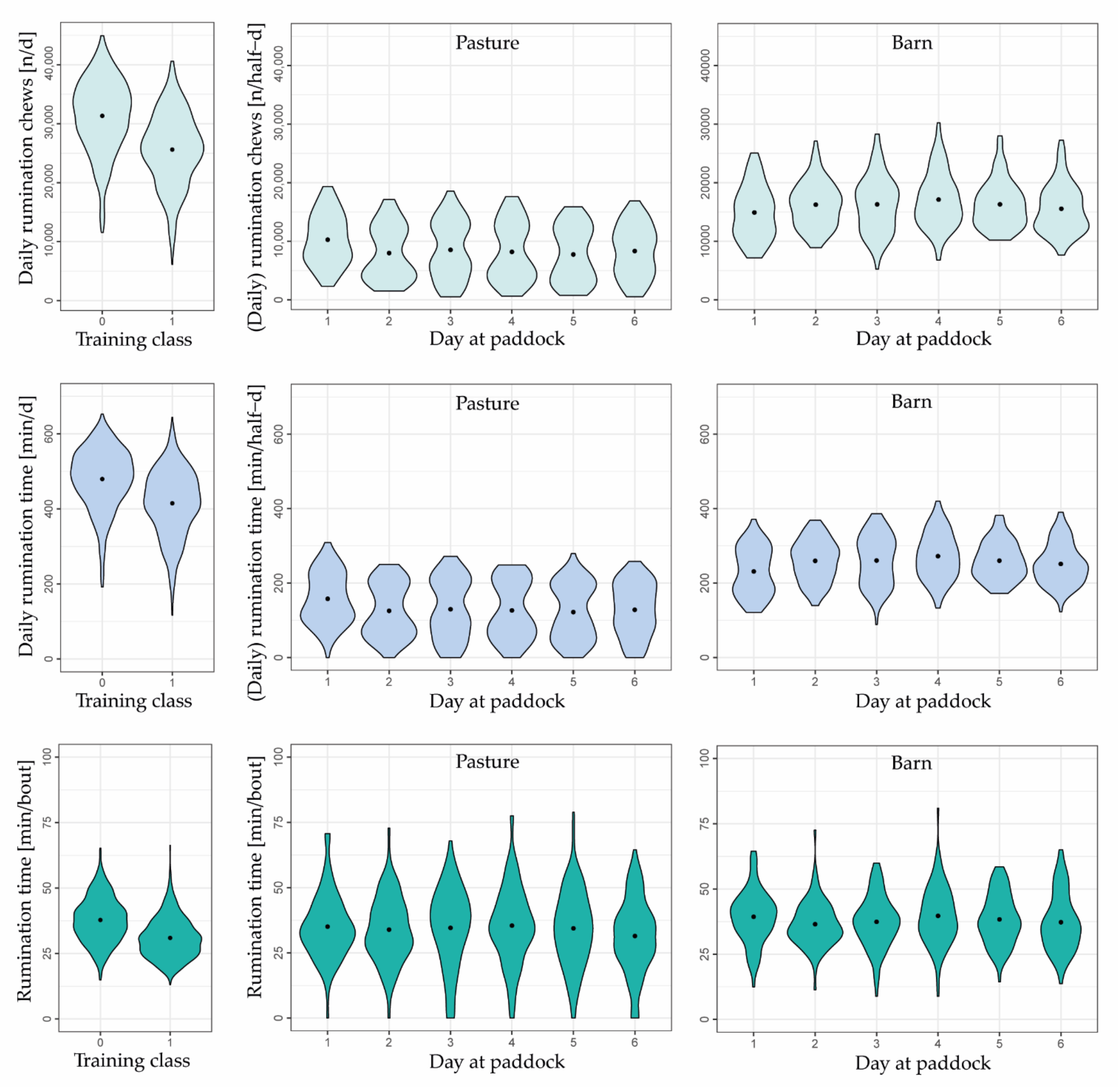
2.6.1. Behavior
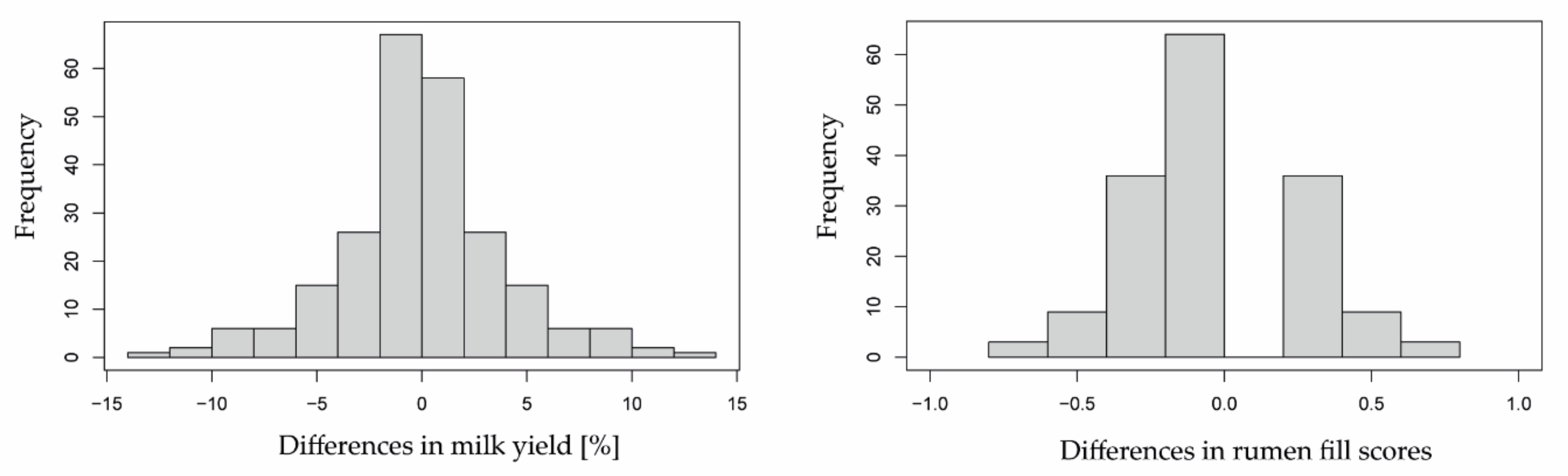
2.6.2. Milk Yield and Rumen Fill
2.7. Reference Indicators for Herbage Scarcity
2.7.1. Baseline Values
2.7.2. Reference Classification
2.8. Application of Prediction Models
2.9. Evaluation of Prediction Models
2.9.1. Predictive Performance
2.9.2. Variable Importance
3. Results
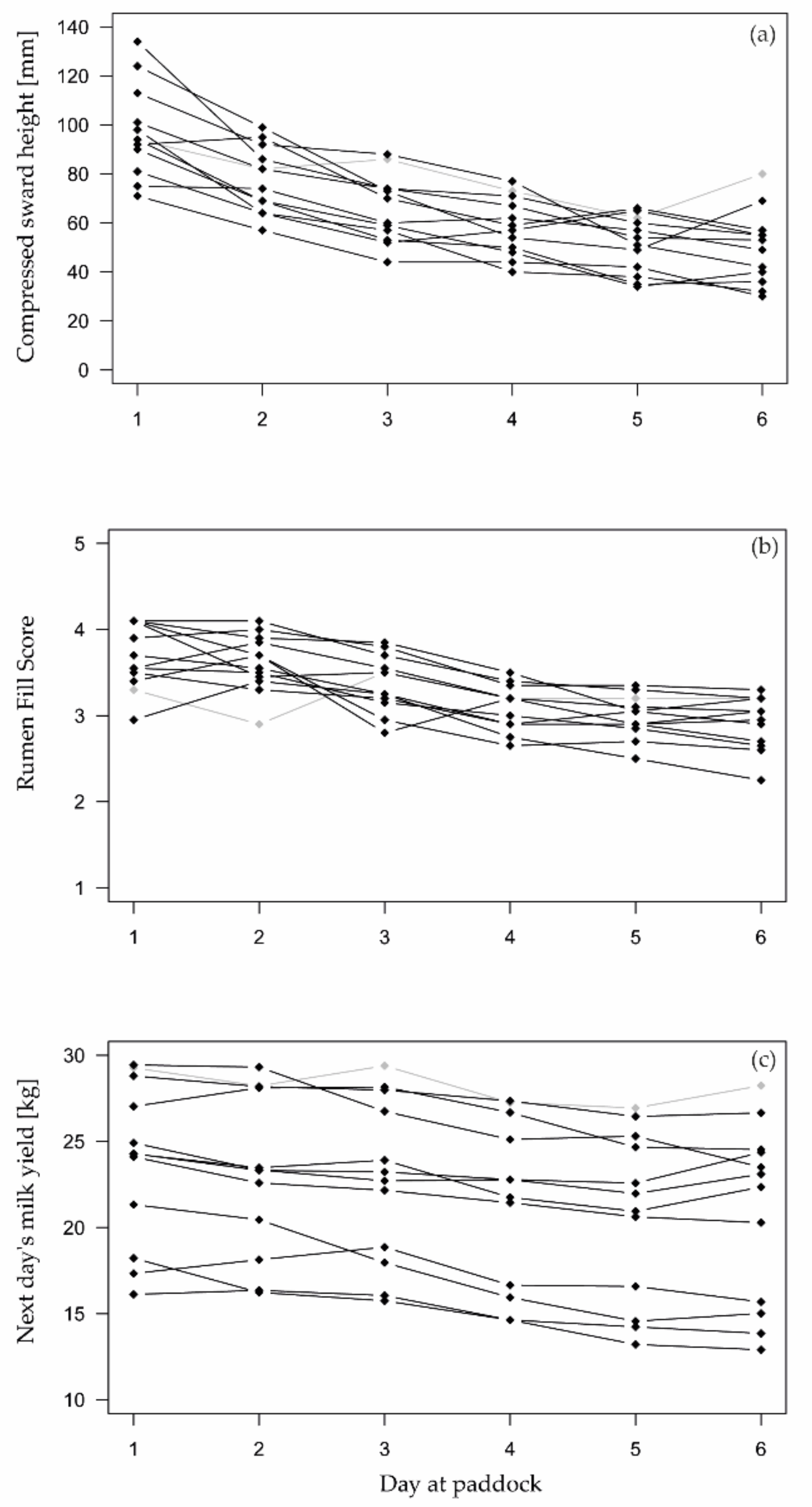
3.1. Effects of the Paddock Allocation
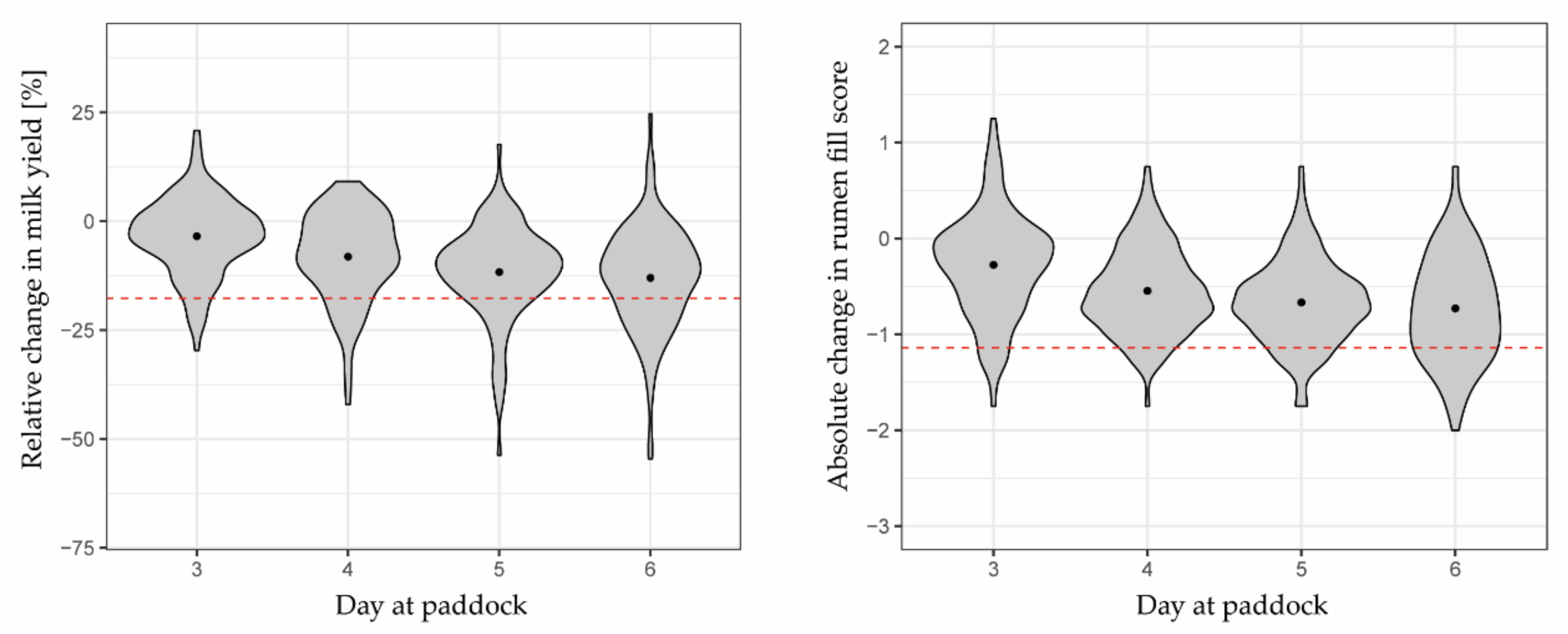
3.2. Herbage Availability According to Reference Indicators
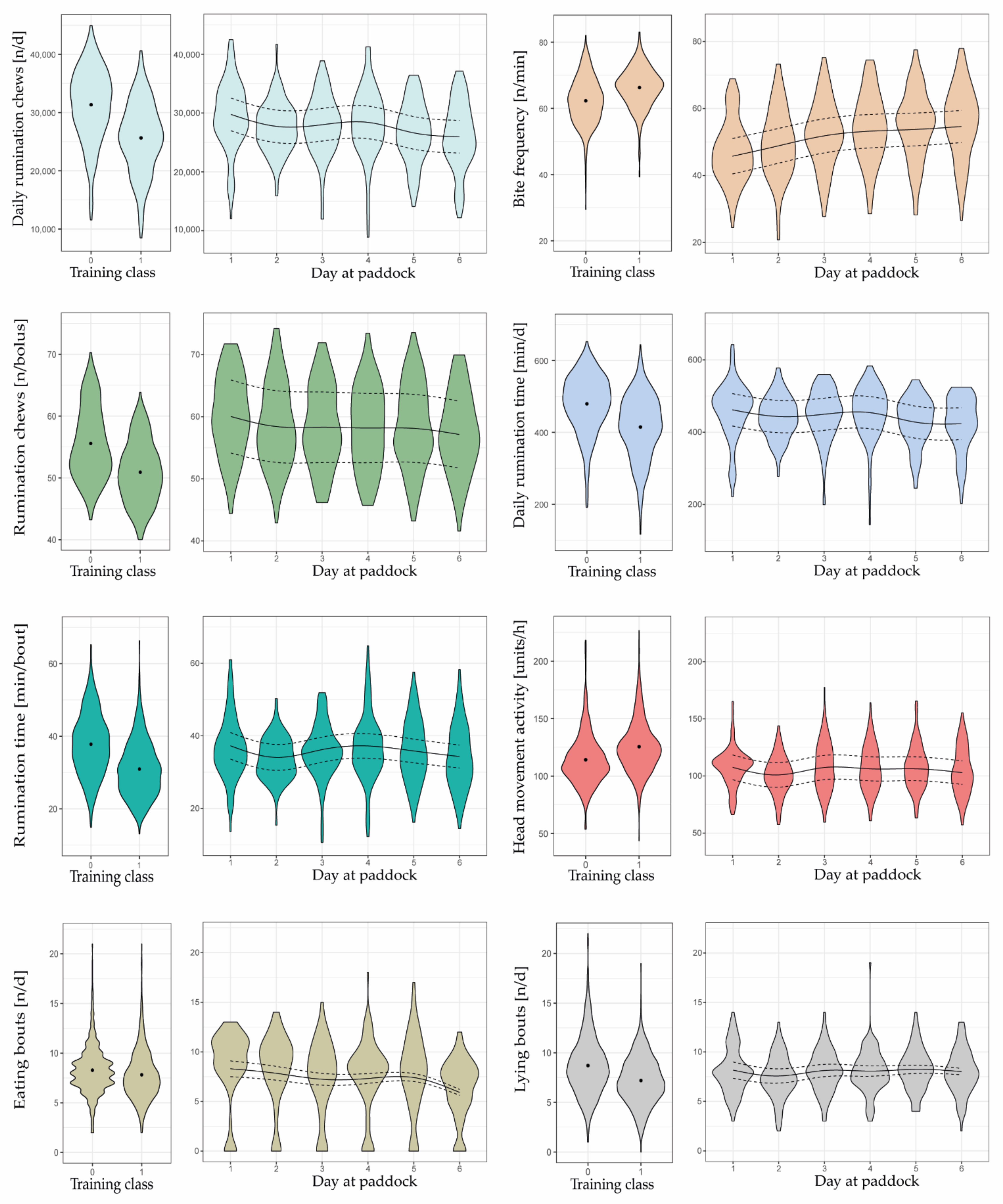
3.3. Behavioral Changes
3.4. Predicted Herbage Availability
3.5. Predictive Performance
4. Discussion
4.1. Differences between Training and Test Data as a Cause for Low Model Performance
4.2. The Challenge of Evaluating the Binary Predictions with Gradually Changing Reference Indicators
4.3. Suitability of Behavioral Variables as Predictors Depends on Grazing Condition
4.4. Outlook
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ordinary Linear Regression | Experimental Month | Sample Number | R2 |
|---|---|---|---|
| y = −1.8173 + 18.012 × x | May | 9 | 0.70 |
| y = 184.8 + 17.794 × x | July | 15 | 0.83 |
| y = 1368 + 15.069 × x | September | 15 | 0.34 |
| Subgroup (Number of Cow-Days) | Prediction Approach | Sensitivity | Specificity | PPV | AUC |
|---|---|---|---|---|---|
| Primiparous cows (n = 209) | GLM | 0.06 (0.09) | 0.91 (0.86) | 0.11 (0.11) | 0.48 (0.47) |
| RFM | 0.06 (0.37) | 0.92 (0.76) | 0.13 (0.24) | 0.49 (0.57) | |
| Multiparous cows (n = 374) | GLM | 0.15 (0.29) | 0.91 (0.84) | 0.26 (0.28) | 0.53 (0.56) |
| RFM | 0.03 (0.17) | 0.92 (0.84) | 0.08 (0.19) | 0.48 (0.51) | |
| High-performing cows (n = 186) | GLM | 0.22 (0.30) | 0.92 (0.82) | 0.28 (0.19) | 0.57 (0.56) |
| RFM | 0.04 (0.17) | 0.93 (0.83) | 0.08 (0.13) | 0.48 (0.50) | |
| Medium-performing cows (n = 212) | GLM | 0.15 (0.29) | 0.89 (0.87) | 0.20 (0.30) | 0.52 (0.58) |
| RFM | 0.06 (0.26) | 0.90 (0.82) | 0.10 (0.22) | 0.48 (0.54) | |
| Low-performing cows (n = 185) | GLM | 0.05 (0.11) | 0.91 (0.85) | 0.14 (0.19) | 0.48 (0.48) |
