
Dihydroaustrasulfone Alcohol Inhibits PDGF-Induced Proliferation and Migration of Human Aortic Smooth Muscle Cells through Inhibition of the Cell Cycle

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
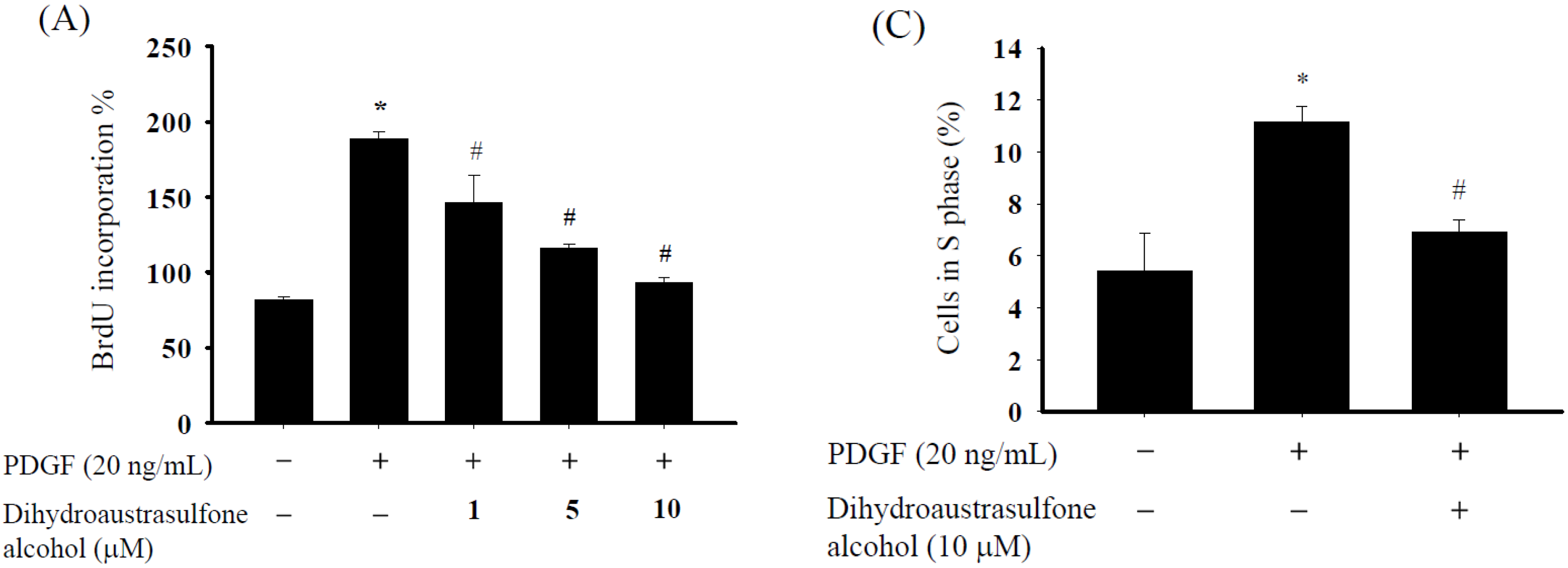
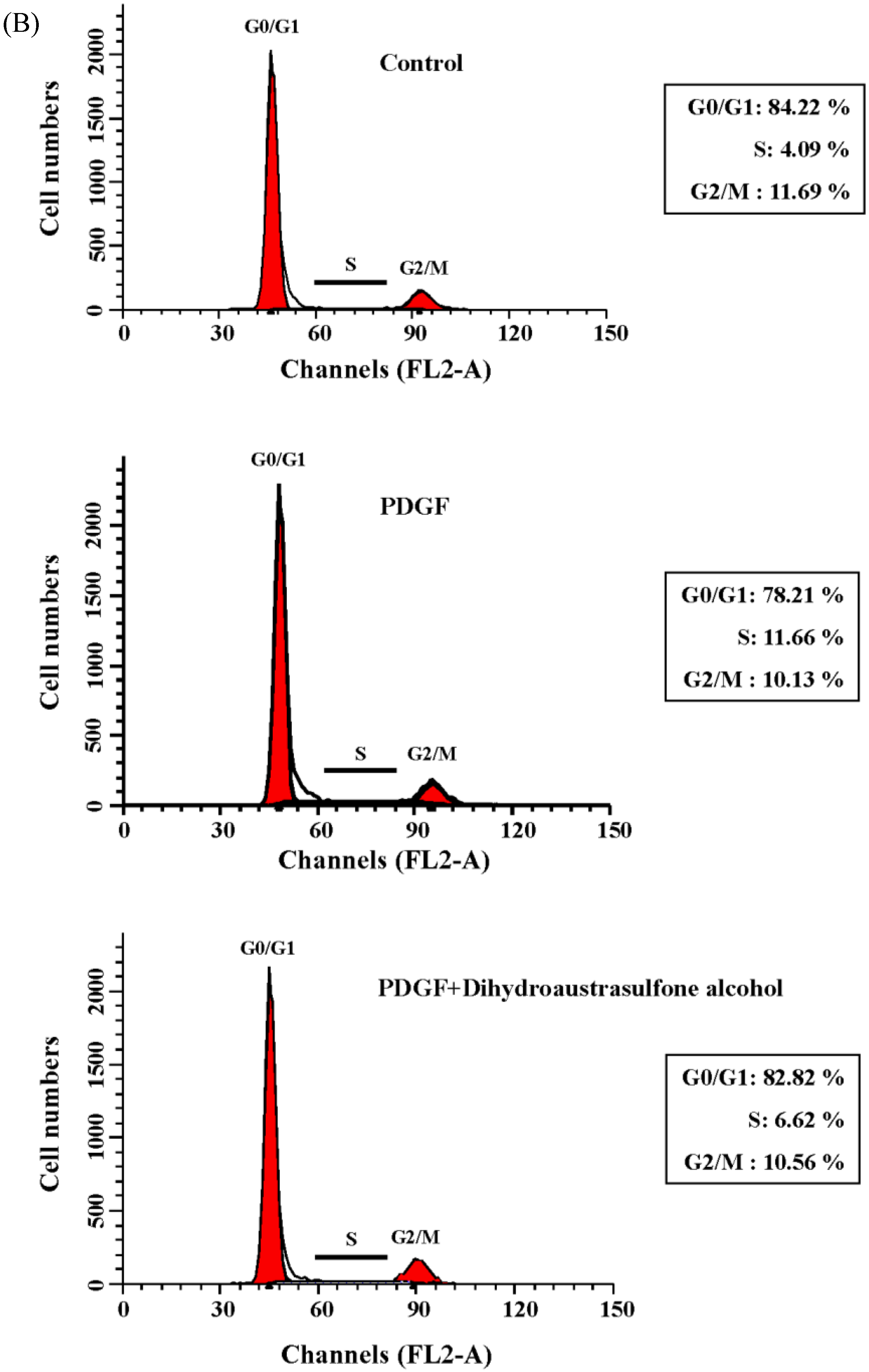
2.1. Dihydroaustrasulfone Alcohol Inhibits PDGF-Stimulated Proliferation in Human Aortic Smooth Muscle Cells


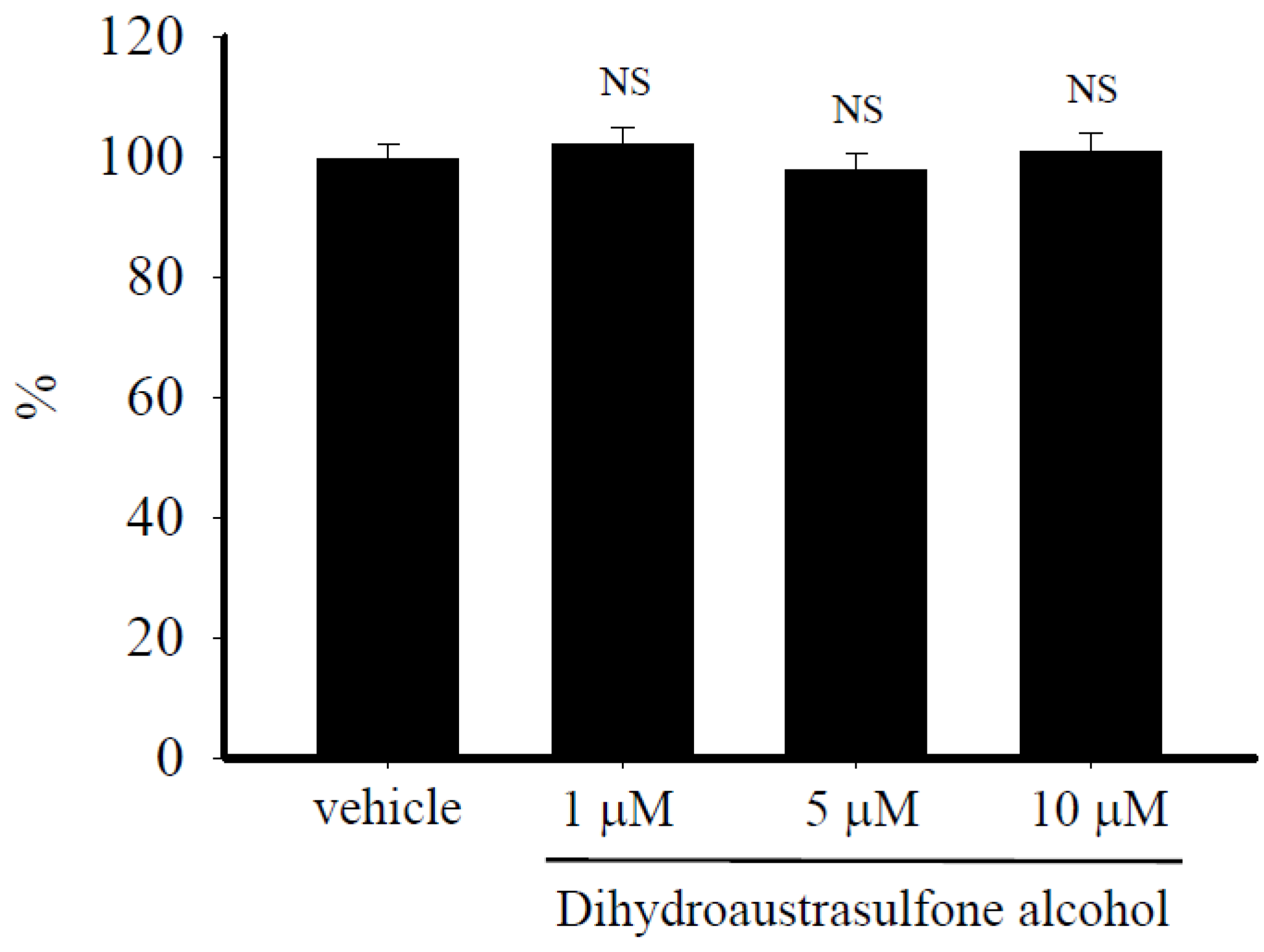
2.2. Dihydroaustrasulfone Alcohol Does not Affect HASMCs Viability

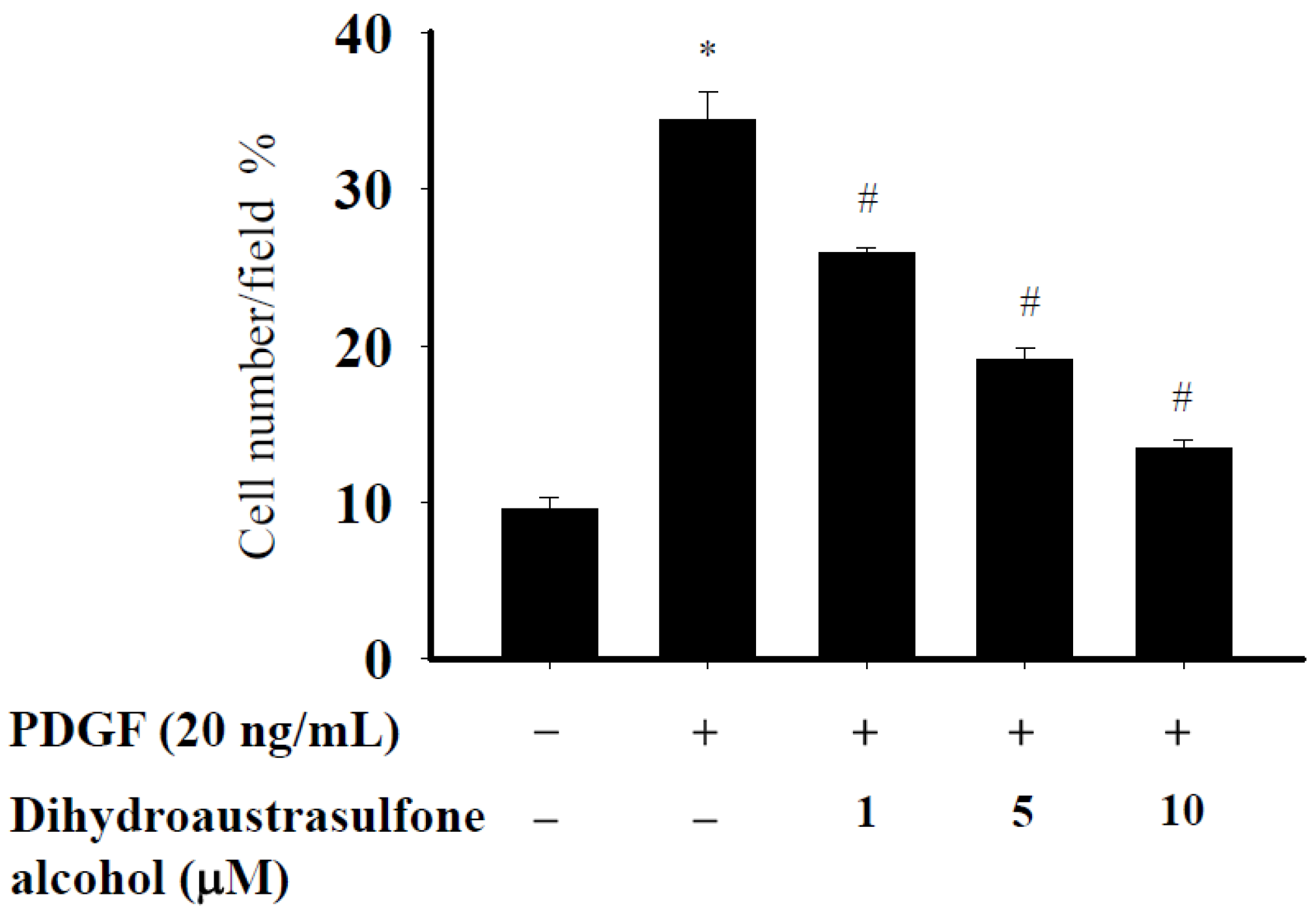
2.3. Dihydroaustrasulfone Alcohol Suppress the PDGF-Induced Migration of HASMCs
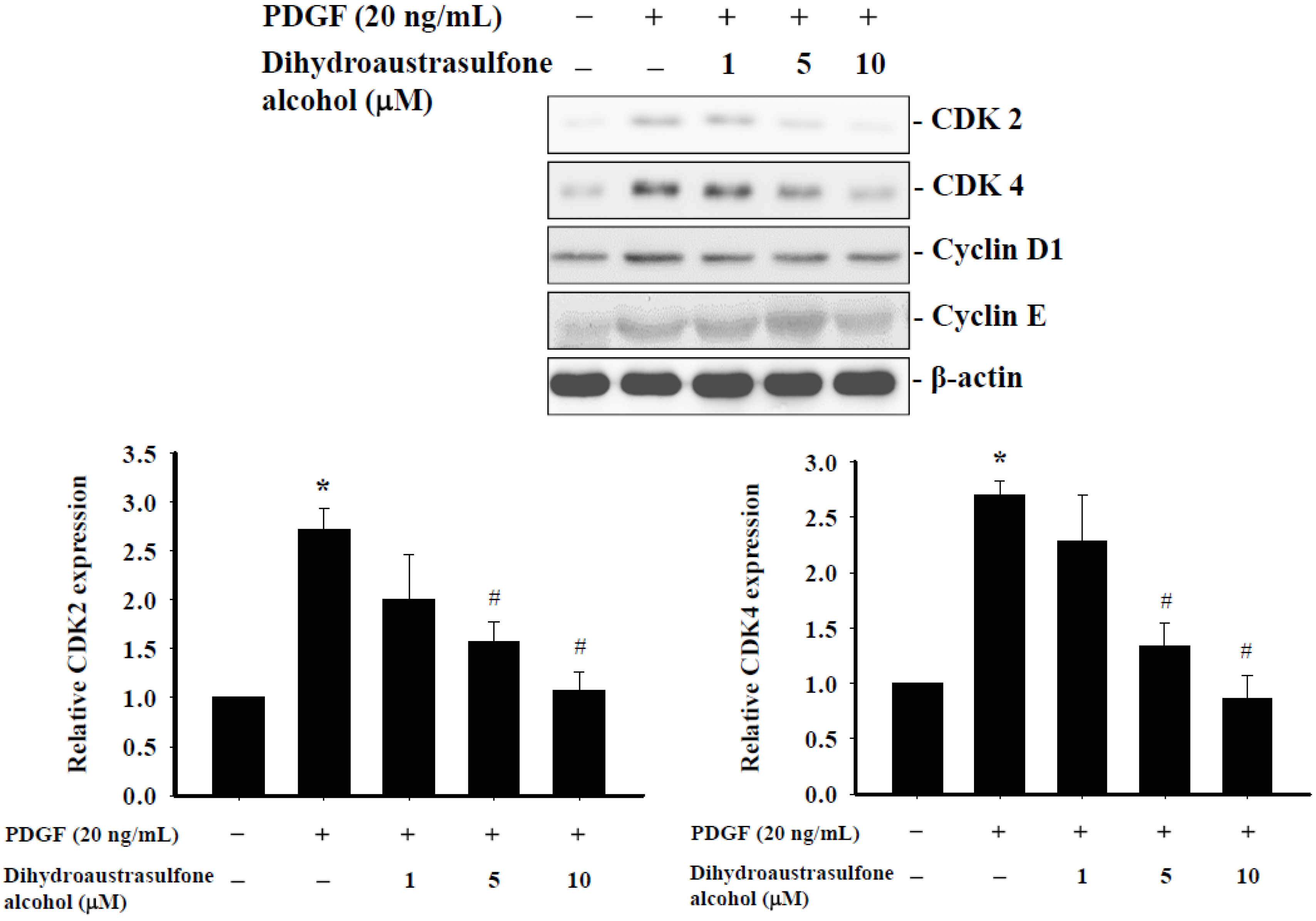
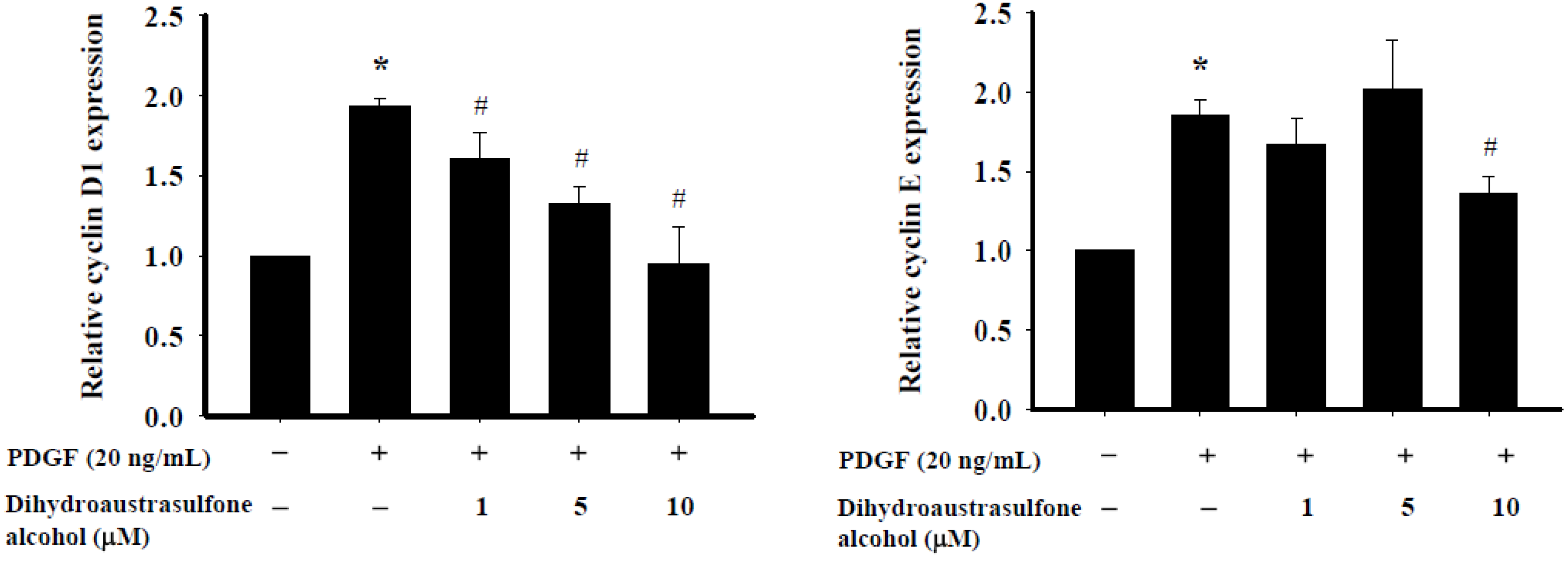
2.4. Dihydroaustrasulfone Alcohol Affects Expression of Cell Cycle Regulatory Proteins in HASMCs



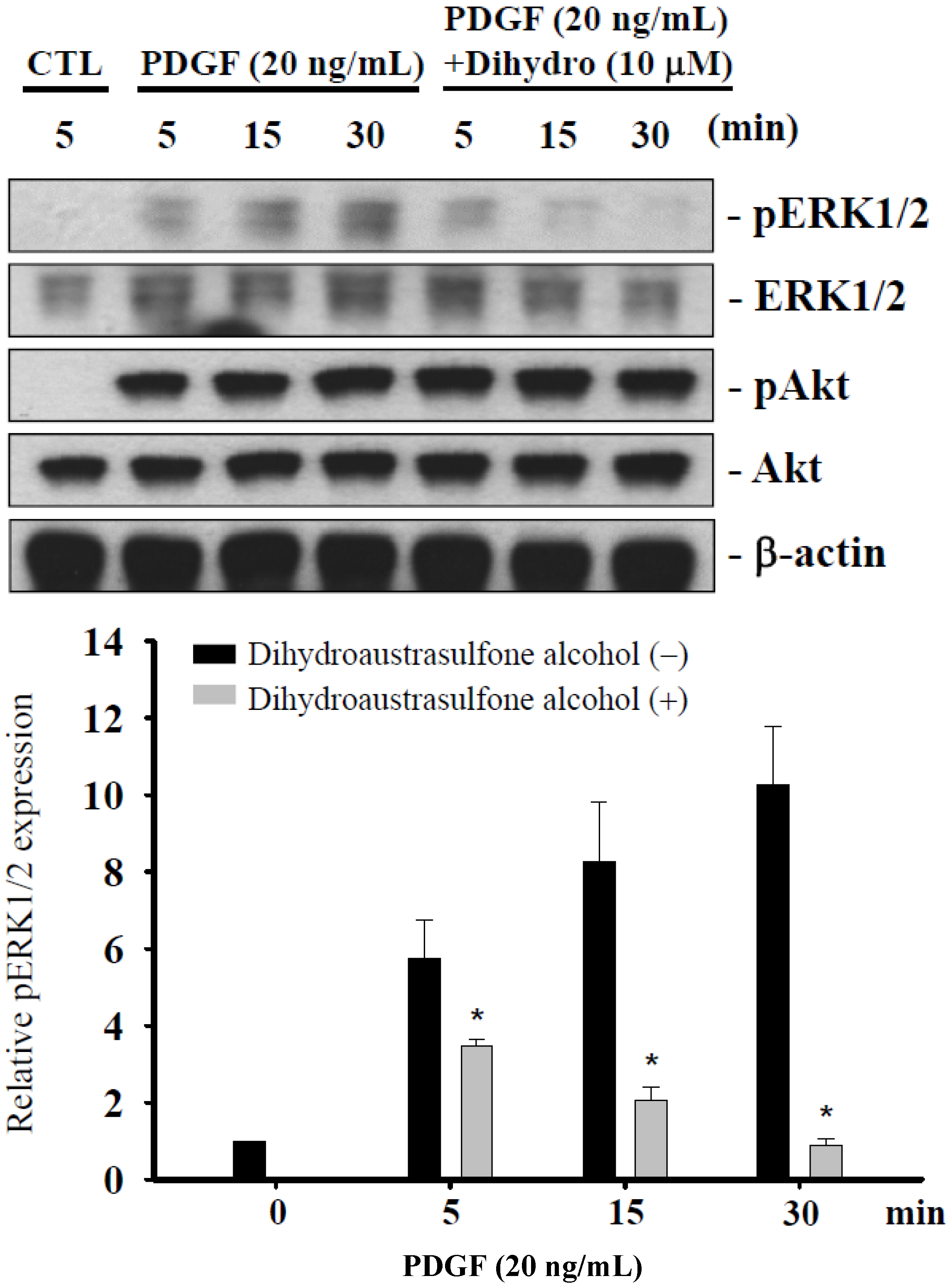
2.5. Dihydroaustrasulfone Alcohol Differentially Affects PDGF-Induced Phosphorylation of Extracellular Signal-Regulated Kinase 1/2 and Akt
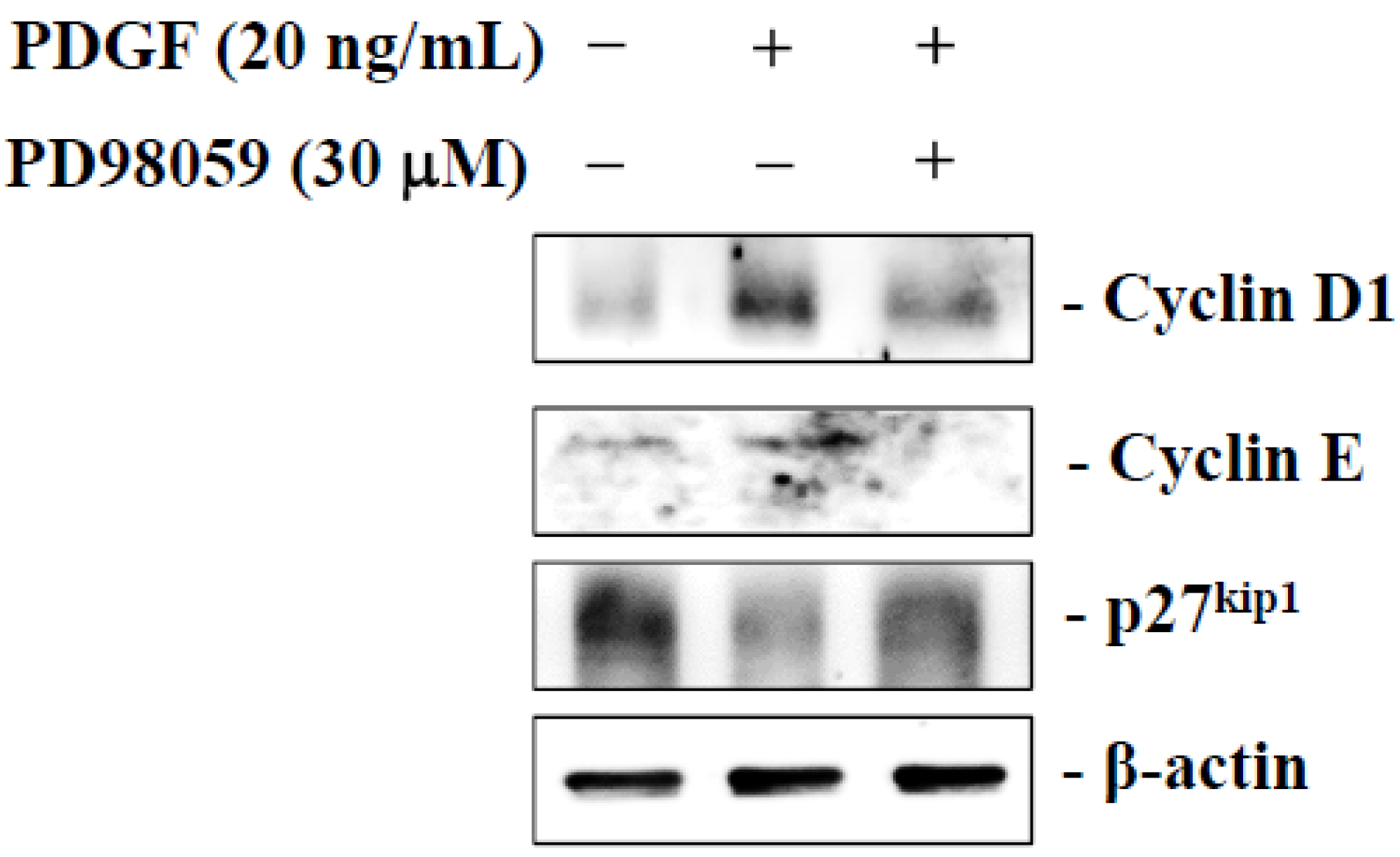
2.6. Dihydroaustrasulfone Alcohol Inhibits PDGF-Stimulated HASMCs Proliferation via the ERK Signaling Pathway

2.7. Dihydroaustrasulfone Alcohol Modulates HASMCs Phenotype


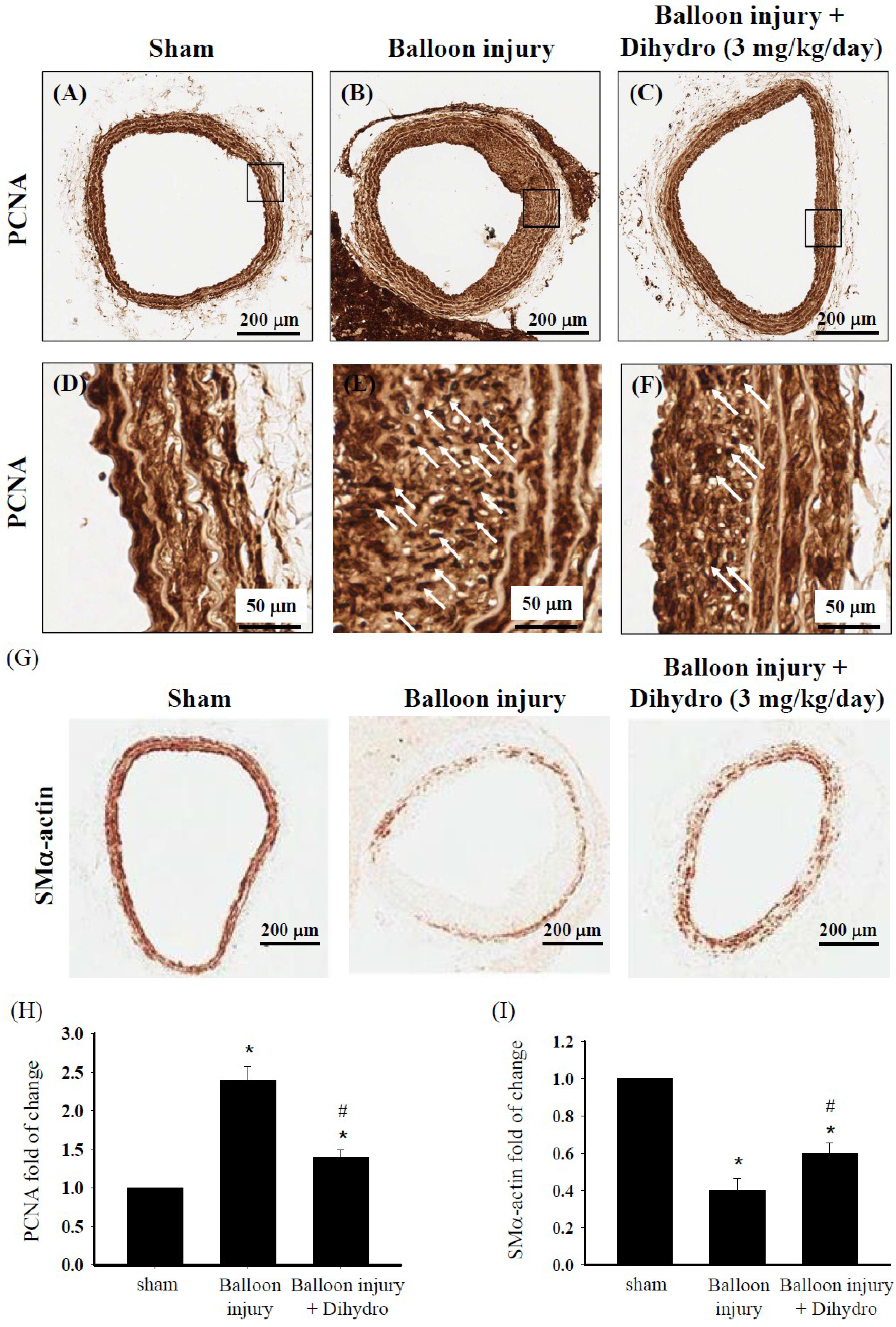
2.8. Dihydroaustrasulfone Alcohol Reduces Neointimal Thickening by Modulating VSMC Phenotype in a Rat Model of Restenosis Induced by Carotid Artery Balloon Injury

3. Discussion
4. Experimental Section
4.1. Materials
