
Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes

Abstract
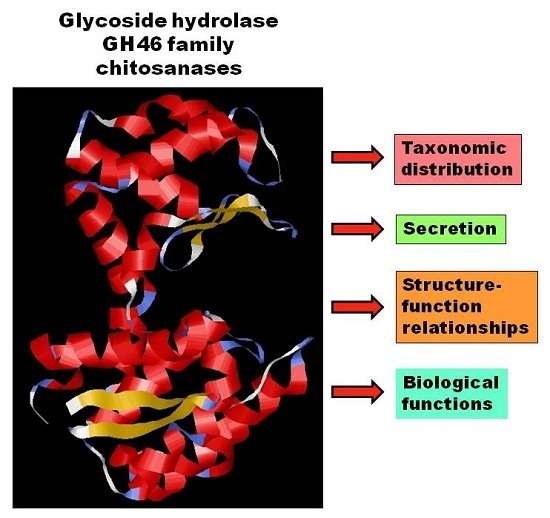
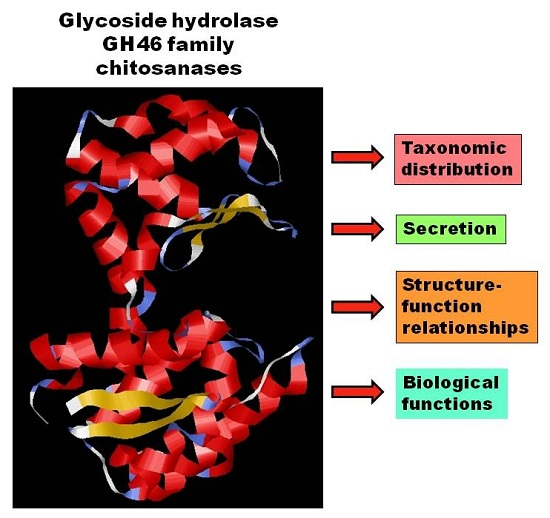
:
1. Introduction
1.1. Why Chitosan?
1.2. Why Chitosanases?
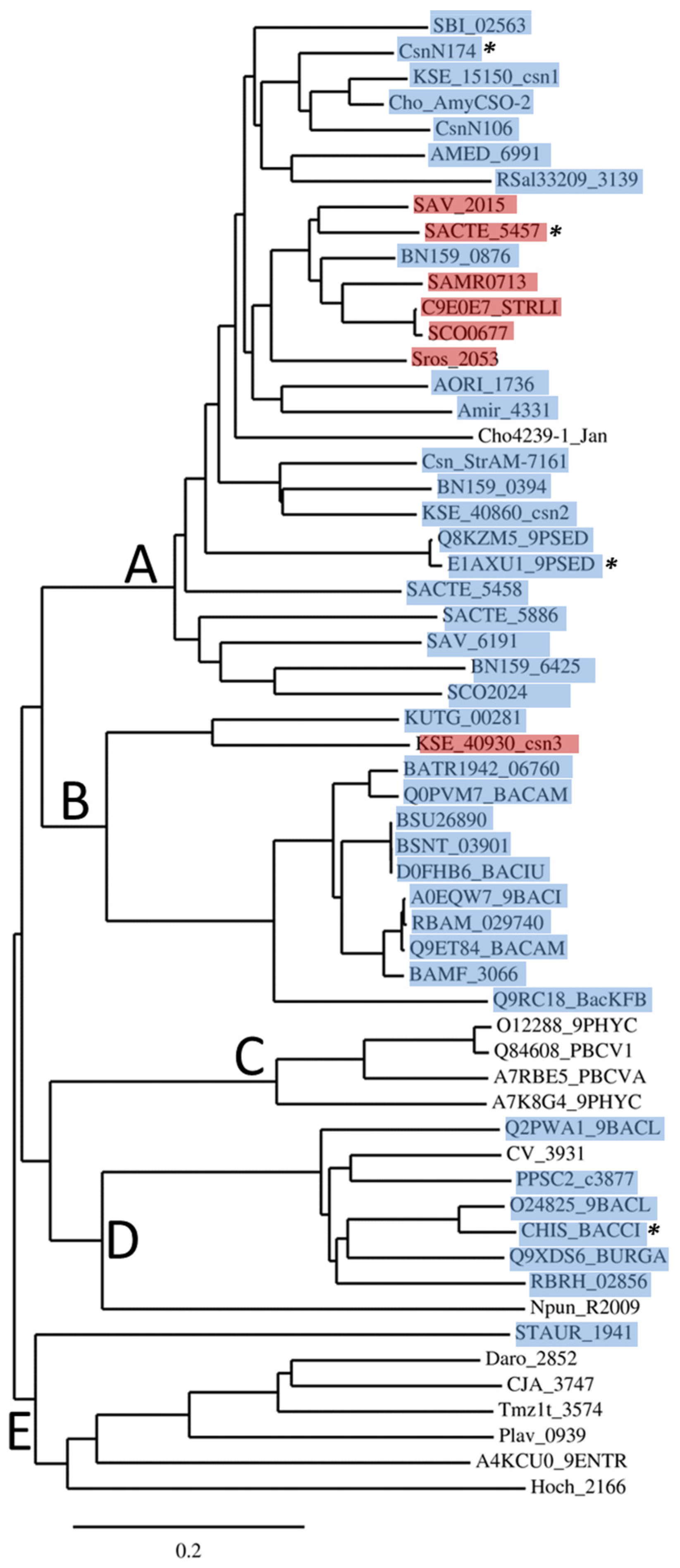
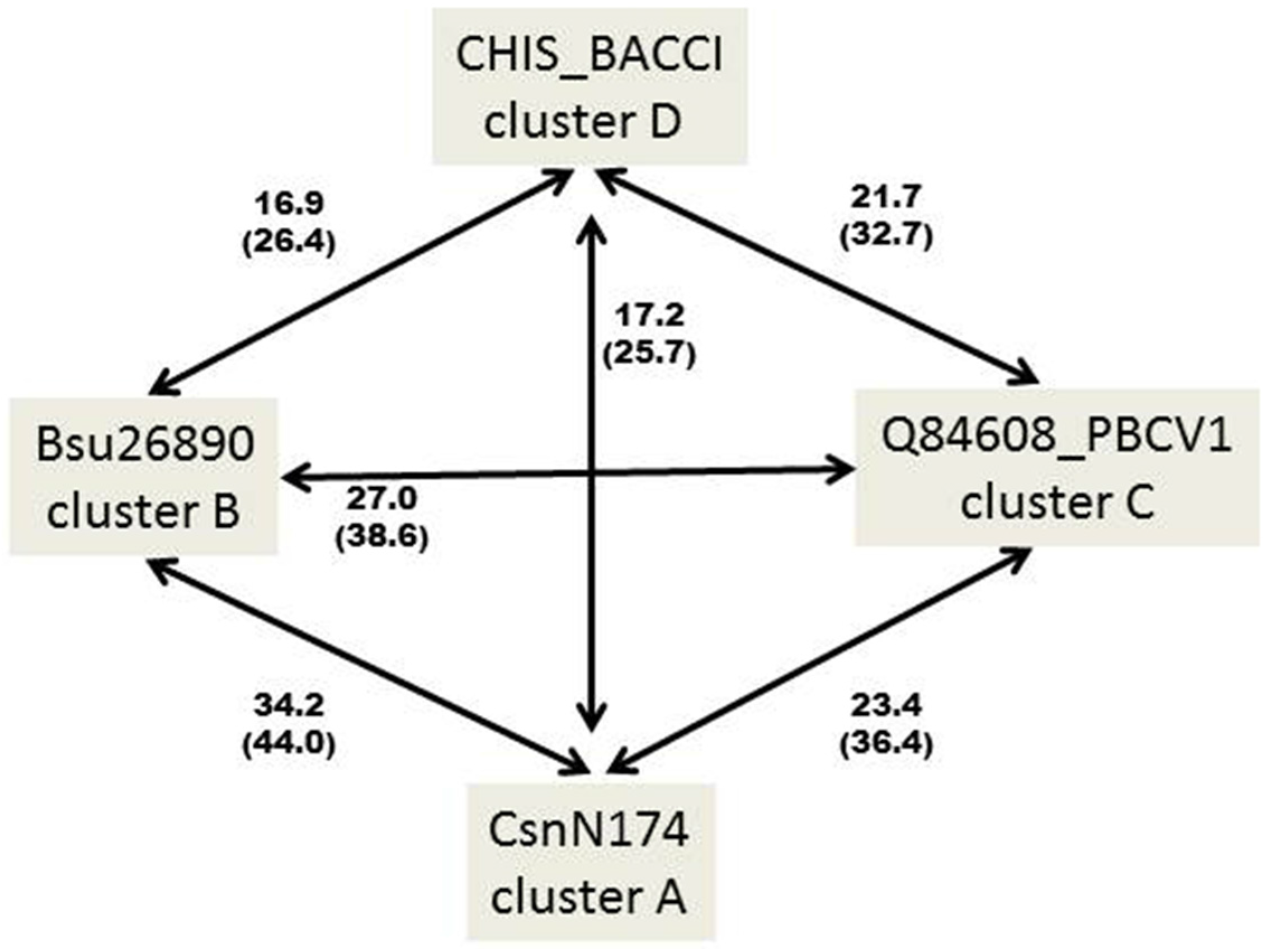
2. GH46 Family Proteins: Phylogenetic Tree and Taxonomic Distribution


3. Multimodularity in GH46 Chitosanases
3.1. Signal Peptides and Secretion of Chitosanases
3.2. Other Modules
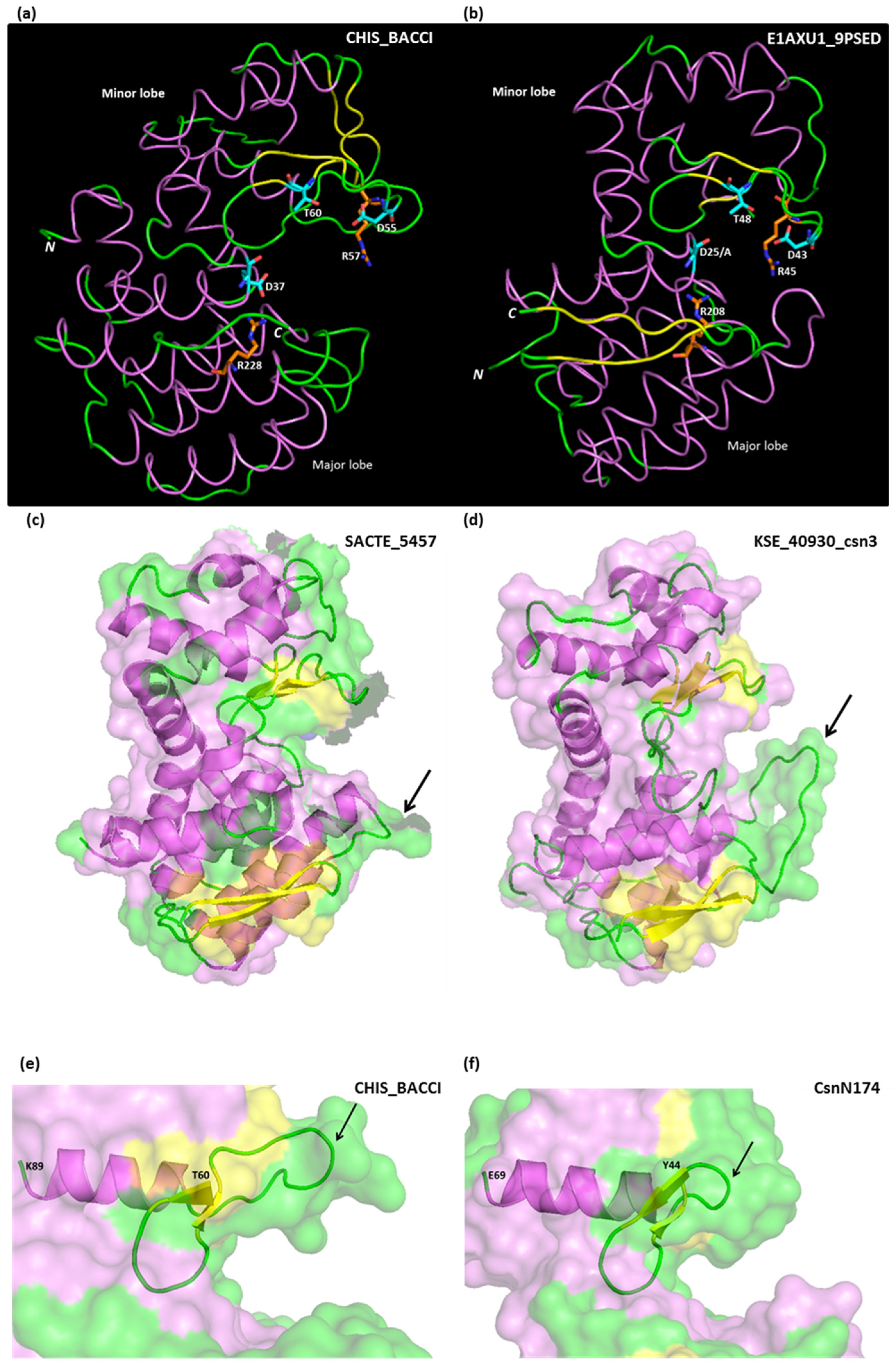
4. Structure–Function Relationships: Summary of Results from Crystallography and Site-Directed Mutagenesis
4.1. Tertiary Structure and Key Residues


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. General Function Residues | |||
| CsnN174 | SACTE_5457 | CHIS_BACCI | OU01 |
| L5 Interlobe hydrophobic interaction | L57 | L21 | L8 |
| E22 * [52] Catalytic general acid | E74 # [27] Catalytic general acid | E37 # [41] Catalytic general acid | E25 * [33,53] Catalytic general acid |
| W28 * [54] Cooperative stabilization of the protein structure via hydrophobic and carboxyl side chains interaction | W80 | W43 | W31 |
| No equivalent | No equivalent | C50 # [41] Disulfide bridge with C124 | No equivalent |
| D40 * [52] Catalytic nucleophile | D92 # [27] Catalytic nucleophile | D55 * [41] Catalytic nucleophile | D43 # [33,53] Catalytic nucleophile |
| R42 * [49] Electrostatic interaction with the catalytic nucleophile | R94 | R57 * [55] Deprotonation of the catalytic nucleophile | R45 # [33] |
| T45 * [51] Water molecule positioning | T97 † [27] | T60 | T48 # [33,53] Water molecule positioning |
| F97 † [54] Hydrophobic interaction network with W101 | F149 | F123 | F100 |
| No equivalent | No equivalent | C124 # [41] Disulfide bridge with C50 | No equivalent |
| W101 * [54] Stabilization of the protein structure via hydrophobic interaction with F97 | W153 | I127 | W104 |
| D145 † [56] Member of ionic interaction network that stabilizes the catalytic cleft with R190 and R205 | D197 | D172 † [56] Member of ionic interaction network that stabilizes the catalytic cleft with R210 and R228 | D148 |
| R190 † [56] Member of ionic interaction network that stabilizes the catalytic cleft with D145 and R205 | R242 | R210 † [56] Member of ionic interaction network that stabilizes the catalytic cleft with D172 and R228 | R193 |
| R205 *,† [56] Member of ionic interaction network that stabilizes the catalytic cleft with D145 and R190. Also in direct interaction with the general acid catalytic residue. | R257 | R228 † [56] Member of ionic interaction network that stabilizes the catalytic cleft with D172 and R210. Also in direct interaction with the general acid catalytic residue. | R208 |
| W227 * [54] Cooperative stabilization of the protein structure via hydrophobic and carboxyl side chains interaction | W279 | No equivalent | W230 |
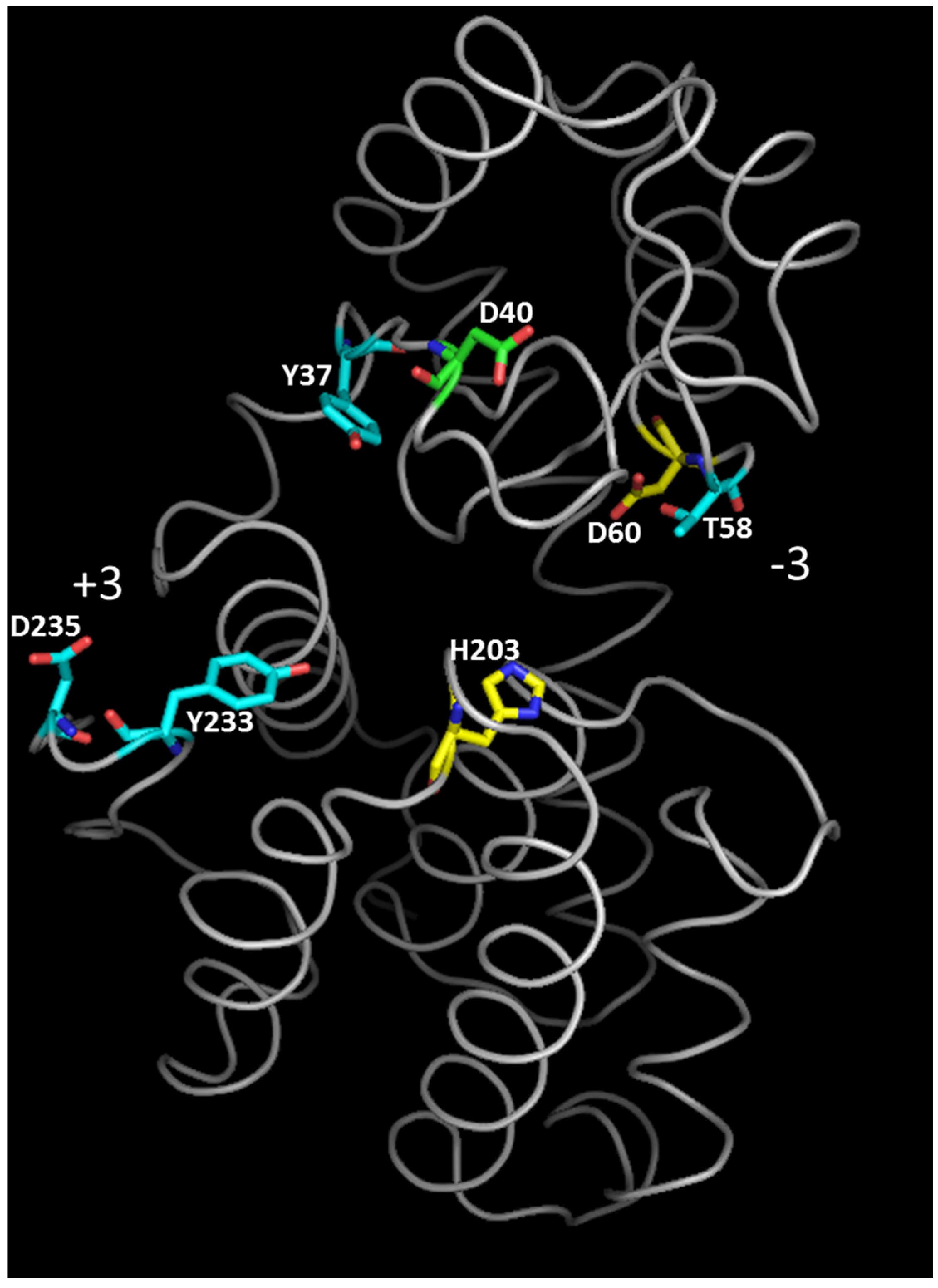
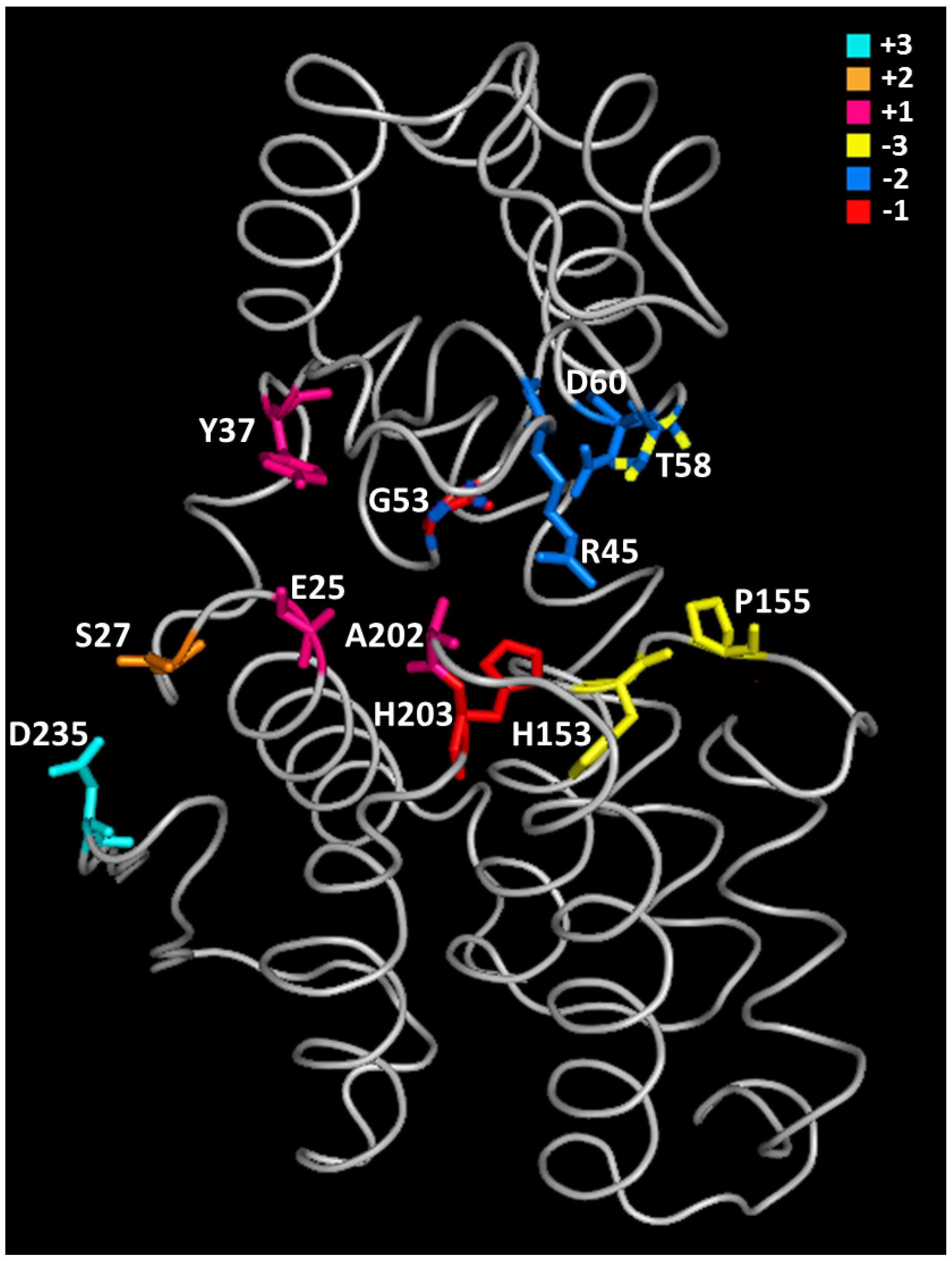