| RFM | 0.02 (0.25) | 0.94 (0.79) | 0.11 (0.27) | 0.48 (0.52) | |
| Cows with grazing experience (n = 274) | GLM | 0.20 (0.30) | 0.91 (0.83) | 0.30 (0.25) | 0.56 (0.56) |
| RFM | 0.05 (0.27) | 0.93 (0.83) | 0.11 (0.23) | 0.49 (0.55) | |
| Cows without grazing experience (n = 309) | GLM | 0.05 (0.16) | 0.90 (0.87) | 0.11 (0.21) | 0.48 (0.51) |
| RFM | 0.04 (0.21) | 0.91 (0.80) | 0.08 (0.19) | 0.47 (0.51) | |
| High BCS (n = 219) | GLM | 0.03 (0.10) | 0.91 (0.86) | 0.06 (0.13) | 0.47 (0.48) |
| RFM | 0.00 (0.23) | 0.92 (0.79) | 0.00 (0.20) | 0.46 (0.51) | |
| Medium BCS (n = 223) | GLM | 0.15 (0.23) | 0.88 (0.81) | 0.25 (0.25) | 0.51 (0.52) |
| RFM | 0.09 (0.21) | 0.93 (0.83) | 0.24 (0.25) | 0.51 (0.52) | |
| Low BCS (n = 141) | GLM | 0.27 (0.47) | 0.94 (0.88) | 0.33 (0.32) | 0.60 (0.67) |
| RFM | 0.00 (0.33) | 0.92 (0.82) | 0.00 (0.18) | 0.46 (0.58) | |
| Empty cows (n = 220) | GLM | 0.04 (0.08) | 0.91 (0.81) | 0.05 (0.05) | 0.47 (0.44) |
| RFM | 0.04 (0.32) | 0.90 (0.79) | 0.05 (0.17) | 0.47 (0.56) | |
| Cows in gestation (n = 363) | GLM | 0.14 (0.26) | 0.91 (0.87) | 0.29 (0.36) | 0.53 (0.57) |
| RFM | 0.04 (0.21) | 0.94 (0.83) | 0.14 (0.24) | 0.49 (0.52) |
| Reference Indicator | Prediction Approach | True Negative | False Negative | True Positive | False Positive |
|---|---|---|---|---|---|
| Milk yield | GLM | 461 | 65 | 7 | 50 |
| RFM | 471 | 68 | 4 | 40 | |
| Rumen fill scoring | GLM | 481 | 45 | 8 | 49 |
| RFM | 490 | 49 | 4 | 40 | |
| Both combined | GLM | 437 | 89 | 12 | 45 |
| RFM | 444 | 95 | 6 | 38 |

References
- Neethirajan, S.; Kemp, B. Digital Livestock Farming. Sens. Bio-Sens. Res. 2021, 32, 100408. [Google Scholar] [CrossRef]
- Akhigbe, B.I.; Munir, K.; Akinade, O.; Akanbi, L.; Oyedele, L.O. IoT Technologies for Livestock Management: A Review of Present Status, Opportunities, and Future Trends. Big Data Cogn. Comput. 2021, 5, 10. [Google Scholar] [CrossRef]
- Slob, N.; Catal, C.; Kassahun, A. Application of machine learning to improve dairy farm management: A systematic literature review. Prev. Vet. Med. 2021, 187, 105237. [Google Scholar] [CrossRef]