4.2. Cell Culture and Drug Treatment
4.3. BrdU Incorporation Assay
4.4. Flow Cytometry Analysis
4.5. Cell Viability Assay
4.6. Cell Migration Assay
4.7. Western Blot Analysis
4.8. Rat Balloon Injury Model
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ross, R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993, 362, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Owens, G.K.; Kumar, M.S.; Wamhoff, B.R. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol. Rev. 2004, 84, 767–801. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.M. Smooth muscle migration in atherosclerosis and restenosis. J. Clin. Invest. 1997, 100, S87–S89. [Google Scholar] [PubMed]
- Majesky, M.W.; Reidy, M.A.; Bowen-Pope, D.F.; Hart, C.E.; Wilcox, J.N.; Schwartz, S.M. PDGF ligand and receptor gene expression during repair of arterial injury. J. Cell Biol. 1990, 111, 2149–2158. [Google Scholar] [CrossRef] [PubMed]
- Miano, J.M.; Vlasic, N.; Tota, R.R.; Stemerman, M.B. Smooth muscle cell immediate-early gene and growth factor activation follows vascular injury. A putative in vivo mechanism for autocrine growth. Arterioscler. Thromb. 1993, 13, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Bowen-Pope, D.F.; Raines, E.W. Platelet-derived growth factor and its role in health and disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 327, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Jawien, A.; Bowen-Pope, D.F.; Lindner, V.; Schwartz, S.M.; Clowes, A.W. Platelet-derived growth factor promotes smooth muscle migration and intimal thickening in a rat model of balloon angioplasty. J. Clin. Investig. 