| B. Substrate Interaction Residues (Subsite Indicated into Brackets) § | |||
| CsnN174 | SACTE_5457 § | CHIS_BACCI | OU01 |
| E22 | E74 | E37 | E25 # [53] (+1) |
| N23 | N75 † [27] (+1) | Q38 | N26 |
| S24 | S76 | D39 | S27 *,# [33] (+2) |
| Q31 | Q83 † [27] (+1) | Y46 | Q34 |
| K33 | G85 † [27] Accommodation of an acetyl group of GlcNAc at (+1) | G48 | G36 |
| Y34 † [32] (+1) | Y86 † [27] (+1) Accommodation of an acetyl group of GlcNAc at (+1) | Y49 | Y37 *,# [33] (+1) |
| R42 * [49] Electrostatic interaction with substrate | R94 † [27] (−2) | R57 * [55] | R45 # [33] (−2) Hydrogen bond with substrate. |
| T45 * [51] | T97 † [27] (−1) | T60 | T48 # [33] |
| G46 | A98 † [27] b Accommodation of an acetyl group of GlcNAc at (+1) | I61 | G49 |
| G47 | G99 † [27] Accommodation of an acetyl group of GlcNAc at (+1) | G62 | G50 |
| I49 † [32] (−2) | I101 † [27] Interference with an acetyl group of GlcNAc at (−2) | F64 | I52 # [33] |
| G50 † [32] (−1) | G102 | G65 | G53 # [33] (−2 and −1) |
| T55 | T107 † [27] (−2) | H75 | T58 *,# [33] (−2 and −3) |
| D57 * [57,58] (−2) | D109 † [27]( −2) | D77 | D60 # [33] (−2) |
| Y122 † [32] (−2) | Y174 † [27] (−2) Interference with an acetyl group of GlcNAc at (−2) | Y148 * [59] (−2) | Y125 |
| H150 | H202 | N177 | H153 *,# [33] (−3) |
| P152 † [32] (−2) | G204 | A179 | P155 # [33] (−3) |
| E197 †,* [32,58] (−1) | E249 | N217 | E200 *,# [33] Hydrogen bond with R45 |
| A199 | A251 | Y219 | A202 # [33] (+1) |
| H200 | H252 | N220 | H203 # [33] (−1) |
| D201 †,* [32,58] (+2) | S253 | K221 | A204 |
| D232 | D284 | T259 | D235 # [33] (+3) |
4.2. Substrate Binding and Cleavage


5. Biological Functions of GH46 Chitosanases
5.1. Metabolic Assimilation of Chitosan
5.2. Protection against the Antimicrobial Activity of Chitosan
5.3. Chitosanase-Aided Lysis of Algal Cell Wall as a Step in Viral Development
5.4. Antifungal Effect
Supplementary Files
Supplementary File 1Acknowledgments
Conflicts of Interest
References
- Aranaz, I.; Mengíbar, M.; Harris, R.; Paños, I.; Miralles, B.; Acosta, N.; Galed, G.; Heras, A. Functional characterization of chitin and chitosan. Curr. Chem. Biol. 2009, 3, 203–230. [Google Scholar]
- Muzzarelli, R.A.A.; Boudrant, J.; Meyer, D.; Manno, N.; DeMarchis, M.; Paoletti, M.G. Current views on fungal chitin/chitosan, human chitinases, food preservation, glucans, pectins and inulin: A tribute to Henri Braconnot, precursor of the carbohydrate polymers science, on the chitin bicentennial. Carbohydr. Polym. 2012, 87, 995–1012. [Google Scholar] [CrossRef]
- Pillai, C.K.S.; Paul, W.; Sharma, C.P. Chitosan: Manufacture, properties and uses. In Chitosan: Manufacture, Properties and Usage; Davis, S.P., Ed.; Nova Publishers: Hauppauge, NY, USA, 2011; pp. 133–216. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. (Oxf.) 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Zhang, J.; Xia, W.; Liu, P.; Cheng, Q.; Tahirou, T.; Gu, W.; Li, B. Chitosan modification and pharmaceutical/biomedical applications. Mar. Drugs 2010, 8, 1962–1987. [Google Scholar] [PubMed]