- Benos, L.; Tagarakis, A.C.; Dolias, G.; Berruto, R.; Kateris, D.; Bochtis, D. Machine Learning in Agriculture: A Comprehensive Updated Review. Sensors 2021, 21, 3758. [Google Scholar] [CrossRef]
- Cockburn, M. Review: Application and Prospective Discussion of Machine Learning for the Management of Dairy Farms. Animals 2020, 10, 1690. [Google Scholar] [CrossRef]
- Stygar, A.H.; Gómez, Y.; Berteselli, G.V.; Dalla Costa, E.; Canali, E.; Niemi, J.K.; Llonch, P.; Pastell, M. A Systematic Review on Commercially Available and Validated Sensor Technologies for Welfare Assessment of Dairy Cattle. Front. Vet. Sci. 2021, 8, 634338. [Google Scholar] [CrossRef]
- Shalloo, L.; O’ Donovan, M.; Leso, L.; Werner, J.; Ruelle, E.; Geoghegan, A.; Delaby, L.; O’Leary, N. Review: Grass-based dairy systems, data and precision technologies. Animal 2018, 12, s262–s271. [Google Scholar] [CrossRef] [Green Version]
- Hamidi, D.; Komainda, M.; Tonn, B.; Harbers, J.; Grinnell, N.A.; Isselstein, J. The Effect of Grazing Intensity and Sward Heterogeneity on the Movement Behavior of Suckler Cows on Semi-natural Grassland. Front. Vet. Sci. 2021, 8, 639096. [Google Scholar] [CrossRef]
- Murphy, D.J.; O’Brien, B.; Murphy, M.D. Development of a grass measurement optimisation tool to efficiently measure herbage mass on grazed pastures. Comput. Electron. Agric. 2020, 178, 105799. [Google Scholar] [CrossRef]
- Deming, J.; Gleeson, D.; O’Dwyer, T.; Kinsella, J.; O’Brien, B. Measuring labor input on pasture-based dairy farms using a smartphone. J. Dairy Sci. 2018, 101, 9527–9543. [Google Scholar] [CrossRef]
- Murphy, D.J.; O’ Brien, B.; Hennessy, D.; Hurley, M.; Murphy, M.D. Evaluation of the precision of the rising plate meter for measuring compressed sward height on heterogeneous grassland swards. Precis. Agric. 2021, 22, 922–946. [Google Scholar] [CrossRef]
- Sishodia, R.P.; Ray, R.L.; Singh, S.K. Applications of Remote Sensing in Precision Agriculture: A Review. Remote Sens. 2020, 12, 3136. [Google Scholar] [CrossRef]
- Sinde-González, I.; Gil-Docampo, M.; Arza-García, M.; Grefa-Sánchez, J.; Yánez-Simba, D.; Pérez-Guerrero, P.; Abril-Porras, V. Biomass estimation of pasture plots with multitemporal UAV-based photogrammetric surveys. Int. J. Appl. Earth Obs. Geoinf. 2021, 101, 102355. [Google Scholar] [CrossRef]