1992, 89, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Warner, S.J.; Salomon, R.N.; Birinyi, L.K. Production of platelet-derived growth factor-like mitogen by smooth muscle cells from human atheroma. N. Engl. J. Med. 1988, 318, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Jean, Y.H.; Chen, W.F.; Duh, C.Y.; Huang, S.Y.; Hsu, C.H.; Lin, C.S.; Sung, C.S.; Chen, I.M.; Wen, Z.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory and analgesic effects of the natural marine compound lemnalol from Formosan soft coral Lemnalia cervicorni. Eur. J. Pharmacol. 2008, 578, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Jean, Y.H.; Chen, W.F.; Sung, C.S.; Duh, C.Y.; Huang, S.Y.; Lin, C.S.; Tai, M.H.; Tzeng, S.F.; Wen, Z.H. Capnellene, a natural marine compound derived from soft coral, attenuates chronic constriction injury-induced neuropathic pain in rats. Br. J. Pharmacol. 2009, 158, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.H.; Chao, C.H.; Wu, M.H.; Sheu, J.H. A neuroprotective sulfone of marine origin and the in vivo anti-inflammatory activity of an analogue. Eur. J. Med. Chem. 2010, 45, 5998–6004. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Chien, Y.C.; Pan, C.H.; Sheu, J.H.; Chen, C.Y.; Wu, C.H. Inhibitory effect of dihydroaustrasulfone alcohol on the migration of human non-small cell lung carcinoma A549 cells and the antitumor effect on a Lewis lung carcinoma-bearing tumor model in C57BL/6J mice. Mar. Drugs 2014, 12, 196–213. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, R.S.; Murphy, J.G.; Edwards, W.D.; Camrud, A.R.; Vliestra, R.E.; Holmes, D.R. Restenosis after balloon angioplasty. A practical proliferative model in porcine coronary arteries. Circulation 1990, 82, 2190–2200. [Google Scholar] [CrossRef] [PubMed]
- Ferns, G.A.; Avades, T.Y. The mechanisms of coronary restenosis: Insights from experimental models. Int. J. Exp. Pathol. 2000, 81, 63–88. [Google Scholar] [CrossRef] [PubMed]
- Leimgruber, P.P.; Roubin, G.S.; Hollman, J.; Cotsonis, G.A.; Meier, B.; Douglas, J.S.; King, S.B., Jr.; Gruentzig, A.R. Restenosis after successful coronary angioplasty in patients with single-vessel disease. Circulation 1986, 73, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Muslin, A.J. MAPK signalling in cardiovascular health and disease: Molecular mechanisms and therapeutic targets. Clin. Sci. (Lond.) 2008, 115, 203–218. [Google Scholar] [CrossRef]
- Zhan, Y.; Kim, S.; Izumi, Y.; Izumiya, Y.; Nakao, T.; Miyazaki, H.; Iwao, H. Role of JNK, p38, and ERK in platelet-derived growth factor-induced vascular proliferation, migration, and gene expression. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef] [PubMed]
