
Nrf2 and NF-κB Signaling Pathways Contribute to Porphyra-334-Mediated Inhibition of UVA-Induced Inflammation in Skin Fibroblasts

Abstract
:1. Introduction
2. Results
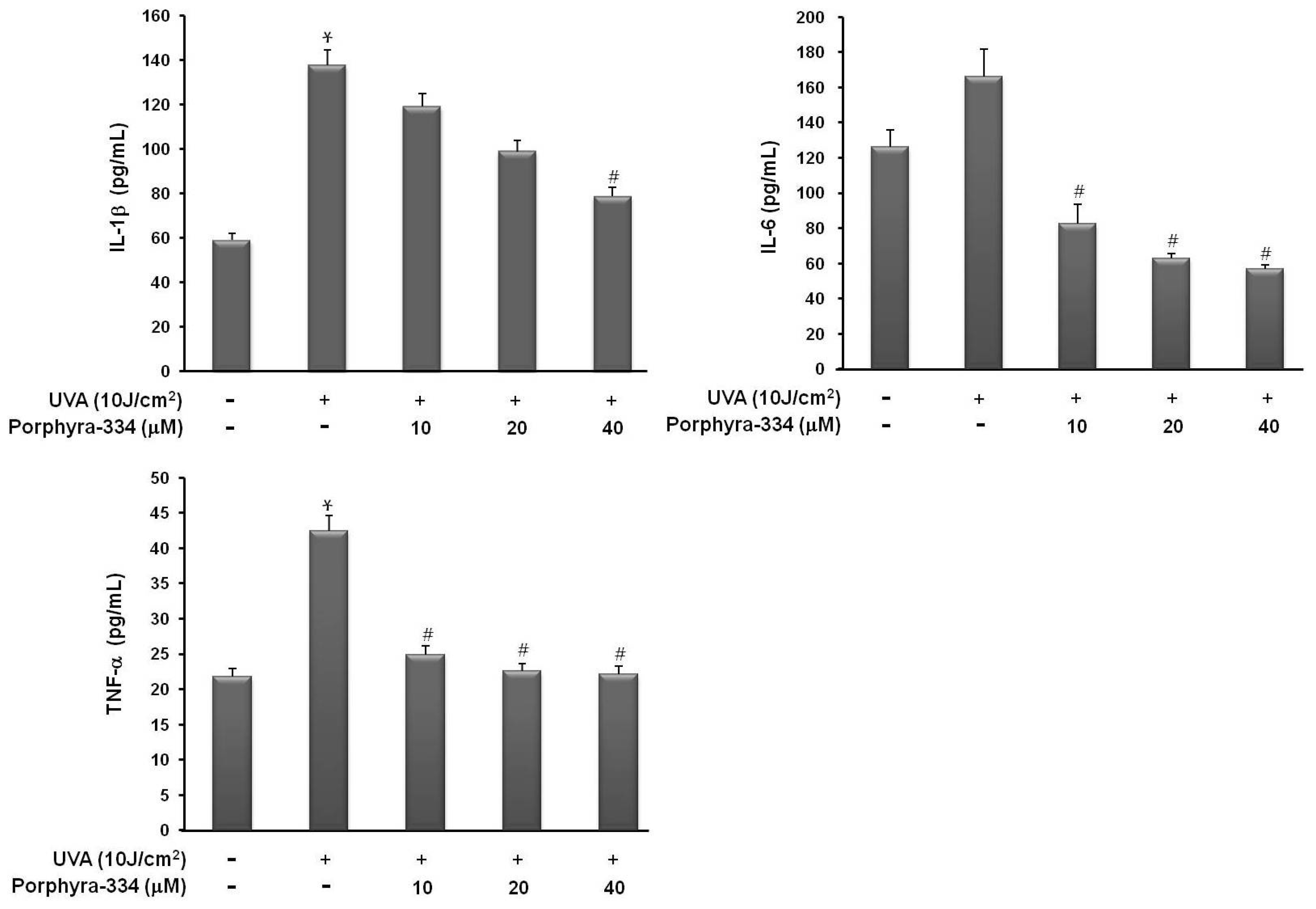
2.1. Effect of Porphyra-334 on UVA-Induced Proinflammatory Cytokine Production

