Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella aurita

Abstract
:1. Introduction
2. Results and Discussion
2.1. Isolation, Purification and Characterization of Chrysolaminarin
2.1.1. Purification and Homogeneity of Chrysolaminarin


2.1.2. Monosaccharide Composition and Molecular Weight of CL2

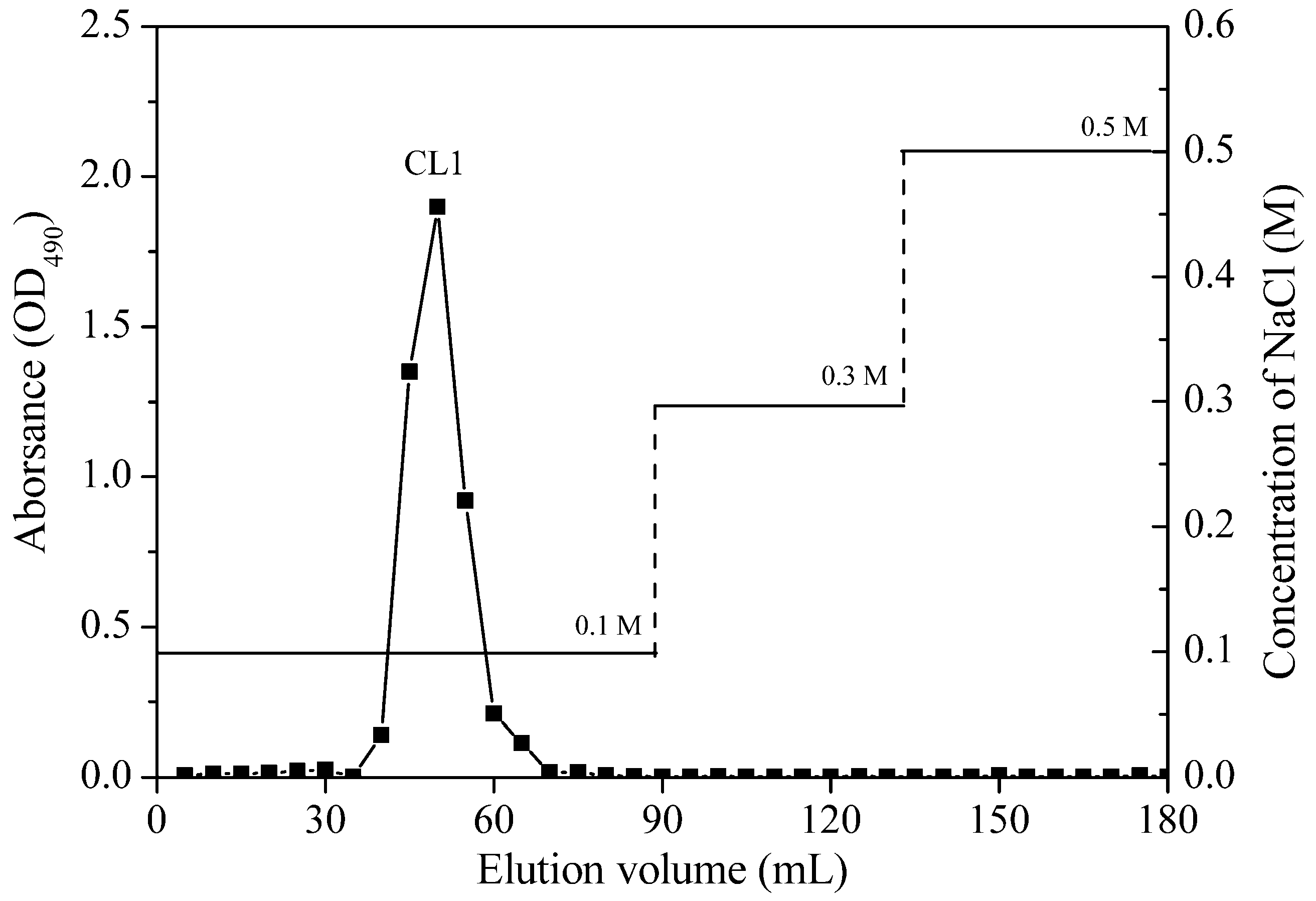{kind=link}
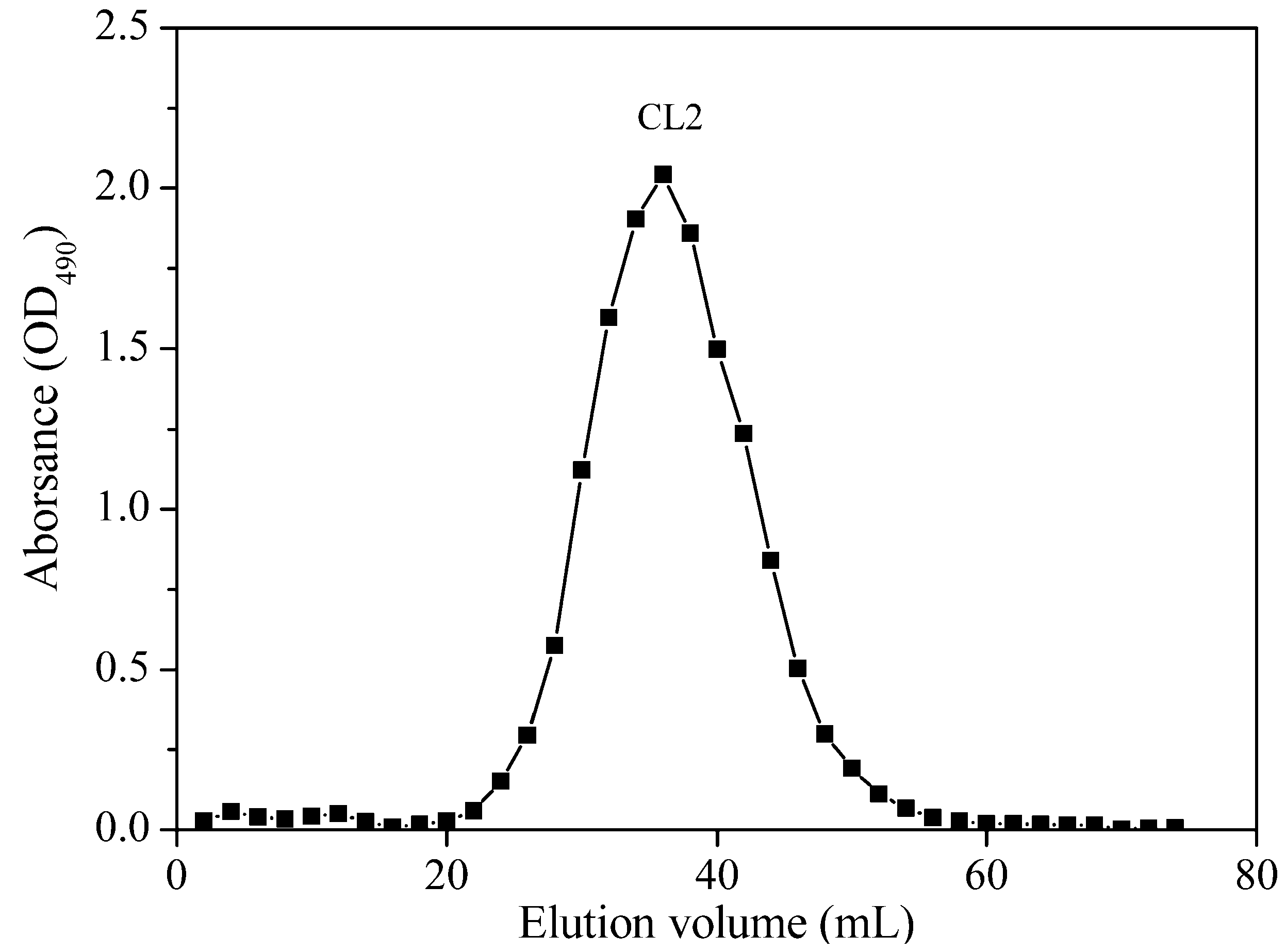{kind=link}
{kind=link}
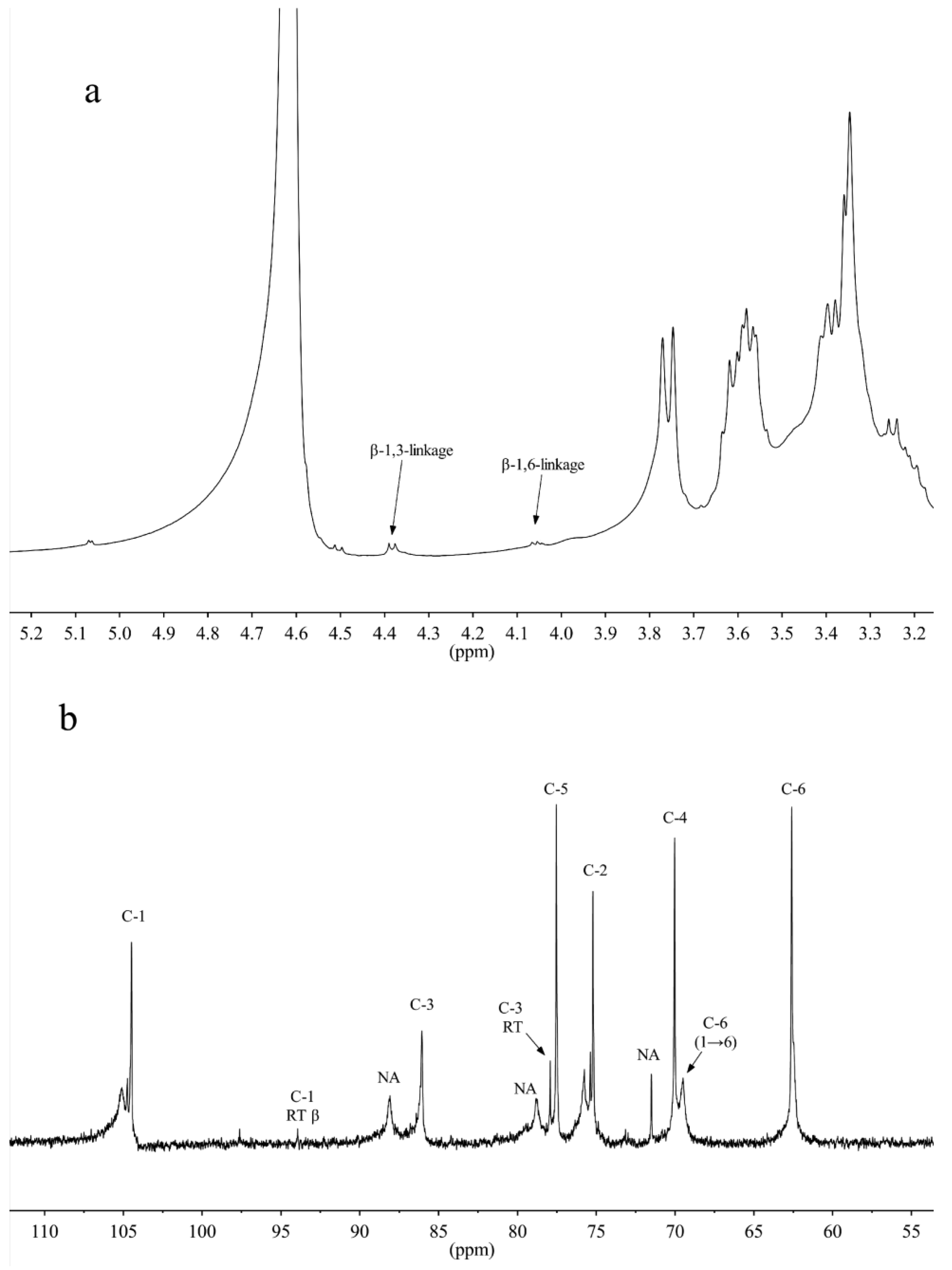{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar Components (Total Sugar a %) | |||||
|---|---|---|---|---|---|
| Glucose | Mannose | Ribose | Arabinose | Xylose | Galactose |
| 82.23 | 13.27 | 0.46 | 3.62 | 0.26 | 0.16 |
2.1.3. FTIR Spectra Analysis

2.1.4. NMR Spectra Analysis

2.2. Assay for Antioxidant Activity
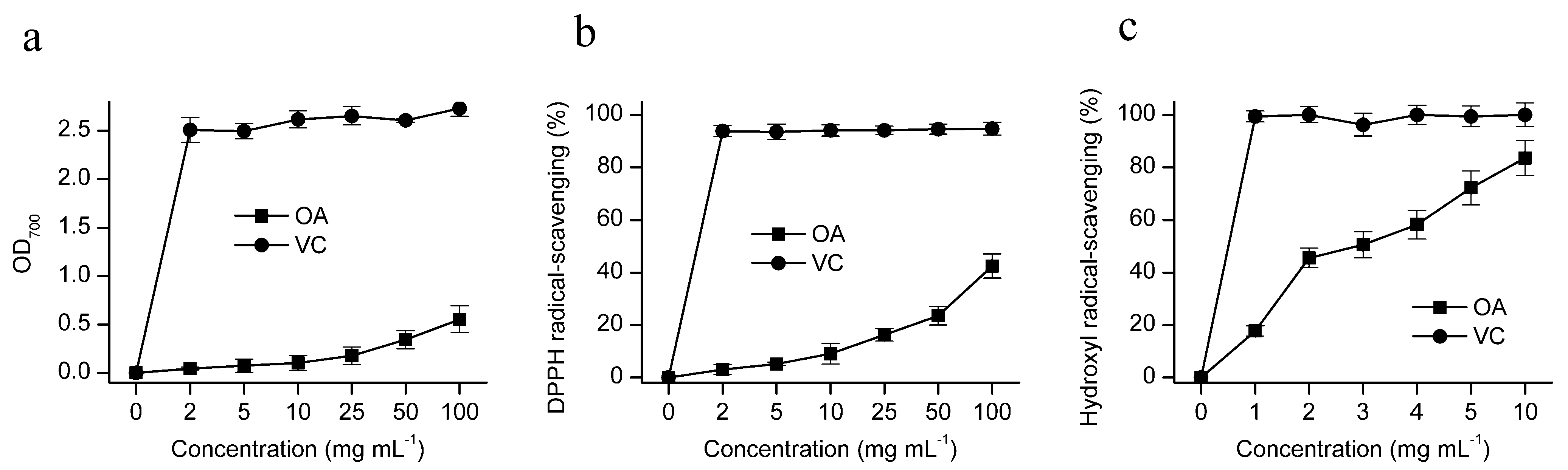
2.2.1. Reducing Power

2.2.2. DPPH Radical Scavenging Activity
2.2.3. Hydroxyl Radical Scavenging Activity
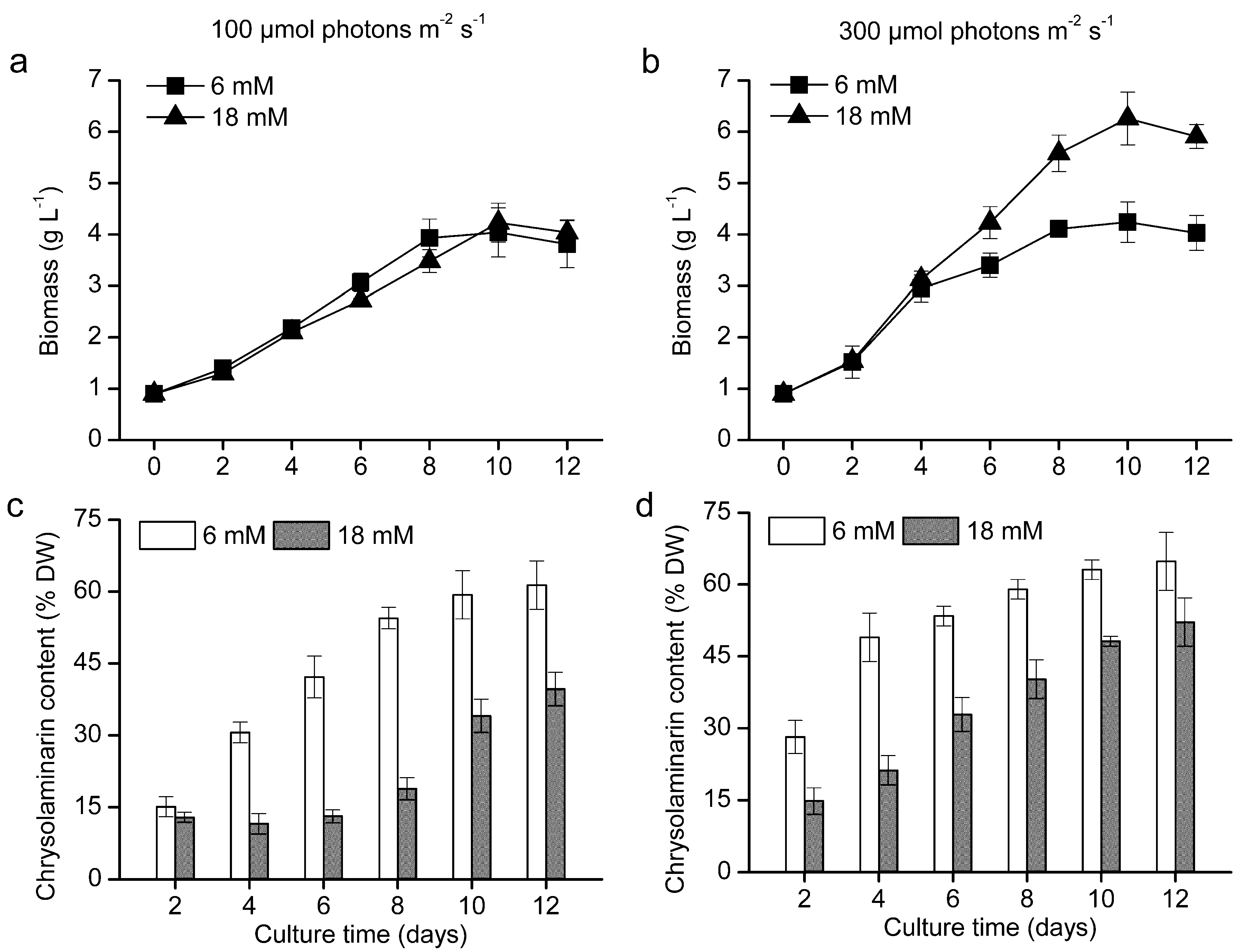
2.3. Production of Chrysolaminarin from O. aurita in a Column Photobioreactor

| Culture Condition a | Biomass Concentration (g L−1) | Chrysolaminarin Content (% Dry Weight) | Chrysolaminarin Productivity (mg L−1 Day−1) |
|---|---|---|---|
| LL + LN | 4.04 | 59.33 | 240 |
| LL + HN | 4.23 | 34.05 | 144 |
| HL + LN | 4.24 | 63.11 | 268 |
| HL + HN | 6.36 | 48.16 | 306 |
3. Experimental Section
3.1. Organism and Culture Conditions
3.2. Biomass Measurement
3.3. Determination of Chrysolaminarin Content
3.4. Preparation of Chrysolaminarin

3.5. Structural Analysis