- Nickmilder, C.; Tedde, A.; Dufrasne, I.; Lessire, F.; Tychon, B.; Curnel, Y.; Bindelle, J.; Soyeurt, H. Development of Machine Learning Models to Predict Compressed Sward Height in Walloon Pastures Based on Sentinel-1, Sentinel-2 and Meteorological Data Using Multiple Data Transformations. Remote Sens. 2021, 13, 408. [Google Scholar] [CrossRef]
- Decruyenaere, V.; Buldgen, A.; Stilmant, D. Factors affecting intake by grazing ruminants and related quantification methods: A review. BASE 2009, 13, 559–573. [Google Scholar]
- O’Driscoll, K.; Lewis, E.; Kennedy, E. Effect of feed allowance at pasture on the lying behaviour of dairy cows. Appl. Anim. Behav. Sci. 2019, 213, 40–46. [Google Scholar] [CrossRef]
- Werner, J.; Umstatter, C.; Kennedy, E.; Grant, J.; Leso, L.; Geoghegan, A.; Shalloo, L.; Schick, M.; O’Brien, B. Identification of possible cow grazing behaviour indicators for restricted grass availability in a pasture-based spring calving dairy system. Livest. Sci. 2019, 220, 74–82. [Google Scholar] [CrossRef]
- Rombach, M.; Südekum, K.H.; Münger, A.; Schori, F. Herbage dry matter intake estimation of grazing dairy cows based on animal, behavioral, environmental, and feed variables. J. Dairy Sci. 2019, 102, 2985–2999. [Google Scholar] [CrossRef] [Green Version]
- Shafiullah, A.Z.; Werner, J.; Kennedy, E.; Leso, L.; O’Brien, B.; Umstätter, C. Machine Learning Based Prediction of Insufficient Herbage Allowance with Automated Feeding Behaviour and Activity Data. Sensors 2019, 19, 4479. [Google Scholar] [CrossRef] [Green Version]
- Post, C.; Rietz, C.; Büscher, W.; Müller, U. The Importance of Low Daily Risk for the Prediction of Treatment Events of Individual Dairy Cows with Sensor Systems. Sensors 2021, 21, 1389. [Google Scholar] [CrossRef]
- FiBL. Koerper-Konditions-Beurteilung; Spengler Neff, A., Notz, C., Ivemeyer, S., Walkenhorst, M., Eds.; Forschungsinstitut fuer biologischen Landbau (FiBL): Frick, Switzerland, 2015; p. 2. [Google Scholar]