- Buschmann, M.D.; Merzouki, A.; Lavertu, M.; Thibault, M.; Jean, M.; Darras, V. Chitosans for delivery of nucleic acids. Adv. Drug Deliv. Rev. 2013, 65, 1234–1270. [Google Scholar] [CrossRef] [PubMed]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.Y.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Muzzarelli, R.A.A. Chitins and chitosans for the repair of wounded skin, nerve, cartilage and bone. Carbohydr. Polym. 2009, 76, 167–182. [Google Scholar] [CrossRef]
- Park, J.H.; Saravanakumar, G.; Kim, K.; Kwon, I.C. Targeted delivery of low molecular drugs using chitosan and its derivatives. Adv. Drug Deliv. Rev. 2010, 62, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Wan Ngah, W.S.; Teong, L.C.; Hanafiah, M.A.K.M. Adsorption of dyes and heavy metal ions by chitosan composites: A review. Carbohydr. Polym. 2011, 83, 1446–1456. [Google Scholar] [CrossRef]
- Blanchard, J.; Park, J.K.; Boucher, I.; Brzezinski, R. Industrial applications of chitosanases. In Recent Advances in Marine Biotechnology; Volume 9: Biomaterials and Bioprocessing; Fingerman, M., Nagabhushanam, R., Eds.; Science Publishers: Enfield, NH, USA, 2003; pp. 257–277. [Google Scholar]
- Thadathil, N.; Velappan, S.P. Recent developments in chitosanase research and its biotechnological applications: A review. Food Chem. 2014, 150, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Du, Y.; Zhang, J. Low molecular weight and oligomeric chitosans and their bioactivities. Curr. Top. Med. Chem. 2009, 9, 1546–1559. [Google Scholar] [CrossRef] [PubMed]
- Aam, B.B.; Heggset, E.B.; Norberg, A.L.; Sørlie, M.; Vårum, K.M.; Eijsink, V.G.H. Production of chitooligosaccharides and their potential applications in medicine. Mar. Drugs 2010, 8, 1482–1517. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Villegas, M.I.; Aam, B.B.; Rupnow, J.; Sølie, M.; Eijsink, V.G.H.; Hutkins, R.W. Adherence inhibition of enteropathogenic Escherichia coli by chitooligosaccharides with specific degrees of acetylation and polymerization. J. Agric. Food Chem. 2013, 61, 2748–2754. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhao, X.; Du, Y. Oligochitosan: A plant diseases vaccine—A review. Carbohydr. Polym. 2010, 82, 1–8. [Google Scholar] [CrossRef]
- Mourya, V.K.; Inamdar, N.N.; Choudhari, Y.M. Chitooligosaccharides: Synthesis, characterization and applications. Polym. Sci. Ser. A 2011, 7, 583–612. [Google Scholar] [CrossRef]
- Fukamizo, T. Chitinolytic enzymes: Catalysis, substrate binding, and their application. Curr. Protein Pept. Sci. 2000, 1, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Hoell, I.A.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Structure and function of enzymes acting on chitin and chitosan. Biotechnol. Genet. Eng. Rev. 2010, 27, 331–366. [Google Scholar] [CrossRef]