3.5.1. Molecular Weight
3.5.2. Monosaccharide Composition
3.5.3. IR Spectroscopy
3.5.4. NMR Analysis
3.6. Antioxidant Activity Assessment
3.6.1. Reducing Power
3.6.2. DPPH Radical Scavenging Activity
3.6.3. Hydroxyl Radical Scavenging Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of aging. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Yang, X.; Wang, R.; Zhang, S.; Zhu, W.; Tang, J.; Liu, J.; Chen, P.; Zhang, D.; Ye, W.; Zheng, Y.; et al. Polysaccharides from Panax japonicus C.A. Meyer and their antioxidant activities. Carbohydr. Polym. 2014, 101, 386–391. [Google Scholar]
- Zhang, Z.S.; Wang, X.M.; Mo, X.F.; Qi, H.M. Degradation and the antioxidant activity of polysaccharide from Enteromorpha linza. Carbohydr. Polym. 2013, 92, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.; Nandan, C.K.; Bhunia, S.K.; Maiti, S.; Maiti, T.K.; Sikdar, S.R.; Islam, S.S. Glucans from alkaline extract of a hybrid mushroom (backcross mating between PfloVv12 and Volvariella volvacea): Structural characterization and study of immunoenhancing and antioxidant properties. Carbohydr. Res. 2012, 347, 107–113. [Google Scholar] [CrossRef]
- Xia, S.; Li, A.F.; Wan, L.L.; Sang, M.; Zhang, C.W. Effects of nutrients and light intensity on the growth and biochemical composition of a marine microalga Odontella aurita. Chin. J. Oceanol. Limnol. 2013, 31, 1163–1173. [Google Scholar] [CrossRef]
- Størseth, T.R.; Kirkvold, S.; Skjermo, J.; Reitan, K.I. A branched β-d-(1→3,1→6)-glucan from the marine diatom Chaetoceros debilis (Bacillariophyceae) characterized by NMR. Carbohydr. Res. 2006, 341, 2108–2114. [Google Scholar] [PubMed]
