
Characterization of a Newly Isolated Marine Fungus Aspergillus dimorphicus for Optimized Production of the Anti-Tumor Agent Wentilactones


Abstract
:
1. Introduction
2. Results and Discussion
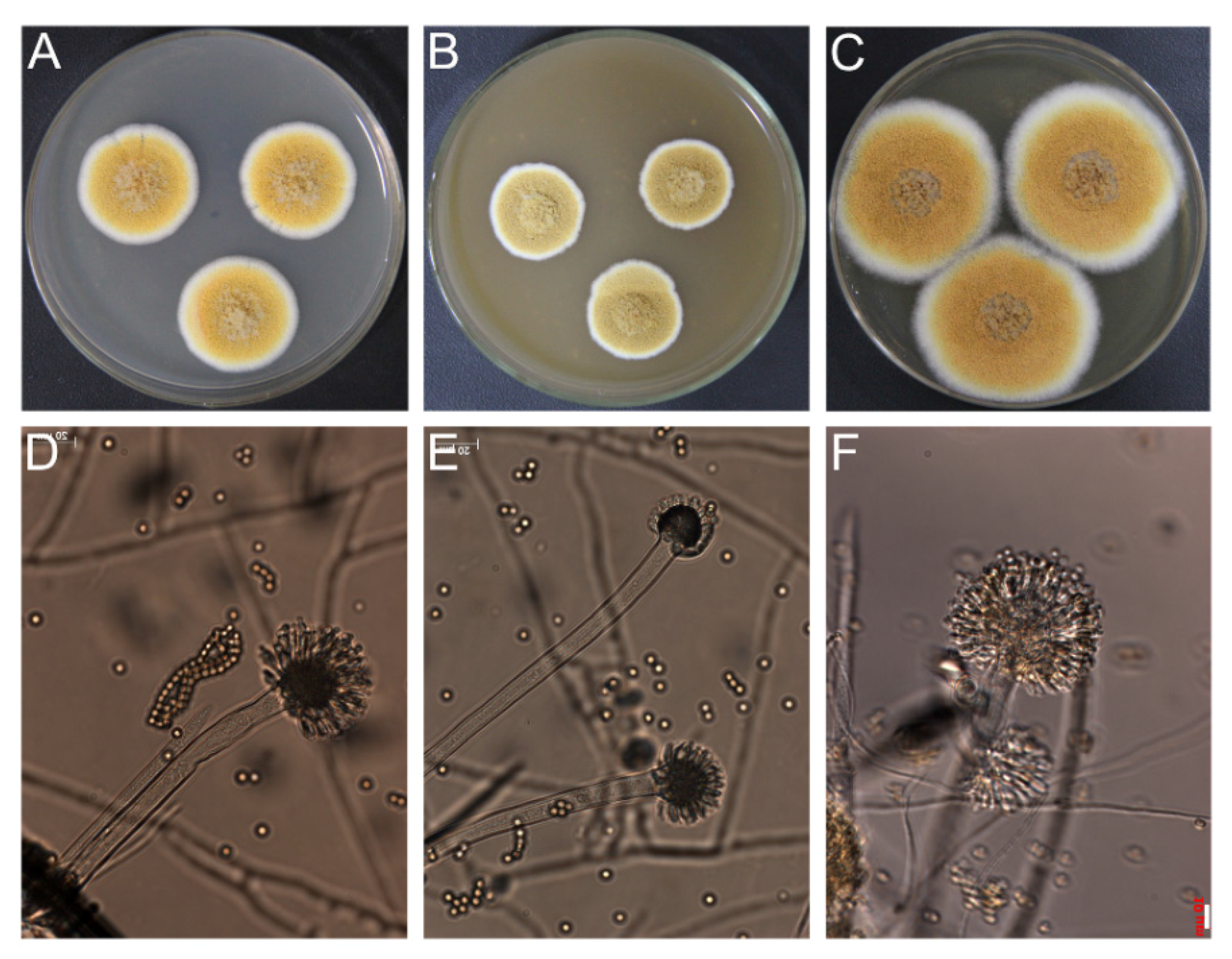
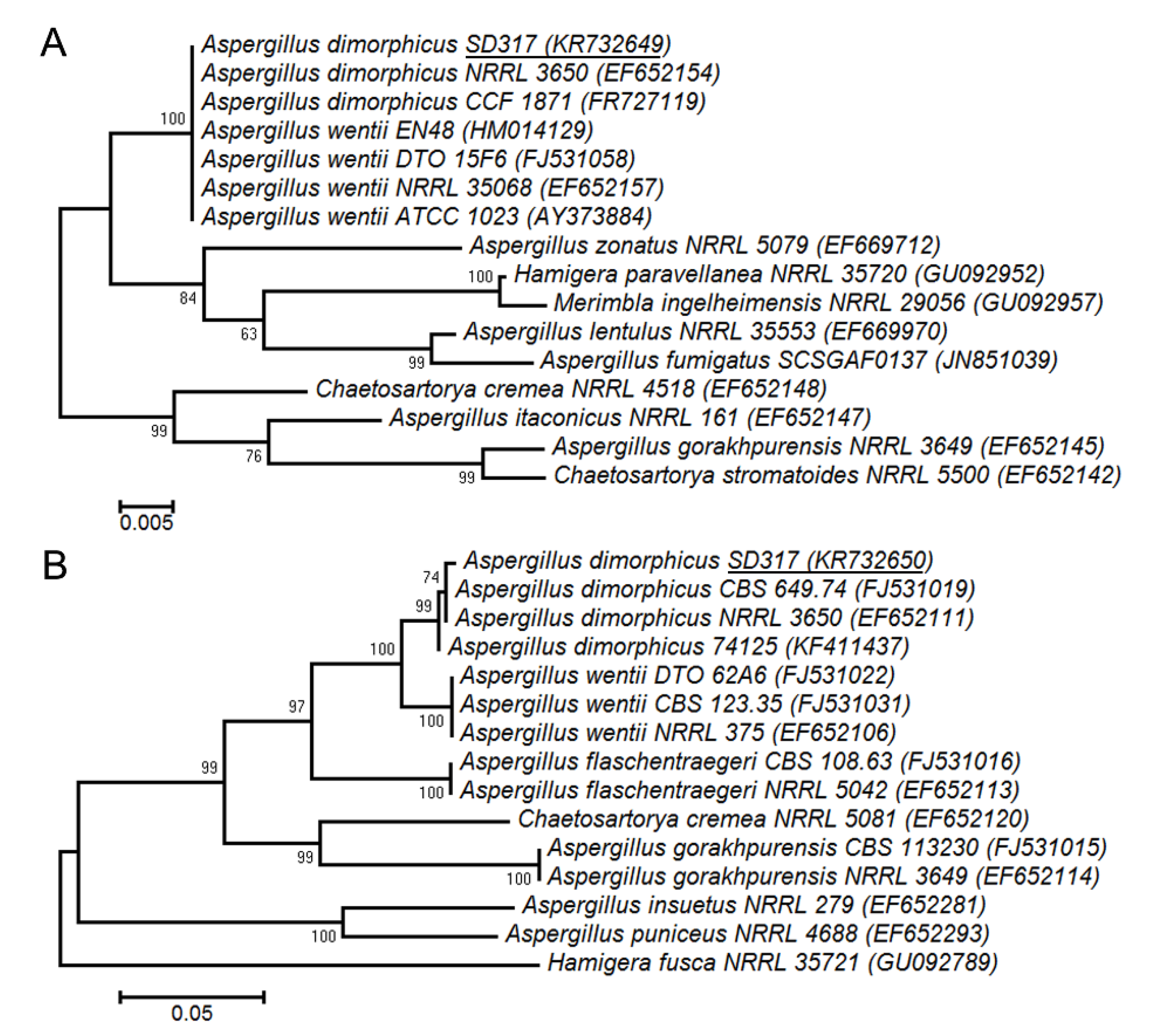
2.1. Identification of Fungal Strain by Polyphasic Approach


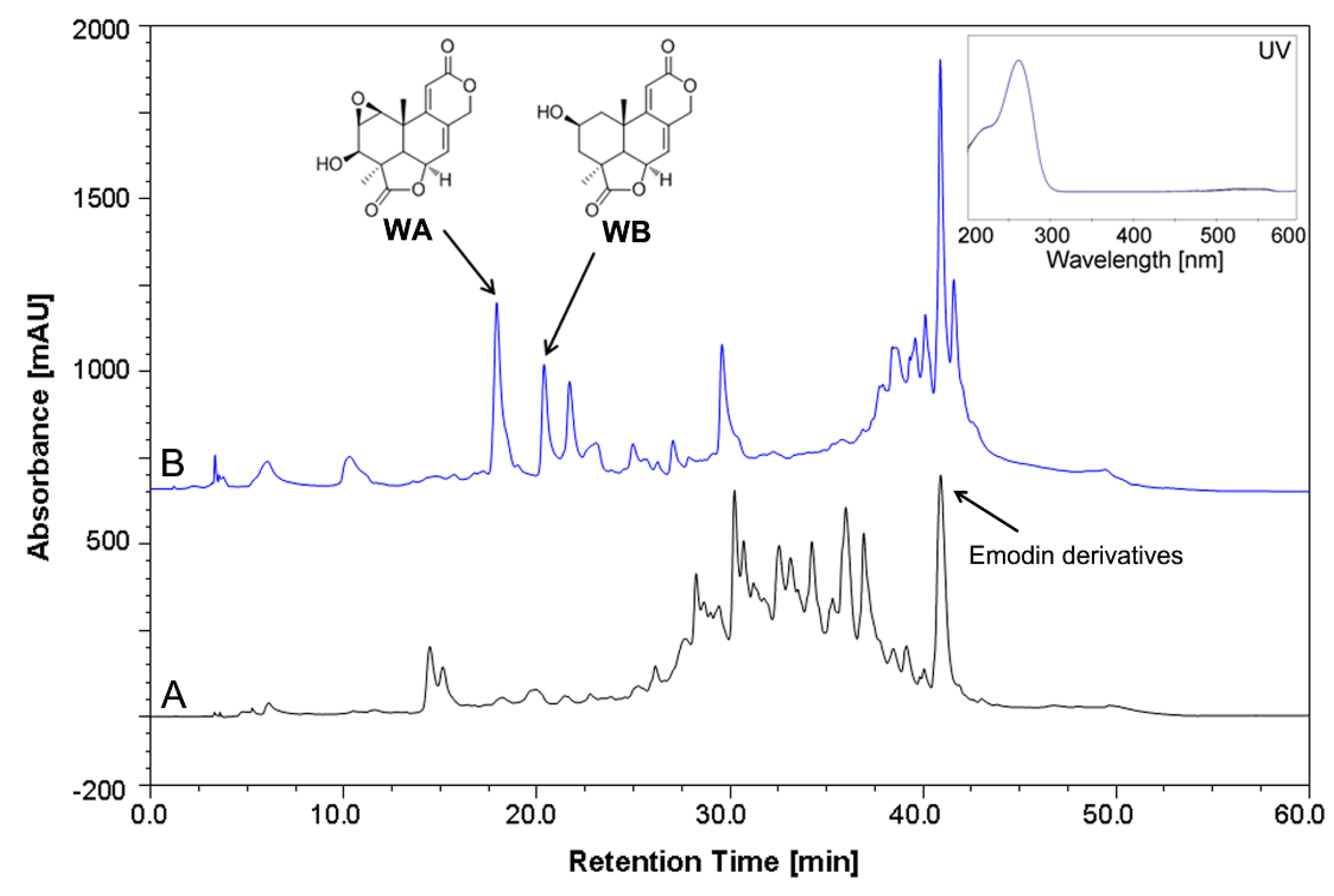
2.2. Quantitative Analysis of Wentilactone Production by HPLC

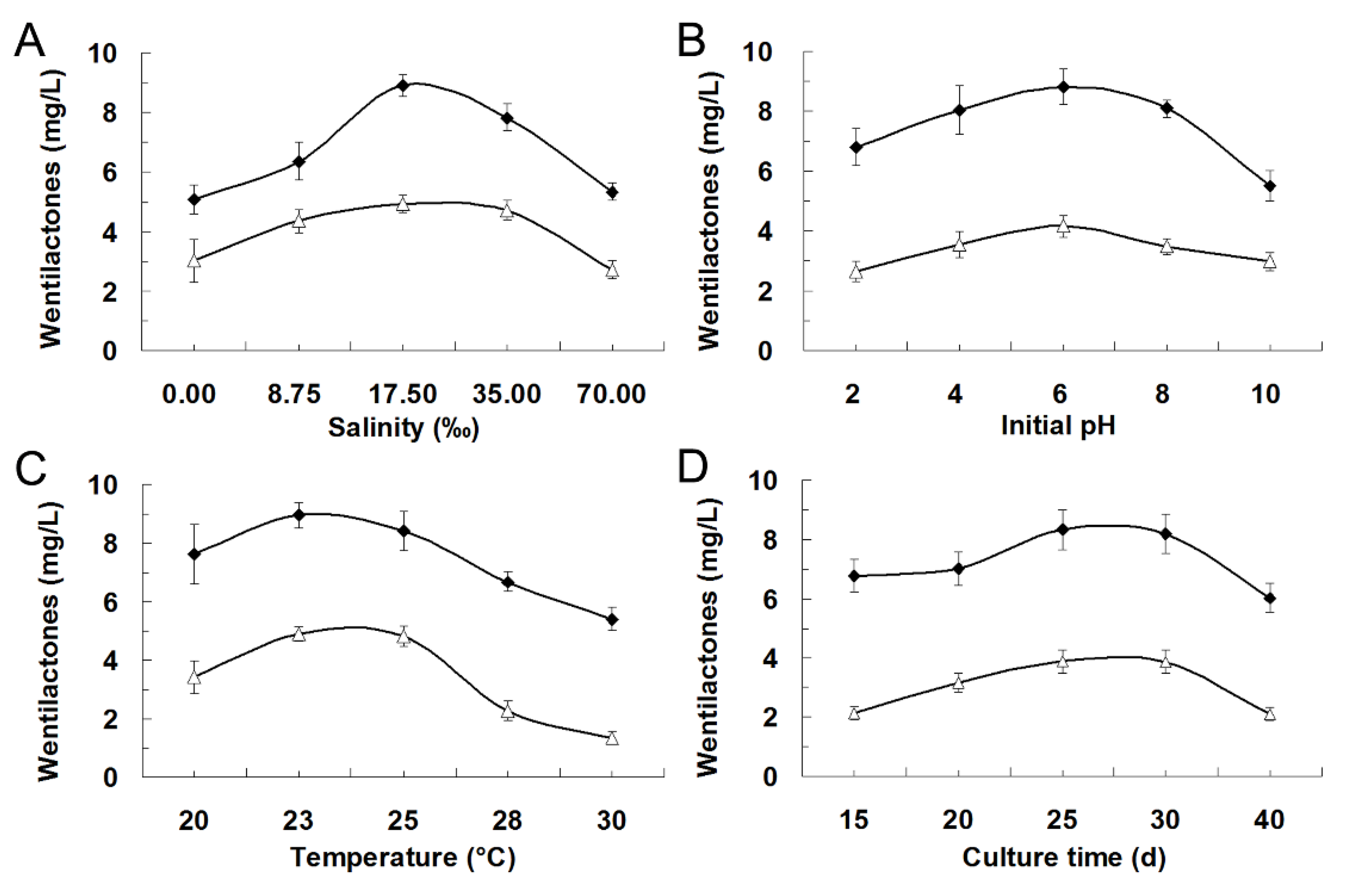
2.3. Effects of Environmental Factors on Wentilactone Production

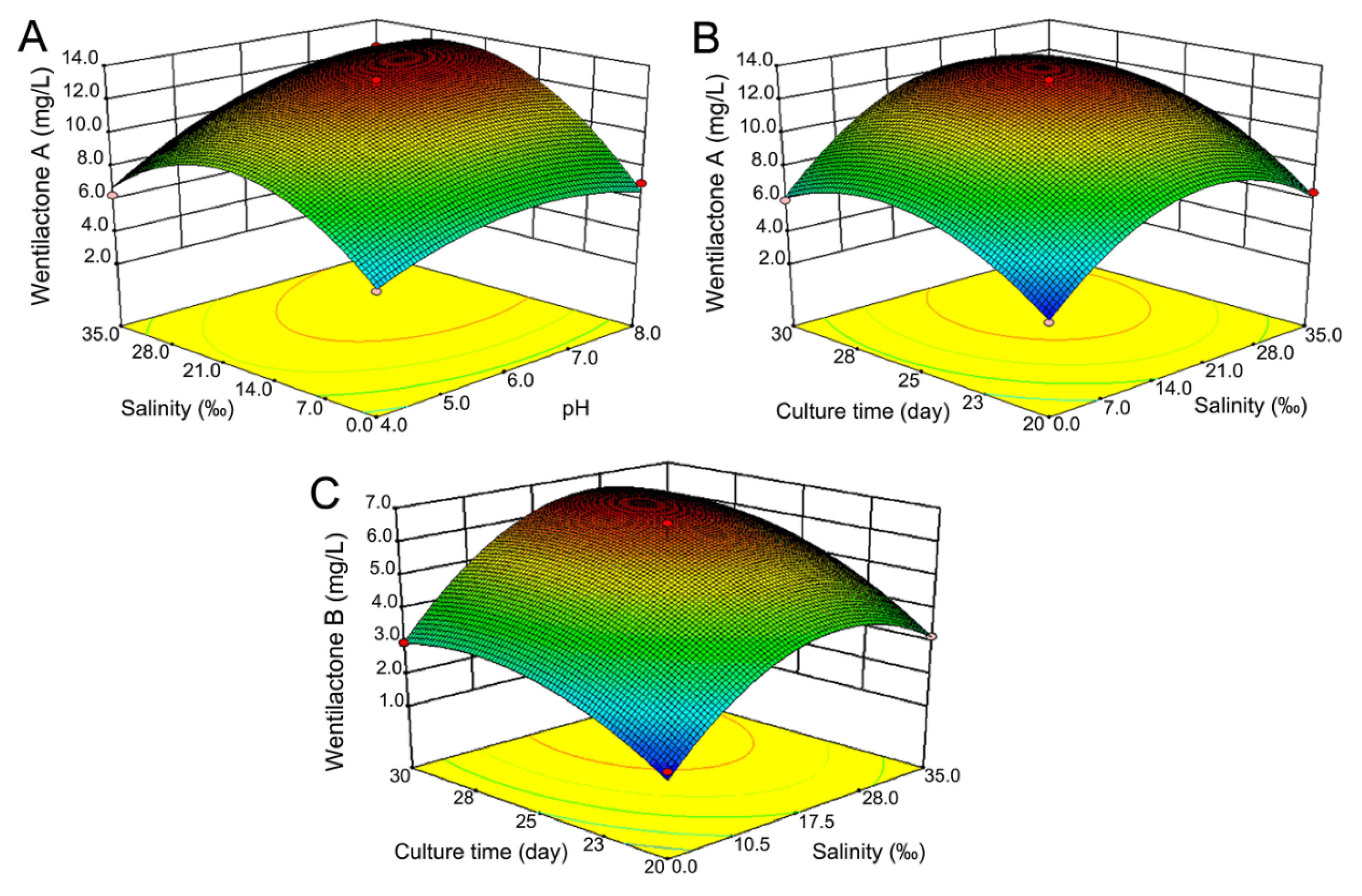
2.4. Optimization of Wentilactone Production by RSM

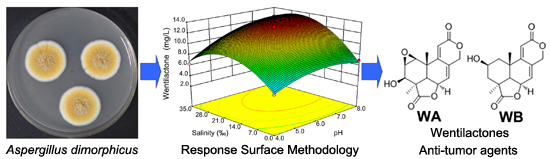{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Runing Order | Variables and Their Coded Values | Yields (mg/L) | ||||||
|---|---|---|---|---|---|---|---|---|
| pH | Salinity (‰) | Cluture Time (d) | Ya (WA) | Yb (WB) | ||||
| Code X1 | X1 | Code X2 | X2 | Code X3 | X3 | |||
| 1 | −1 | 4.0 | −1 | 0.0 | 0 | 25 | 5.16 | 1.26 |
| 2 | 1 | 8.0 | 1 | 35.0 | 0 | 25 | 12.28 | 6.21 |
| 3 | −1 | 4.0 | 1 | 35.0 | 0 | 25 | 6.28 | 3.49 |
| 4 | 0 | 6.0 | 1 | 35.0 | 1 | 30 | 10.74 | 5.64 |
| 5 | −1 | 4.0 | 0 | 17.5 | −1 | 20 | 5.66 | 2.80 |
| 6 | 0 | 6.0 | 0 | 17.5 | 0 | 25 | 12.17 | 5.77 |
| 7 | 0 | 6.0 | −1 | 0.0 | 1 | 30 | 5.96 | 2.97 |
| 8 | 1 | 8.0 | 0 | 17.5 | 1 | 30 | 11.92 | 6.53 |