- Henrissat, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1991, 280, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Ando, A.; Noguchi, K.; Yanagi, M.; Shinoyama, H.; Kagawa, Y.; Hirata, H.; Yabuki, M.; Fujii, T. Primary structure of chitosanase produced by Bacillus circulans MH-K1. J. Gen. Appl. Microbiol. 1992, 38, 135–144. [Google Scholar] [CrossRef]
- Masson, J.-Y.; Denis, F.; Brzezinski, R. Primary sequence of the chitosanase from Streptomyces sp. N174 and comparison with other endoglycosidases. Gene 1994, 140, 103–107. [Google Scholar] [CrossRef]
- Henrissat, B.; Bairoch, A. Updating the sequence-based classification of glycosyl hydrolases. Biochem. J. 1996, 316, 695–696. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.J.; Sinnott, M.L. Sorting the diverse: The sequence-based classifications of carbohydrate-active enzymes. Biochem. J. 2008. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Takasuka, T.E.; Bianchetti, C.M.; Tobimatsu, Y.; Bergeman, L.F.; Ralph, J.; Fox, B.G. Structure-guided analysis of catalytic specificity of the abundantly secreted chitosanase SACTE_5457 from Streptomyces sp. SirexAA-E. Proteins 2014, 82, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Chevenet, E.; Brun, C.; Bañuls, A.L.; Jacq, B.; Christen, R. TreeDyn: Towards dynamic graphics and annotations for analyses of trees. BMC Bioinform. 2006, 7, 439. [Google Scholar] [CrossRef] [PubMed]
- Bagos, P.G.; Nikolaou, E.P.; Liakopoulos, T.D.; Tsirigos, K.D. Combined prediction of Tat and Sec signal peptides with hidden Markov models. Bioinformatics 2010, 26, 2811–2817. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jacques, P-É.; Ghinet, M.G.; Brzezinski, R.; Morosoli, R. Determining the functionality of putative Tat-dependent signal peptides in Streptomyces coelicolor A3(2) using two different reporter proteins. Microbiology 2005, 151, 2189–2198. [Google Scholar] [CrossRef] [PubMed]
- Ando, A.; Saito, A.; Arai, S.; Usuda, S.; Furuno, M.; Kaneko, N.; Shida, O.; Nagata, Y. Molecular characterization of a novel family-46 chitosanase from Pseudomonas sp. A-01. Biosci. Biotechnol. Biochem. 2008, 72, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Marcotte, E.M.; Monzingo, A.F.; Ernst, S.R.; Brzezinski, R.; Robertus, J.D. X-ray structure of an anti-fungal chitosanase from Streptomyces N174. Nat. Struct. Biol. 1996, 3, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Wang, S.; Xu, W.; Han, B.; Liu, W.; Jones, D.N.M.; Liu, W. Structural insights into the substrate-binding mechanism for a novel chitosanase. Biochem. J. 2014, 461, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Heggset, E.B.; Dybvik, A.I.; Hoell, I.A.; Norberg, A.L.; Sørlie, M.; Eijsink, V.G.H.; Vårum, K.M. Degradation of chitosans with a family 46 chitosanase from Streptomyces coelicolor A3(2). Biomacromolecules 2010, 11, 2487–2497. [Google Scholar] [CrossRef] [PubMed]