- Muramatsu, D.; Iwai, A.; Aoki, S.; Uchiyama, H.; Kawata, K.; Nakayama, Y.; Nikawa, Y.; Kusano, K.; Okabe, M.; Miyazaki, T.; et al. β-glucan derived from Aureobasidium pullulans is effective for the prevention of influenza in mice. PLoS One 2012, 7, e41399. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Anjum, F.M.; Zahoor, T.; Nawaz, H.; Dilshad, S.M. Beta glucan: A valuable functional ingredient in foods. Crit. Rev. Food Sci. Nutr. 2012, 52, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Bhanja, S.K.; Nandan, C.K.; Mandal, S.; Bhunia, B.; Maiti, T.K.; Mondal, S.; Islam, S.S. Isolation and characterization of the immunostimulating β-glucans of an edible mushroom Termitomyces robustus var. Carbohydr. Res. 2012, 357, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Hu, Z.; Fu, H.; Hu, M.; Xu, X.; Chen, J. Chemical analysis and antioxidant activity in vitro of a β-d-glucan isolated from Dictyophora indusiata. Int. J. Biol. Macromol. 2012, 51, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Braud, J.P. Simultaneous culture in pilot tanks of the macroalga Chondrus crispus (Gigartinaceae) and the microalga Odontella aurita (Eupodiscaceae) producing EPA. In Marine Microorganisms for Industry; Le Gal, Y., Muller-Feuga, A., Eds.; Ifremer Edition: Plouzane, France, 1998; pp. 39–47. [Google Scholar]