- Pérez-Ramírez, E.; Peyraud, J.; Delagarde, R. Restricting daily time at pasture at low and high pasture allowance: Effects on pasture intake and behavioral adaptation of lactating dairy cows. J. Dairy Sci. 2009, 92, 3331–3340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulati, A.; Galvin, N.; Kennedy, E.; Lewis, E.; McManus, J.J.; Fenelon, M.A.; Guinee, T.P. Effect of reducing daily herbage allowance during early lactation on composition and processing characteristics of milk from spring-calved herds. Int. Dairy J. 2019, 92, 69–76. [Google Scholar] [CrossRef]
- Dessauge, F.; Lollivier, V.; Ponchon, B.; Bruckmaier, R.; Finot, L.; Wiart, S.; Cutullic, E.; Disenhaus, C.; Barbey, S.; Boutinaud, M. Effects of nutrient restriction on mammary cell turnover and mammary gland remodeling in lactating dairy cows. J. Dairy Sci. 2011, 94, 4623–4635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, J.; van Dorland, H.A.; Bruckmaier, R.M.; Schwarz, F.J. Performance and metabolic profile of dairy cows during a lactational and deliberately induced negative energy balance with subsequent realimentation. J. Dairy Sci. 2011, 94, 1820–1830. [Google Scholar] [CrossRef]
- AGFF. Weiden von Milchkühen: Berechnung der angepassten Besatzstärke. In Weide- und Alpwirtschaft Information W10; Mosimann, E., Stettler, M., Eds.; Arbeitsgemeinschaft zur Förderung des Futterbaus: Zürich, Switzerland, 2004; p. 2. [Google Scholar]
- Zehner, N.; Umstätter, C.; Niederhauser, J.J.; Schick, M. System specification and validation of a noseband pressure sensor for measurement of ruminating and eating behavior in stable-fed cows. Comput. Electron. Agric. 2017, 136, 31–41. [Google Scholar] [CrossRef]
- Werner, J.; Leso, L.; Umstatter, C.; Niederhauser, J.; Kennedy, E.; Geoghegan, A.; Shalloo, L.; Schick, M.; O’Brien, B. Evaluation of the RumiWatchSystem for measuring grazing behaviour of cows. J. Neurosci. Methods 2018, 300, 138–146. [Google Scholar] [CrossRef]
- Reith, S.; Hoy, S. Relationship between daily rumination time and estrus of dairy cows. J. Dairy Sci. 2012, 95, 6416–6420. [Google Scholar] [CrossRef]
- Zaaijer, D.; Noordhuizen, J.P.T.M. A novel scoring system for monitoring the relationship between nutritional efficiency and fertility in dairy cows. Ir. Vet. J. 2003, 56, 145–151. [Google Scholar]