- Ghinet, M.G.; Roy, S.; Poulin-Laprade, D.; Lacombe-Harvey, M.-È.; Morosoli, M.; Brzezinski, R. Chitosanase from Streptomyces coelicolor A3(2): Biochemical properties and role in protection against antibacterial effect of chitosan. Biochem. Cell Biol. 2010, 88, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Ooya, T.; Miyatsuchi, D.; Fuchigami, H.; Terakado, K.; Nakayama, S.-Y.; Watanabe, T.; Nagata, Y.; Ando, A. Molecular characterization and antifungal activity of a family 46 chitosanase from Amycolatopsis sp. CsO-2. FEMS Microbiol. Lett. 2009, 293, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Rivas, L.A.; Parro, V.; Moreno-Paz, M.; Mellado, R.P. The Bacillus subtilis 168 csn gene encodes a chitosanase with similar properties to a Streptomyces enzyme. Microbiology 2000, 146, 2929–2936. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.L.; Meints, R.H. Giant viruses infecting algae. Annu. Rev. Microbiol. 1999, 53, 447–494. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Chuchird, N.; Kawasaki, T.; Nishida, K.; Hiramatsu, S. Chlorella viruses as a source of novel enzymes. J. Biosci. Bioeng. 1999, 88, 353–361. [Google Scholar] [CrossRef]
- Sun, L.; Adams, B.; Gurnon, J.R.; Ye, Y.; van Etten, J.L. Characterization of two chitinase genes and one chitosanase gene encoded by Chlorella virus PBCV-1. Virology 1999, 263, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Saito, J.; Kita, A.; Higuchi, Y.; Nagata, Y.; Ando, A.; Miki, K. Crystal structure of chitosanase from Bacillus circulans MH-K1 at 1.6-Å resolution and its substrate recognition mechanism. J. Biol. Chem. 1999, 274, 30818–30825. [Google Scholar] [CrossRef] [PubMed]
- Linde, J.; Schwartze, V.; Binder, U.; Lass-Flörl, C.; Voigt, K.; Horn, F. De novo whole-genome sequence and genome annotation of Lichtheimia ramosa. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 29, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Sargent, F. The twin-arginine transport system: Moving folded proteins across membranes. Biochem. Soc. Trans. 2007, 35, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, H.; Akamatsu, M.; Fujii, Y.; Tatsumi, H.; Kusaoke, H.; Taketo, A. Discoidin domain of chitosanase is required for binding to the fungal cell wall. J. Mol. Microbiol. Biotechnol. 2010, 18, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Shinya, S.; Ohnuma, T.; Yamashiro, R.; Kimoto, H.; Kusaoke, H.; Anbazhagan, P.; Juffer, A.H.; Fukamizo, T. The first identification of carbohydrate binding modules specific to chitosan. J. Biol. Chem. 2013, 288, 30042–30053. [Google Scholar] [CrossRef] [PubMed]