- Mimouni, V.; Ulmann, L.; Pasquet, V.; Mathieu, M.; Picot, L.; Bougaran, G.; Cadoret, J.P.; Morant-Manceau, A.; Schoefs, B. The potential of microalgae for the production of bioactive molecules of pharmaceutical interest. Curr. Pharm. Biotechnol. 2012, 13, 2733–2750. [Google Scholar] [CrossRef] [PubMed]
- Toume, K.; Ishibashi, M. 5α,8α-Epidioxysterol sulfate from a diatom Odontella aurita. Phytochemistry 2002, 61, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Moreau, D.; Tomasoni, C.; Cathrine, J.; Kaas, R.; Guedes, L.; Cadoret, J.P.; Muller-Feuga, A.; Kontiza, I.; Vagias, C.; Roussis, V.; et al. Cultivated microalgae and the carotenoid fucoxanthin from Odontella aurita as potent anti-proliferative agents in bronchopulmonary and epithelial cell lines. Environ. Toxicol. Pharmacol. 2006, 22, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Wang, K.; Wan, L.L.; Li, A.F.; Hu, Q.; Zhang, C.W. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, P.; Jia, F.; Tang, J.; Ma, F. Optimization of polysaccharides from Panax japonicus C.A. Meyer by RSM and its anti-oxidant activity. Int. J. Biol. Macromol. 2012; 50, 331–336. [Google Scholar]
- You, Q.H.; Yin, X.L.; Zhang, S.N.; Jiang, Z.H. Extraction, purification, and antioxidant activities of polysaccharides from Tricholoma mongolicum Imai. Carbohydr. Polym. 2014, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Du, Y.; Li, Y.; Yang, J.; Zhang, L. Preparation and in vitro antioxidant activity of lacquer polysaccharides with low molecular weights and their sulfated derivatives. Int. J. Biol. Macromol. 