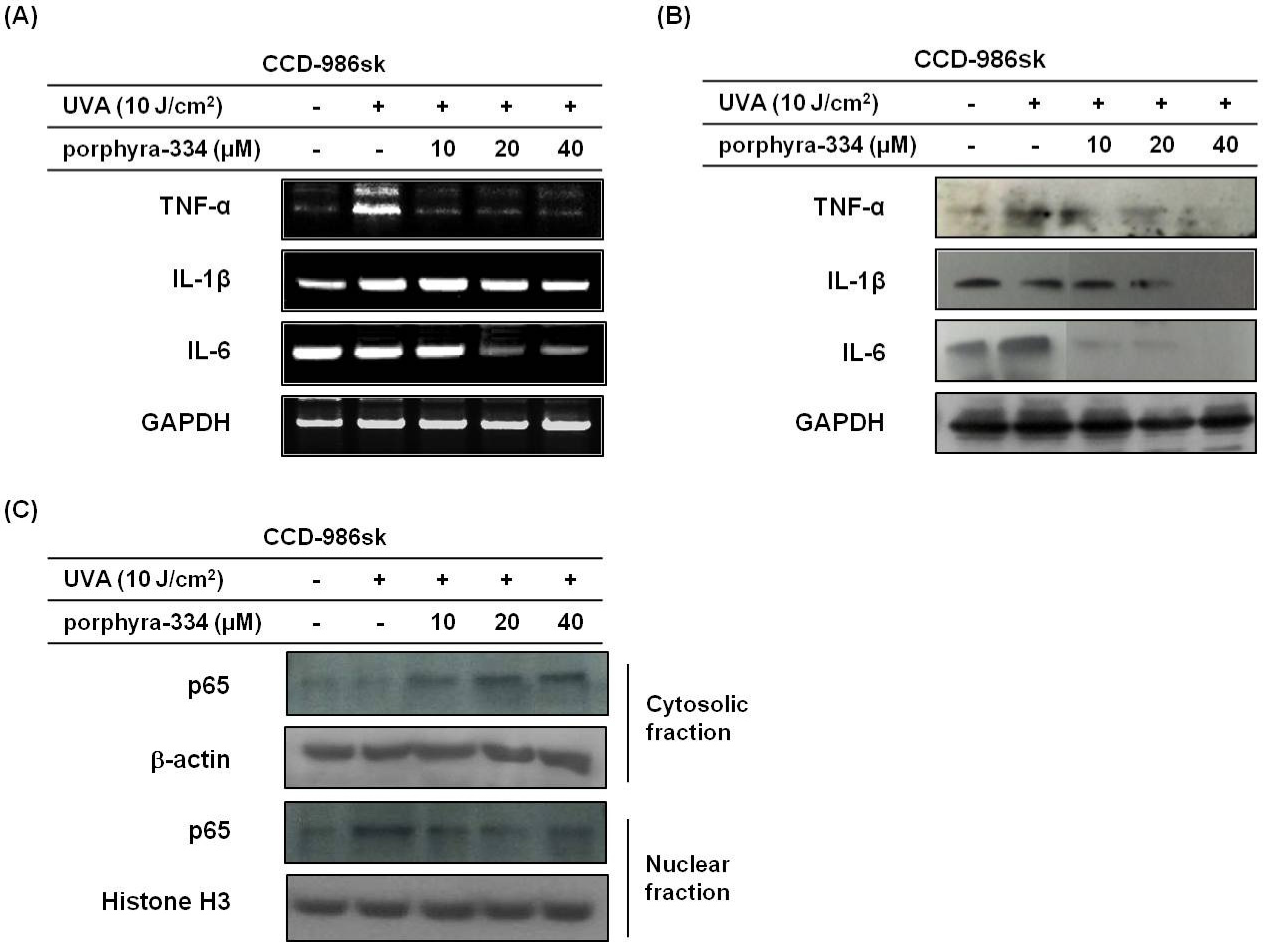
2.2. Effect of Porphyra-334 on UVA-Induced Inflammatory Response