- Monzingo, A.F.; Marcotte, E.M.; Hart, P.J.; Robertus, J.D. Chitinases, chitosanases, and lysozymes can be divided into prokaryotic and eukaryotic families sharing a conserved core. Nat. Struct. Biol. 1996, 3, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.A.; Kelley, L.A.; MacCallum, R.M.; Sternberg, M.J.E. Enhancement of protein modelling by human intervention in applying the automatic programs 3D-JIGSAW and 3D-PSSM. Proteins Struct. Func. Genet. Suppl. 2001, 5, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Lacombe-Harvey, M.-È.; Fortin, M.; Ohnuma, T.; Fukamizo, T.; Letzel, T.; Brzezinski, R. A highly conserved arginine residue of the chitosanase from Streptomyces sp. N174 is involved both in catalysis and substrate binding. BMC Biochem. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, M.G.; Hansen, O.C.; Stougaard, P. Isolation, characterization and heterologous expression of a novel chitosanase from Janthinobacterium sp. strain 4239. Microb. Cell Fact. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Lacombe-Harvey, M.-È.; Fukamizo, T.; Gagnon, J.; Ghinet, M.G.; Dennhart, N.; Letzel, T.; Brzezinski, R. Accessory active site residues of Streptomyces sp. N174 chitosanase. Variations on a common theme in the lysozyme superfamily. FEBS J. 2009, 276, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Boucher, I.; Fukamizo, T.; Honda, Y.; Willick, G.E.; Neugebauer, W.A.; Brzezinski, R. Site-directed mutagenesis of evolutionary conserved carboxylic amino acids in the chitosanase from Streptomyces sp. N174 reveals two residues essential for catalysis. J. Biol. Chem. 1995, 270, 31077–31082. [Google Scholar] [PubMed]
- Lyu, Q.; Shi, Y.; Wang, S.; Yang, Y.; Han, B.; Liu, W.; Jones, D.N.M.; Liu, W. Structural and biochemical insights into the degradation mechanism of chitosan by chitosanase OU01. Biochim. Biophys. Acta 2015, 1850, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Fukamizo, T.; Okajima, T.; Goto, S.; Boucher, I.; Brzezinski, R. Thermal unfolding of chitosanase from Streptomyces sp. N174: Role of tryptophan residues in the protein structure stabilization. Biochim. Biophys. Acta 1999, 1429, 365–376. [Google Scholar] [CrossRef]
- Ando, A.; Saito, A. Structure and function of chitosanase. In Advances in Chitin Science: Volume XI; Rustichelli, F., Caramella, C., Senel, S., Vårum, K.M., Eds.; The European Chitin Society: Venice, Italy, 2009; pp. 265–271. [Google Scholar]
- Fukamizo, T.; Juffer, A.H.; Vogel, H.J.; Honda, Y.; Tremblay, H.; Boucher, I.; Neugebauer, W.A.; Brzezinski, R. Theoretical calculation of pKa reveals an important role of Arg205 in the activity and stability of Streptomyces sp. N174 chitosanase. J. Biol. Chem. 2000, 275, 25633–25640. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, H.; Yamaguchi, T.; Fukamizo, T.; Brzezinski, R. Mechanism of chitosanase-oligosaccharide interaction: Subsite structure of Streptomyces sp. N174 chitosanase and the role of Asp57 carboxylate. J. Biochem. 2001, 130, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Katsumi, T.; Lacombe-Harvey, M.-È.; Tremblay, H.; Brzezinski, R.; Fukamizo, T. Role of acidic amino acid residues in chitooligosaccharide-binding to Streptomyces sp. N174 chitosanase. Biochem. Biophys. Res. Commun. 2005, 338, 1839–1844. [Google Scholar] [CrossRef] [PubMed]