2010, 46, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Tang, J.; Cao, M.; Guo, C.X.; Zhang, X.; Zhong, J.; Zhang, J.; Sun, Q.; Feng, S.; Yang, Z.R.; et al. Structure elucidation and antioxidant activity of a novel polysaccharide isolated from Tricholoma matsutake. Int. J. Biol. Macromol. 2010, 47, 271–275. [Google Scholar]
- Zhang, Y.X.; Dai, L.; Kong, X.W.; Chen, L.W. Characterization and in vitro antioxidant activities of polysaccharides from Pleurotus ostreatus. Int. J. Biol. Macromol. 2012, 51, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, Y.; Fujiwara, S.; Suzuki, M.; Akiyama, T.; Sakamoto, M.; Kobayashi, S.; Tsuzuki, M. Structural and physiological studies on the storage β-polyglucan of haptophyte Pleurochrysis haptonemofera. Planta 2008, 227, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.I.; Kim, H.J.; Lee, J.W. Structural feature and antioxidant activity of low molecular weight laminarin degraded by gamma irradiation. Food Chem. 2011, 129, 520–523. [Google Scholar] [CrossRef]
- Størseth, T.R.; Hansen, K.; Skjermo, J.; Krane, J. Characterization of a β-d-(1→3)-glucan from the marine diatom Chaetoceros mülleri by high-resolution magic-angle spinning NMR spectroscopy on whole algal cells. Carbohydr. Res. 2004, 339, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Niksic, M.; Vrvic, M.M.; Todorovic, N.; Jakovljevic, D.; van Griensven, L.J.L.D. Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor. J. Food Compos. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Lo, T.C.-T.; Chang, C.A.; Chiuc, K.-H.; Tsayd, P.-K.; Jena, J.