| 9 | 0 | 6.0 | −1 | 0.0 | −1 | 20 | 3.43 | 1.50 |
| 10 | 1 | 8.0 | −1 | 0.0 | 0 | 25 | 7.00 | 3.16 |
| 11 | 0 | 6.0 | 0 | 17.5 | 0 | 25 | 13.15 | 6.55 |
| 12 | 0 | 6.0 | 0 | 17.5 | 0 | 25 | 12.93 | 5.83 |
| 13 | −1 | 4.0 | 0 | 17.5 | 1 | 30 | 9.09 | 4.81 |
| 14 | 1 | 8.0 | 0 | 17.5 | −1 | 20 | 8.14 | 3.62 |
| 15 | 0 | 6.0 | 1 | 35.0 | −1 | 20 | 6.44 | 3.16 |
2.4.1. Response Surface Analysis of WA
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Value | p-Value Prob > F |
|---|---|---|---|---|---|
| Model | 146.79 | 8 | 18.35 | 67.53 | <0.0001 |
| X1 | 21.64 | 1 | 21.64 | 79.63 | 0.0001 |
| X2 | 25.15 | 1 | 25.15 | 92.57 | <0.0001 |
| X3 | 24.61 | 1 | 24.61 | 90.58 | <0.0001 |
| X1X2 | 4.32 | 1 | 4.32 | 15.90 | 0.0072 |
| X2X3 | 0.78 | 1 | 0.78 | 2.89 | 0.1403 |
| X12 | 25.93 | 1 | 8.36 | 30.76 | 0.0015 |
| X22 | 86.86 | 1 | 46.92 | 172.69 | <0.0001 |
| X32 | 23.86 | 1 | 23.68 | 87.82 | <0.0001 |
| Residual | 1.63 | 6 | 0.27 | ||
| Lack of fit | 1.30 | 4 | 0.27 | 1.02 | 0.5484 |
| Pure error | 0.53 | 2 | 0.27 | ||
| Cor. total | 148.42 | 14 |

2.4.2. Response Surface Analysis of WB
| Source | Sum of Squares | Degrees of Freedom | Mean Square | F-Value | p-Value Prob > F |
|---|---|---|---|---|---|
| Model | 43.54 | 7 | 6.22 | 28.64 | 0.0001 |
| X1 | 6.42 | 1 | 6.42 | 29.57 | 0.0010 |