- Force, T.; Bonventre, J.V. Growth factors and mitogen-activated protein kinases. Hypertension 1998, 31, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. Atherosclerosis-an inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Ross, R. Mechanisms of atherosclerosis—A review. Adv. Nephrol. Necker Hosp. 1990, 19, 79–86. [Google Scholar] [PubMed]
- Crews, C.M.; Alessandrini, A.; Erikson, R.L. The primary structure of MEK, a protein kinase that phosphorylates the ERK gene product. Science 1992, 258, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, B.A. Akt signaling: Linking membrane events to life and death decisions. Science 1997, 275, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Franke, T.F.; Kaplan, D.R.; Cantley, L.C. PI3K: Downstream AKTion blocks apoptosis. Cell 1997, 88, 435–437. [Google Scholar] [CrossRef] [PubMed]
- Kulik, G.; Klippel, A.; Weber, M.J. Antiapoptotic signalling by the insulin-like growth factor I receptor, phosphatidylinositol 3-kinase, and Akt. Mol. Cell. Biol. 1997, 17, 1595–1606. [Google Scholar] [PubMed]
- Lim, H.J.; Lee, S.; Park, J.H.; Lee, K.S.; Choi, H.E.; Chung, K.S.; Lee, H.H.; Park, H.Y. PPAR delta agonist L-165041 inhibits rat vascular smooth muscle cell proliferation and migration via inhibition of cell cycle. Atherosclerosis 2009, 202, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Rainger, G.E.; Nash, G.B. Cellular pathology of atherosclerosis: Smooth muscle cells prime co-cultured endothelial cells for enhanced leukocyte adhesion. Circ. Res. 2001, 88, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Landry, D.B.; Couper, L.L.; Bryant, S.R.; Lindner, V. Activation of the NF-kappa B and I kappa B system in smooth muscle cells after rat arterial injury. Induction of VCAM-1 and MCP-1. Am. J. Pathol. 1997, 151, 1085–1095. [Google Scholar] [PubMed]
- Porreca, E.; Di Febbo, C.; Reale, M.; Castellani, M.L.; Baccante, G.; Barbacane, R.; Conti, P.; Cuccurullo, F.; Poggi, A. Monocyte chemotactic protein 1 (MCP-1) is a mitogen for cultured rat vascular smooth muscle cells. J. Vasc. Res. 1997, 34, 58–65. [Google Scholar] [PubMed]
- Yue, T.L.; Wang, X.; Sung, C.P.; Olson, B.; McKenna, P.J.; Gu, J.L.; Feuerstein, G.Z. Interleukin-8—A mitogen and chemoattractant for vascular smooth muscle cells. Circ. Res. 1994, 75, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dzau, V.J.; Braun-Dullaeus, R.C.; Sedding, D.G. Vascular proliferation and atherosclerosis: New perspectives and therapeutic strategies. Nat. Med. 2002, 8, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Rensen, S.S.; Doevendans, P.A.; van Eys, G.J. Regulation and characteristics of vascular smooth muscle cell phenotypic diversity. Neth. Heart J. 2007, 15, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yang, X.; Friesel, R.E.; Vary, C.P.; Liaw, L. Mechanisms of TGF-β-induced differentiation in human vascular smooth muscle cells. J. Vasc. Res. 2011, 48, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, G.; Betts, C.; Harmon, E.Y.; Keller, R.S.; Van