-F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Kanmani, P.; Kumar, R.S.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Arul, V. Production and purification of a novel exopolysaccharide from lactic acid bacterium Streptococcus phocae PI80 and its functinoal characteristics activity invitro. Bioresour. Technol. 2011, 102, 4827–4833. [Google Scholar] [CrossRef] [PubMed]
- Ker, Y.-B.; Chen, K.-C.; Chyau, C.-C.; Chen, C.-C.; Guo, J.-H.; Hsien, C.-L.; Wang, H.-E.; Peng, C.-C.; Chang, C.-H.; Peng, R.-P.; et al. Antioxidant capability of polysaccharides fractionated from submerge-cultured Agaricus blazei Mycelia. J. Agric. Food Chem. 2005; 53, 7052–7058. [Google Scholar]
- Li, J.; Liu, Y.; Fan, L.; Ai, L.; Shan, L. Antioxidant activities of polysaccharides from the fruiting bodies of Zizyphus jujuba cv. Jinsixiaozao. Carbohydr. Polym. 2011, 84, 390–394. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; Brabanter, J.D.; Cooman, L.D. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Guedes, A.C.; Giao, M.S.; Seabra, R.; Ferreira, A.C.S.; Tamagnini, P.; Moradas-Ferreira, P.; Malcata, F.X. Evaluation of the antioxidant activity of cell extracts from microalgae. Mar. Drugs 2013, 11, 1256–1270. [Google Scholar] [CrossRef] [PubMed]
- Myklestad, S.M. Production, chemical structure, metabolism, and biological function of the (1→3)-linked, β-d-glucans in diatoms. Biol. Oceanogr. 1989, 6, 313–326. [Google Scholar]
- Granum, E.; Myklestad, S.M. A simple combined method for determination of β-1,3-glucan and cell wall polysaccharides in diatoms. Hydrobiologia 2002, 477, 155–161. [Google Scholar] [CrossRef]