| X2 | 11.57 | 1 | 11.57 | 53.27 | 0.0002 |
| X3 | 9.87 | 1 | 9.87 | 45.44 | 0.0003 |
| X2X3 | 0.26 | 1 | 0.26 | 1.19 | 0.3121 |
| X12 | 1.80 | 1 | 1.80 | 8.30 | 0.0236 |
| X22 | 12.26 | 1 | 12.26 | 56.46 | 0.0001 |
| X32 | 3.07 | 1 | 3.07 | 14.11 | 0.0071 |
| Residual | 1.52 | 7 | 0.22 | ||
| Lack of fit | 1.15 | 5 | 0.23 | 1.23 | 0.5047 |
| Pure error | 0.37 | 2 | 0.19 | ||
| Cor. total | 45.06 | 14 |
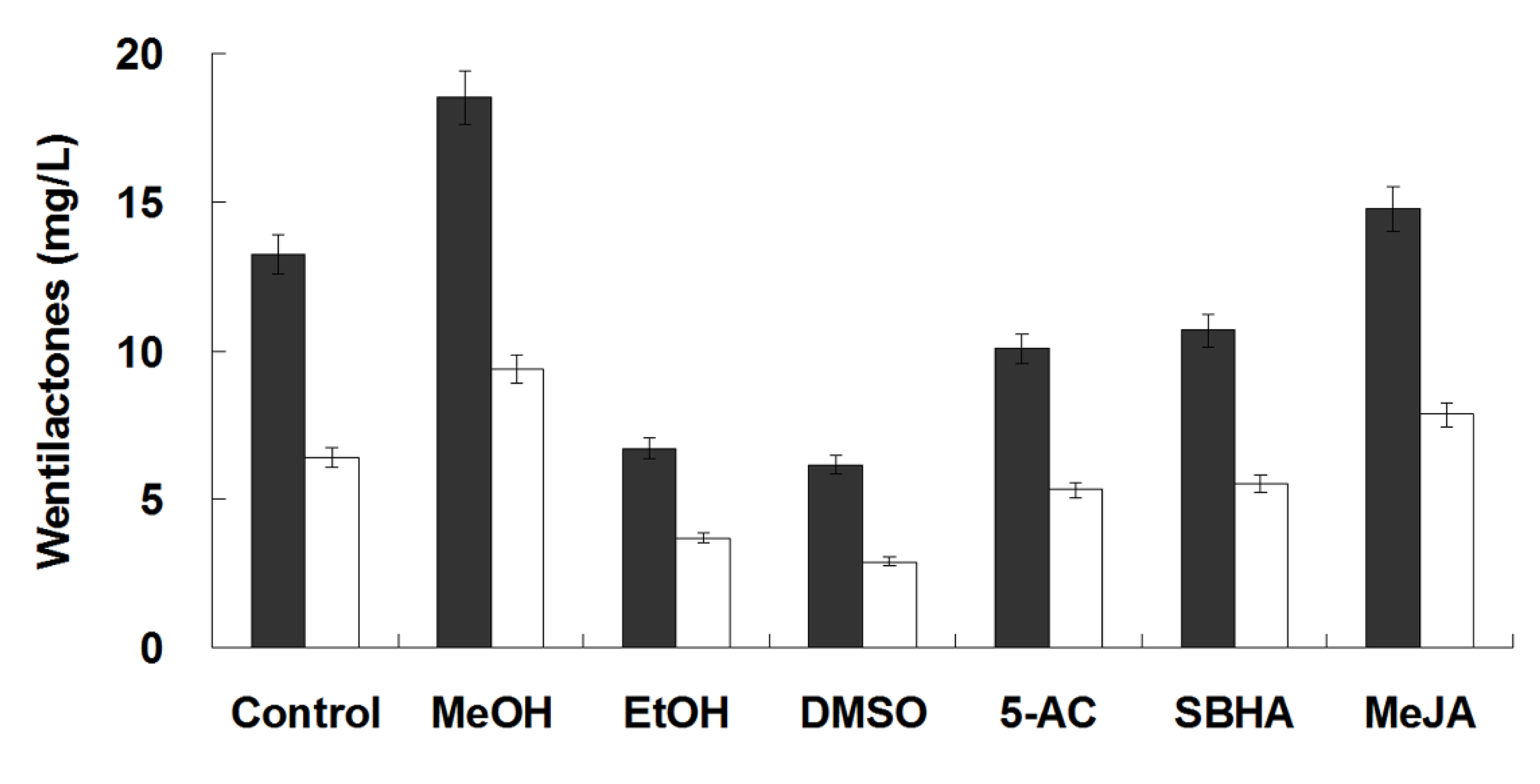
2.5. Elicitation of Wentilactone Production by Small Molecules

3. Experimental Section
3.1. Fungal Strain and Chemicals
3.2. Identification of Fungal Strain by Polyphasic Approach
3.3. Fungal Fermentation and Metabolite Extraction
3.4. Quantification of Wentilactone Production by HPLC Analysis
3.5. Effects of Environmental Factors on Wentilactone Production
3.6. Optimization of Wentilactone Production by RSM
3.7. Effects of Small-Molecule Elicitors on Wentilactone Production
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Hong, Y.; Miao, L.; Li, C.; Xu, G.; Wei, S.; Wang, B.; Huang, C.; Jiao, B. Wentilactone A as a novel potential antitumor agent induces apoptosis and G2/M arrest of human lung carcinoma cells, and is mediated by HRas-GTP accumulation to excessively activate the Ras/Raf/ERK/p53-p21 pathway. Cell Death Dis. 2013, 4, e952. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.D.; Roberts, L.R. Hepatocellular carcinoma: A global view. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Miao, L.; Sun, W.; Jiao, B.; Wang, B.; Yao, L.; Huang, C. Wentilactone B from Aspergillus wentii induces apoptosis and inhibits proliferation and migration of human hepatoma SMMC-7721 cells. Biol. Pharm. Bull. 2012, 35, 1964–1971. [Google Scholar] [CrossRef] [PubMed]
- Chabner, B.A.; Roberts, T.G.J. Timeline: Chemotherapy and the war on cancer. Nat. Rev. Cancer 2005, 5, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ajikumar, P.K.; Xiao, W.H.; Tyo, K.E.; Wang, Y.; Simeon, F.; Leonard, E.; Mucha, O.; Phon, T.H.; Pfeifer, B.; Stephanopoulos, G. Isoprenoid pathway optimization for taxol precursor overproduction in Escherichia coli. Science 2010, 330, 70–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.Q.; Yang, Y.Y.; Zhao, N.; Wang, Y. Diversity of endophytic fungi and screening of fungal paclitaxel producer from Anglojap yew, Taxus x media. BMC Microbiol. 2013, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Gomes, N.; Lefranc, F.; Kijjoa, A.; Kiss, R. Can some marine-derived fungal metabolites become actual anticancer agents? Mar. Drugs 2015, 13, 3950–3991. [Google Scholar] [CrossRef] [PubMed]