De Water, L.; Zhou, J. Transforming growth factor-β1-induced transcript 1 protein, a novel marker for smooth muscle contractile phenotype, is regulated by serum response factor/myocardin protein. J. Biol. Chem. 2011, 286, 41589–41599. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.W.; Huang, S.C.; Kuo, H.M.; Chen, C.H.; Ma, Y.L.; Chu, T.H.; Bee, Y.S.; Wang, E.M.; Wu, C.Y.; Sung, P.J.; et al. Coral-derived compound WA-25 inhibits angiogenesis by attenuating the VEGF/VEGFR2 signaling pathway. Mar. Drugs 2015, 13, 861–878. [Google Scholar] [CrossRef] [PubMed]
- Redmond, E.M.; Cahill, P.A.; Hirsch, M.; Wang, Y.N.; Sitzmann, J.V.; Okada, S.S. Effect of pulse pressure on vascular smooth muscle cell migration: The role of urokinase and matrix metalloproteinase. Thromb. Haemost. 1999, 81, 293–300. [Google Scholar] [PubMed]
- Berger, M.; Rubinraut, E.; Barshack, I.; Roth, A.; Keren, G.; George, J. Zinc reduces intimal hyperplasia in the rat carotid injury model. Atherosclerosis 2004, 175, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Wu, C.C.; Hsiao, G.; Yen, M.H. Magnolol induces apoptosis in vascular smooth muscle. Naunyn Schmiedebergs Arch. Pharmacol. 2003, 368, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Chen, N.F.; Chen, W.F.; Hung, H.C.; Lee, H.P.; Lin, Y.Y.; Wang, H.M.; Sung, P.J.; Sheu, J.H.; Wen, Z.H. Sinularin from indigenous soft coral attenuates nociceptive responses and spinal neuroinflammation in carrageenan-induced inflammatory rat model. Mar. Drugs 2012, 10, 1899–1919. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Wen, Z.-H.; Lee, Y.-H.; Chen, C.-L.; Hung, H.-C.; Chen, C.-H.; Chen, W.-F.; Tsai, M.-C. Dihydroaustrasulfone Alcohol Inhibits PDGF-Induced Proliferation and Migration of Human Aortic Smooth Muscle Cells through Inhibition of the Cell Cycle. Mar. Drugs 2015, 13, 2390-2406. https://doi.org/10.3390/md13042390
Chen Y-C, Wen Z-H, Lee Y-H, Chen C-L, Hung H-C, Chen C-H, Chen W-F, Tsai M-C. Dihydroaustrasulfone Alcohol Inhibits PDGF-Induced Proliferation and Migration of Human Aortic Smooth Muscle Cells through Inhibition of the Cell Cycle. Marine Drugs. 2015; 13(4):2390-2406. https://doi.org/10.3390/md13042390
Chicago/Turabian StyleChen, Yao-Chang, Zhi-Hong Wen, Yen-Hsien Lee, Chu-Lun Chen, Han-Chun Hung, Chun-Hong Chen, Wu-Fu Chen, and Min-Chien Tsai. 2015. "Dihydroaustrasulfone Alcohol Inhibits PDGF-Induced Proliferation and Migration of Human Aortic Smooth Muscle Cells through Inhibition of the Cell Cycle" Marine Drugs 13, no. 4: 2390-2406. https://doi.org/10.3390/md13042390
APA StyleChen, Y. -C., Wen, Z. -H., Lee, Y. -H., Chen, C. -L., Hung, H. -C., Chen, C. -H., Chen, W. -F., & Tsai, M. -C. (2015). Dihydroaustrasulfone Alcohol Inhibits PDGF-Induced Proliferation and Migration of Human Aortic Smooth Muscle Cells through Inhibition of the Cell Cycle. Marine Drugs, 13(4), 2390-2406. https://doi.org/10.3390/md13042390