- Dubois, M.; Gillies, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for the determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar]
- Ye, M.; Chen, W.X.; Qiu, T.; Yuan, R.Y.; Ye, Y.W.; Cai, J.M. Structural characterization and anti-ageing activity of extracellular polysaccharide from a strain of Lachnum sp. Food Chem. 2012, 132, 338–343. [Google Scholar] [CrossRef]
- Sachindra, N.M.; Sato, E.; Maeda, H.; Hosokawa, M.; Niwano, Y.; Kohno, M.; Miyashita, K. Radical scavenging and singlet oxygen quenching activity of marine carotenoid fucoxanthin and its metabolites. J. Agric. Food Chem. 2007, 55, 8516–8522. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella aurita. Mar. Drugs 2014, 12, 4883-4897. https://doi.org/10.3390/md12094883
Xia S, Gao B, Li A, Xiong J, Ao Z, Zhang C. Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella aurita. Marine Drugs. 2014; 12(9):4883-4897. https://doi.org/10.3390/md12094883
Chicago/Turabian StyleXia, Song, Baoyan Gao, Aifen Li, Jihai Xiong, Ziqiang Ao, and Chengwu Zhang. 2014. "Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella aurita" Marine Drugs 12, no. 9: 4883-4897. https://doi.org/10.3390/md12094883
APA StyleXia, S., Gao, B., Li, A., Xiong, J., Ao, Z., & Zhang, C. (2014). Preliminary Characterization, Antioxidant Properties and Production of Chrysolaminarin from Marine Diatom Odontella aurita. Marine Drugs, 12(9), 4883-4897. https://doi.org/10.3390/md12094883