- Fukamizo, T.; Amano, S.; Yamaguchi, K.; Yoshikawa, T.; Katsumi, T.; Saito, J.; Suzuki, M.; Miki, K.; Nagata, Y.; Ando, A. Bacillus circulans MH-K1 chitosanase: Amino acid residues responsible for substrate binding. J. Biochem. 2005, 138, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Fukamizo, T.; Honda, Y.; Goto, S.; Boucher, I.; Brzezinski, R. Reaction mechanism of chitosanase from Streptomyces sp. N174. Biochem. J. 1995, 311, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Rye, C.S.; Withers, S.G. Glycosidase mechanisms. Curr. Opin. Chem. Biol. 2000, 4, 573–580. [Google Scholar] [CrossRef]
- Fukamizo, T.; Ohkawa, T.; Ikeda, Y.; Goto, S. Specificity of chitosanase from Bacillus pumilus. Biochim. Biophys. Acta 1994, 1205, 183–188. [Google Scholar] [CrossRef]
- Mitsutomi, M.; Ueda, M.; Arai, M.; Ando, A.; Watanabe, T. Action patterns of microbial chitinases and chitosanases on partially N-acetylated chitosan. In Chitin Enzymology, Volume 2; Muzzarelli, R.A.A., Ed.; ATEC Edizioni: Grottamare, Italy, 1996; pp. 273–284. [Google Scholar]
- Vollmer, W.; Blanot, D.; de Pedro, A.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [PubMed]
- Lenardon, M.D.; Munro, C.A.; Gow, N.A.R. Chitin synthesis and fungal pathogenesis. Curr. Opin. Microbiol. 2010, 13, 416–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Park, R.-D.; Muzzarelli, R.A.A. Chitin deacetylases: Properties and applications. Mar. Drugs 2010, 8, 24–46. [Google Scholar] [CrossRef] [PubMed]
- Ohtakara, A. Chitosanase from Streptomyces griseus. Method Enzymol. 1988, 161, 505–510. [Google Scholar]
- Yabuki, M.; Uchiyama, A.; Suzuki, K.; Ando, A.; Fujii, T. Purification and properties of chitosanase from Bacillus circulans MH-K1. J. Gen. Appl. Microbiol. 1988, 34, 255–270. [Google Scholar] [CrossRef]
- Brzezinski, R. Uncoupling chitosanase production from chitosan. Bioeng. Bugs 2011, 2, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Viens, P.; Dubeau, M.-P.; Kimura, A.; Desaki, Y.; Shinya, T.; Shibuya, N.; Saito, A.; Brzezinski, R. Uptake of chitosan-derived d-glucosamine oligosaccharides in Streptomyces coelicolor A3(2). FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef] [PubMed]
- Kendra, D.F.; Hadwiger, L.A. Characterization of the smallest chitosan oligomer that is maximally antifungal to Fusarium solani and elicits pisatin formation in Pisum sativum. Exp. Mycol. 1984, 8, 276–281. [Google Scholar] [CrossRef]
- Tokura, S.; Ueno, K.; Miyazaki, S.; Nishi, N. Molecular weight dependent antimicrobial activity by chitosan. Macromol. Symp. 1997, 120, 1–9. [Google Scholar] [CrossRef]
- Ueno, K.; Yamaguchi, T.; Sakairi, N.; Nishi, N.; Tokura, S. Antimicrobial activity by fractionated chitosan oligomers. In Advances in Chitin Science: Volume II; Domard, A., Roberts, G.A.F., Vårum, K.M., Eds.; Jacques André Publisher: Lyon, France, 1997; pp. 156–161. [Google Scholar]