- Bromann, K.; Toivari, M.; Viljanen, K.; Vuoristo, A.; Ruohonen, L.; Nakari-Setala, T. Identification and characterization of a novel diterpene gene cluster in Aspergillus nidulans. PLoS ONE 2012, 7, e35450. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.C. Production and engineering of terpenoids in plant cell culture. Nat. Chem. Biol. 2007, 3, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Dorner, J.W.; Cole, R.J.; Springer, J.P.; Cox, R.H.; Cutler, H.; Wicklow, D.T. Isolation and identification of two new biologically-active norditerpene dilactones from Aspergillus wentii. Phytochemistry 1980, 19, 1157–1161. [Google Scholar] [CrossRef]
- Barrero, A.F.; Herrador, M.M.; Quilez del Moral, J.F.; Valdivia, M.V. A convenient synthesis of a-ring-functionalized podolactones. Revision of the structure of wentilactone B. Org. Lett. 2002, 4, 1379–1382. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.G.; Gloer, J.B.; Ji, N.Y.; Zhao, J.C. Halogenated organic molecules of Rhodomelaceae origin: Chemistry and biology. Chem. Rev. 2013, 113, 3632–3685. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.F.; Li, X.M.; Meng, L.; Cui, C.M.; Gao, S.S.; Li, C.S.; Huang, C.G.; Wang, B.G. Asperolides A–C, tetranorlabdane diterpenoids from the marine alga-derived endophytic fungus Aspergillus wentii EN-48. J. Nat. Prod. 2012, 75, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Miao, L.; Lv, C.; Sun, H.; Wei, S.; Wang, B.; Huang, C.; Jiao, B. Wentilactone B induces G2/M phase arrest and apoptosis via the Ras/Raf/MAPK signaling pathway in human hepatoma SMMC-7721 cells. Cell Death Dis. 2013, 4, e657. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.-P.; Liang, X.-R.; Liu, X.-H.; Ji, N.-Y. Aspewentins A–C, norditerpenes from a cryptic pathway in an algicolous strain of Aspergillus wentii. J. Nat. Prod. 2014, 77, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, B.S.; Prasad, R. Aspergillus dimorphicus and Emericella cleisto-minuta spp. nov. from Indian soils. Trans. Br. Mycol. Soc. 1969, 52, 331–336. [Google Scholar] [CrossRef]
- Box, G.E.P.; Hunter, W.G.; Hunter, J.S.B. Statistics for Experimenters: An Introduction to Design, Data Analysis, and Model Building; Wiley: New York, NY, USA, 1978. [Google Scholar]
- Zhu, Y.X.; Yao, L.Y.; Jiao, R.H.; Lu, Y.H.; Tan, R.X. Enhanced production of Fumigaclavine C in liquid culture of Aspergillus fumigatus under a two-stage process. Bioresour. Technol. 2014, 152, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.W. Phylogenetic analysis of Aspergillus sections Cremei and Wentii, based on ribosomal DNA-sequences. Mycol. Res. 1995, 99, 1349–1355. [Google Scholar] [CrossRef]
- Peterson, S.W. Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 2008, 100, 205–226. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.M.; Cole, R.J.; Kirksey, J.W. Emodin, a toxic metabolite of Aspergillus wentii isolated from weevil-damaged chestnuts. Appl. Microbiol. 1975, 30, 26–28. [Google Scholar] [PubMed]