- Burfeind, O.; Sepúlveda, P.; von Keyserlingk, M.A.G.; Weary, D.M.; Veira, D.M.; Heuwieser, W. Technical note: Evaluation of a scoring system for rumen fill in dairy cows. J. Dairy Sci. 2010, 93, 3635–3640. [Google Scholar] [CrossRef]
- Agroscope. Frisst die Kuh Genug? Available online: https://www.agroscope.admin.ch/agroscope/de/home/aktuell/newsroom/2021/04-07_bewertung-hungergrube-kuh.html (accessed on 21 September 2021).
- Schneider, M.; Hart, L.; Gallmann, E.; Umstaetter, C. A Novel Chart to Score Rumen Fill Following Simple Sequential Instructions. 2022; under review. [Google Scholar]
- Huhtanen, P.; Hetta, M. Comparison of feed intake and milk production responses in continuous and change-over design dairy cow experiments. Livest. Sci. 2012, 143, 184–194. [Google Scholar] [CrossRef]
- Faverdin, P.; Baratte, C.; Delagarde, R.; Peyraud, J.L. GrazeIn: A model of herbage intake and milk production for grazing dairy cows. 1. Prediction of intake capacity, voluntary intake and milk production during lactation. Grass Forage Sci. 2011, 66, 29–44. [Google Scholar] [CrossRef]
- Hristov, A.N.; Price, W.J.; Shafii, B. A Meta-Analysis Examining the Relationship Among Dietary Factors, Dry Matter Intake, and Milk and Milk Protein Yield in Dairy Cows. J. Dairy Sci. 2004, 87, 2184–2196. [Google Scholar] [CrossRef] [Green Version]
- Delagarde, R.; Faverdin, P.; Baratte, C.; Peyraud, J.L. GrazeIn: A model of herbage intake and milk production for grazing dairy cows. 2. Prediction of intake under rotational and continuously stocked grazing management. Grass Forage Sci. 2011, 66, 45–60. [Google Scholar] [CrossRef]
- Oudshoorn, F.W.; Cornou, C.; Hellwing, A.L.F.; Hansen, H.H.; Munksgaard, L.; Lund, P.; Kristensen, T. Estimation of grass intake on pasture for dairy cows using tightly and loosely mounted di- and tri-axial accelerometers combined with bite count. Comput. Electron. Agric. 2013, 99, 227–235. [Google Scholar] [CrossRef]
- Fisher, R. Statistical Methods for Research Workers; Oliver and Boyd: Edinburgh, UK, 1925. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 25 June 2021).