- Takeda, H. Cell wall composition and taxonomy of symbiotic Chlorella from Paramecium and Acanthocystis. Phytochemistry 1995, 40, 457–459. [Google Scholar] [CrossRef]
- Sugimoto, I.; Hiramatsu, S.; Murakami, D.; Fujie, M.; Usami, S.; Yamada, T. Algal-lytic activities encoded by Chlorella virus CVK2. Virology 2000, 277, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Hiramatsu, S.; Songsri, P.; Fujie, M. Alternative expression of a chitosanase gene produces two different proteins in cells infected with Chlorella virus CVK2. Virology 1997, 230, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Dunigan, D.D.; Cerny, R.L.; Bauman, A.T.; Roach, J.C.; Lane, L.C.; Agarkova, I.V.; Wulser, K.; Yanai-Balser, G.M.; Gurnon, J.R.; Vitek, J.C.; et al. Paramecium bursaria chlorella virus 1 proteome reveals novel architectural and regulatory features of a giant virus. J. Virol. 2012, 86, 8821–8834. [Google Scholar] [CrossRef] [PubMed]
- Jeanniard, A.; Dunigan, D.D.; Gurnon, J.R.; Agarkova, I.V.; Kang, M.; Vitek, J.; Duncan, G.; McClung, O.W.; Larsen, M.; Claverie, J.-M.; et al. Towards defining the chloroviruses: A genomic journey through a genus of large DNA viruses. BMC Genomics 2013, 14, 158. [Google Scholar] [CrossRef] [PubMed]
- Tomita, M.; Kikuchi, A.; Kobayashi, M.; Yamaguchi, M.; Ifuku, S.; Yamashoji, S.; Ando, A.; Saito, A. Characterization of antifungal activity of the GH-46 subclass III chitosanase from Bacillus circulans MH-K1. Antonie Leeuwenhoek 2013, 104, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Kouzai, Y.; Mochizuki, S.; Saito, A.; Ando, A.; Minami, E.; Nishizawa, Y. Expression of a bacterial chitosanase in rice improves disease resistance to the rice blast fungus Magnaporthe oryzae. Plant Cell Rep. 2012, 31, 629–636. [Google Scholar] [CrossRef] [PubMed]
- El Ouakfaoui, S.; Potvin, C.; Brzezinski, R.; Asselin, A. A Streptomyces chitosanase is active in transgenic tobacco. Plant Cell Rep. 1995, 15, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Kilani-Feki, O.; Frikha, F.; Zouari, I.; Jaoua, S. Heterologous expression and secretion of an antifungal Bacillus subtilis chitosanase (CSNV26) in Escherichia coli. Bioproc. Biosyst. Eng. 2013, 36, 985–992. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viens, P.; Lacombe-Harvey, M.-È.; Brzezinski, R. Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes. Mar. Drugs 2015, 13, 6566-6587. https://doi.org/10.3390/md13116566
Viens P, Lacombe-Harvey M-È, Brzezinski R. Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes. Marine Drugs. 2015; 13(11):6566-6587. https://doi.org/10.3390/md13116566
Chicago/Turabian StyleViens, Pascal, Marie-Ève Lacombe-Harvey, and Ryszard Brzezinski. 2015. "Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes" Marine Drugs 13, no. 11: 6566-6587. https://doi.org/10.3390/md13116566
APA StyleViens, P., Lacombe-Harvey, M. -È., & Brzezinski, R. (2015). Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes. Marine Drugs, 13(11), 6566-6587. https://doi.org/10.3390/md13116566