- Daynes, C.N.; Zhang, N.; Saleeba, J.A.; McGee, P.A. Soil aggregates formed in vitro by saprotrophic Trichocomaceae have transient water-stability. Soil Biol. Biochem. 2012, 48, 151–161. [Google Scholar] [CrossRef]
- Li, W.K.; Fitzloff, J.F. HPLC-PDA determination of bioactive diterpenoids from plant materials and commercial products of Andrographis paniculata. J. Liq. Chromatogr. Relat. Technol. 2004, 27, 2407–2420. [Google Scholar] [CrossRef]
- Pu, X.; Qu, X.; Chen, F.; Bao, J.; Zhang, G.; Luo, Y. Camptothecin-producing endophytic fungus Trichoderma atroviride LY357: Isolation, identification, and fermentation conditions optimization for camptothecin production. Appl. Microbiol. Biotechnol. 2013, 97, 9365–9375. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, J.; Liu, P.; Wang, W.; Zhu, W. Three new compounds from Aspergillus terreus PT06-2 grown in a high salt medium. Mar. Drugs 2011, 9, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.R.; Lehmann, L.; Nielsen, J. Systemic analysis of the response of Aspergillus niger to ambient pH. Genome Biol. 2009, 10, R47. [Google Scholar] [CrossRef] [PubMed]