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Kuhn, M. Building predictive models in R using the caret package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Lindstrom, M.J.; Bates, D.M. Nonlinear Mixed Effects Models for Repeated Measures Data. Biometrics 1990, 46, 673–687. [Google Scholar] [CrossRef]
- Hastie, T.J. Generalized additive models. In Statistical Models in S; Chambers, J.M., Hastie, T.J., Eds.; Wadsworth & Brooks/Cole: London, UK, 1992; pp. 249–308. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; p. 224. [Google Scholar]
- Herve, L.; Quesnel, H.; Veron, M.; Portanguen, J.; Gross, J.J.; Bruckmaier, R.M.; Boutinaud, M. Milk yield loss in response to feed restriction is associated with mammary epithelial cell exfoliation in dairy cows. J. Dairy Sci. 2019, 102, 2670–2685. [Google Scholar] [CrossRef] [Green Version]
- Götze, K.; Crivellaro, P.; Pieper, L.; Engelhard, T.; Staufenbiel, R. Assessment of rumen fill in dairy cows for evaluation of the individual feed intake in herd management. Tierartzl. Prax. Ausg. G Grosstiere Nutztiere 2019, 2019, 5–13. [Google Scholar] [CrossRef]
- Bareille, N.; Beaudeau, F.; Billon, S.; Robert, A.; Faverdin, P. Effects of health disorders on feed intake and milk production in dairy cows. Livest. Prod. Sci. 2003, 83, 53–62. [Google Scholar] [CrossRef]
- Norring, M.; Häggman, J.; Simojoki, H.; Tamminen, P.; Winckler, C.; Pastell, M. Short communication: Lameness impairs feeding behavior of dairy cows. J. Dairy Sci. 2014, 97, 4317–4321. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.H.; Collier, R.J.; Bauman, D.E. A 100-Year Review: Regulation of nutrient partitioning to support lactation. J. Dairy Sci. 2017, 100, 10353–10366. [Google Scholar] [CrossRef] [PubMed]
- Post, C.; Rietz, C.; Büscher, W.; Müller, U. Using Sensor Data to Detect Lameness and Mastitis Treatment Events in Dairy Cows: A Comparison of Classification Models. Sensors 2020, 20, 3863. [Google Scholar] [CrossRef] [PubMed]
- Schils, R.; Philipsen, B.; Hoekstra, N.; Holshof, G.; Zom, R.; Hoving, I.; van Reenen, K.; Stienezen, M.; Klootwijk, C.; van der Werf, J.; et al. Amazing Grazing: A Public and Private Partnership to Stimulate Grazing Practices in Intensive Dairy Systems. Sustainability 2019, 11, 5868. [Google Scholar] [CrossRef] [Green Version]
- Halachmi, I.; Ben Meir, Y.; Miron, J.; Maltz, E. Feeding behavior improves prediction of dairy cow voluntary feed intake but cannot serve as the sole indicator. Animal 2016, 10, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Andriamandroso, A.L.H.; Lebeau, F.; Bindelle, J. Changes in biting characteristics recorded using the inertial measurement unit of a smartphone reflect differences in sward attributes. In Proceedings of the 7th European Conference on Precision Livestock Farming, Milan, Italy, 15–18 September 2015; pp. 283–289. [Google Scholar]
- Gregorini, P.; DelaRue, B.; McLeod, K.; Clark, C.E.F.; Glassey, C.B.; Jago, J. Rumination behavior of grazing dairy cows in response to restricted time at pasture. Livest. Sci. 2012, 146, 95–98. [Google Scholar] [CrossRef]
- Kennedy, E.; McEvoy, M.; Murphy, J.P.; O’Donovan, M. Effect of restricted access time to pasture on dairy cow milk production, grazing behavior, and dry matter intake. J. Dairy Sci. 2009, 92, 168–176. [Google Scholar] [CrossRef]
- Pérez-Ramírez, E.; Delagarde, R.; Delaby, L. Herbage intake and behavioural adaptation of grazing dairy cows by restricting time at pasture under two feeding regimes. Animal 2008, 2, 1384–1392. [Google Scholar] [CrossRef] [Green Version]
- Paulenz, E.; Gygax, L.; Barth, K.; Hart, L.; Hillmann, E. Effect of Sward Height on the Behavior of Dairy Cows in a Rotational Grazing System. 2022; manuscript in preparation. [Google Scholar]
- Krizsan, S.J.; Sairanen, A.; Höjer, A.; Huhtanen, P. Evaluation of different feed intake models for dairy cows. J. Dairy Sci. 2014, 97, 2387–2397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codrea, M.C.; Højsgaard, S.; Friggens, N.C. Differential smoothing of time-series measurements to identify disturbances in performance and quantify animal response characteristics: An example using milk yield profiles in dairy cows1. J. Anim. Sci. 2011, 89, 3089–3098. [Google Scholar] [CrossRef] [PubMed]