- Pettit, R.K. Small-molecule elicitation of microbial secondary metabolites. Microb. Biotechnol. 2011, 4, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Peterson, S.W.; Frisvad, J.C.; Varga, J. New species in Aspergillus section Terrei. Stud. Mycol. 2011, 69, 39–55. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, R.; Xu, G.-M.; Li, X.-M.; Li, C.-S.; Wang, B.-G. Characterization of a Newly Isolated Marine Fungus Aspergillus dimorphicus for Optimized Production of the Anti-Tumor Agent Wentilactones. Mar. Drugs 2015, 13, 7040-7054. https://doi.org/10.3390/md13117040
Xu R, Xu G-M, Li X-M, Li C-S, Wang B-G. Characterization of a Newly Isolated Marine Fungus Aspergillus dimorphicus for Optimized Production of the Anti-Tumor Agent Wentilactones. Marine Drugs. 2015; 13(11):7040-7054. https://doi.org/10.3390/md13117040
Chicago/Turabian StyleXu, Rui, Gang-Ming Xu, Xiao-Ming Li, Chun-Shun Li, and Bin-Gui Wang. 2015. "Characterization of a Newly Isolated Marine Fungus Aspergillus dimorphicus for Optimized Production of the Anti-Tumor Agent Wentilactones" Marine Drugs 13, no. 11: 7040-7054. https://doi.org/10.3390/md13117040
APA StyleXu, R., Xu, G. -M., Li, X. -M., Li, C. -S., & Wang, B. -G. (2015). Characterization of a Newly Isolated Marine Fungus Aspergillus dimorphicus for Optimized Production of the Anti-Tumor Agent Wentilactones. Marine Drugs, 13(11), 7040-7054. https://doi.org/10.3390/md13117040