| Number of Grazing Cycle and Month | Cow Group | Paddock ID | Paddock Area (ha) | Pre-Grazing CSH (mm) | Allocated HM at Beginning of Grazing Cycles (kg DM Group−1 Paddock−1) * |
|---|---|---|---|---|---|
| 1–May | A | 1 | 0.52 | 93 | 679 # |
| 1–May | B | 2 | 0.31 | 117 | 534 |
| 2–May | A | 3 | 0.19 | 164 | 496 |
| 2–May | B | 4 | 0.19 | 162 | 494 |
| 3–July | A | 5 | 0.34 | 89 | 476 |
| 3–July | B | 6 | 0.29 | 104 | 493 |
| 4–July | A | 7 | 0.25 | 125 | 509 |
| 4–July | B | 8 | 0.27 | 113 | 500 |
| 5–September | A | 9 | 0.17 | 152 | 569 |
| 5–September | B | 10 | 0.19 | 125 | 564 |
| 6–September | A | 11 | 0.20 | 120 | 564 |
| 6–September | B | 12 | 0.19 | 131 | 565 |
| Variable (Unit) | Corresponding Variable Name * |
|---|---|
| Daily rumination chews (n d−1) | RUMINATECHEW |
| Bite frequency (n min−1), represents the prehension bites with head down during eating | BITEFREQ |
| Rumination chews (n bolus−1) | RUMICHEWBOLUS |
| Head movement activity index (units h−1), calculated from acceleration in x, y, and z directions | HACTIVITY |
| Rumination time (min bout−1) | RUMIBOUTLENGTH |
| Daily rumination time (min d−1), calculated as the sum of rumination bout minutes | RUMIBOUTTIME |
| Eating bouts (n d−1) | GRAZINGSTART |
| Lying bouts (n d−1) | LAYDOWN |
| Subgroup | Splitting Criteria | Number of Cow-Days (n) |
|---|---|---|
| Primiparous cows | Parity = 1 | 209 |
| Multiparous cows | Parity > 1 | 374 |
| Cows with grazing experience | Cows grazed in previous years | 274 |
| Cows without grazing experience | Cows never grazed previously | 309 |
| High-performing cows | Milk yield > 27 kg d−1 (mean: 31.6) | 186 |
| Medium-performing cows | Milk yield 20–27 kg d−1 (mean: 23.8) | 212 |
| Low-performing cows | Milk yield < 20 kg d−1 (mean: 16.5) | 185 |
| Empty cows | Gestation verified | 220 |
| Cows in gestation | Gestation not verified | 363 |
| High BCS | BCS ≥ 3.25 | 219 |
| Medium BCS | BCS 2.75–3.00 | 223 |
| Low BCS | BCS ≤ 2.5 | 141 |
| Reference Methods | Day 1 + (n = 91) | Day 2 + (n = 92) | Day 3 (n = 101) | Day 4 (n = 101) | Day 5 (n = 99) | Day 6 (n = 99) | Total (n = 583) |
|---|---|---|---|---|---|---|---|
| Milk yield | 0 | 0 | 8 | 14 | 19 | 31 | 72 |
| Rumen fill score | 0 | 0 | 7 | 8 | 14 | 24 | 53 |
| Both combined * | 0 | 0 | 13 | 18 | 28 | 42 | 101 |
| Prediction Approach | Day 1 (n = 91) | Day 2 (n = 92) | Day 3 (n = 101) | Day 4 (n = 101) | Day 5 (n = 99) | Day 6 (n = 99) | Total (n = 583) |
|---|---|---|---|---|---|---|---|
| GLM | 0 | 4 | 7 | 12 | 13 | 21 | 57 |
| RFM | 0 | 2 | 9 | 10 | 8 | 16 | 45 |
| Predicted Versus Reference Classification | Sensitivity | Specificity | PPV | AUC |
|---|---|---|---|---|
| GLM versus MYRef | 0.10 (0.19) | 0.90 (0.84) | 0.12 (0.15) | 0.50 (0.52) |
| RFM versus MYRef | 0.06 (0.25) | 0.92 (0.81) | 0.09 (0.16) | 0.49 (0.53) |
| GLM versus RFRef | 0.15 (0.25) | 0.91 (0.84) | 0.14 (0.14) | 0.53 (0.54) |
| RFM versus RFRef | 0.06 (0.28) | 0.92 (0.82) | 0.07 (0.13) | 0.49 (0.55) |
| GLM versus COMBRef | 0.12 (0.22) | 0.91 (0.85) | 0.21 (0.23) | 0.51 (0.53) |
| RFM versus COMBRef | 0.06 (0.24) | 0.92 (0.82) | 0.13 (0.21) | 0.49 (0.53) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hart, L.; Dickhoefer, U.; Paulenz, E.; Umstaetter, C. Evaluation of a Binary Classification Approach to Detect Herbage Scarcity Based on Behavioral Responses of Grazing Dairy Cows. Sensors 2022, 22, 968. https://doi.org/10.3390/s22030968
Hart L, Dickhoefer U, Paulenz E, Umstaetter C. Evaluation of a Binary Classification Approach to Detect Herbage Scarcity Based on Behavioral Responses of Grazing Dairy Cows. Sensors. 2022; 22(3):968. https://doi.org/10.3390/s22030968
Chicago/Turabian StyleHart, Leonie, Uta Dickhoefer, Esther Paulenz, and Christina Umstaetter. 2022. "Evaluation of a Binary Classification Approach to Detect Herbage Scarcity Based on Behavioral Responses of Grazing Dairy Cows" Sensors 22, no. 3: 968. https://doi.org/10.3390/s22030968
APA StyleHart, L., Dickhoefer, U., Paulenz, E., & Umstaetter, C. (2022). Evaluation of a Binary Classification Approach to Detect Herbage Scarcity Based on Behavioral Responses of Grazing Dairy Cows. Sensors, 22(3), 968. https://doi.org/10.3390/s22030968