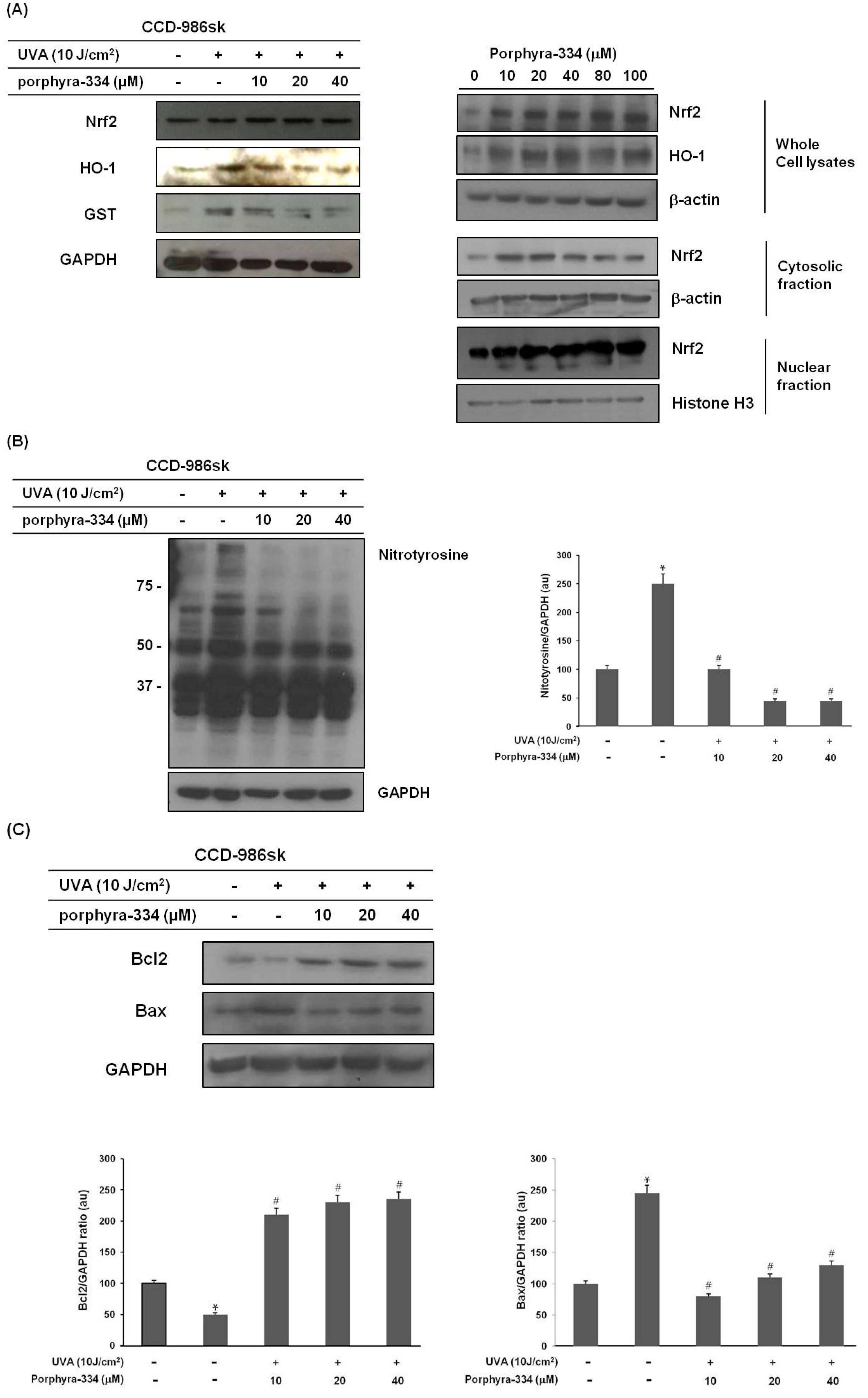
2.3. Porphyra-334 Induces the Nuclear Translocation of Nrf2 and Triggers Nrf2-Dependent Induction of HO-1 Expression Levels in Human Skin Fibroblasts

2.4. Porphyra-334 Inhibits UVA-Induced Oxidative Stress and Prevents Cell Death
3. Discussion

4. Experimental Section
4.1. Extraction and Isolation of Water-Soluble Porphyra-334
4.2. Cell Culture
4.3. UVA Irradiation and Porphyra-334 Treatment
4.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′–3′) | |
|---|---|---|
| Forward Primer | Reverse Primer | |
| TNF-α | TGCACCACAGTTTAAACCCA | GACTCCTTCAGGTGCTCAGG |
| IL-6 | AGGAGACTTGCCTGGTGAAA | CAGGGGTGGTTATTGCATCT |
| IL-1β | CTGTCCTGCGTGTTGAAAGA | TTCTGCTTGAGAGGTGCTGA |
4.5. Subcellular Fractionation
4.6. Western Blot Analysis
| Primary Antibody | COMPANY | Dilution Rate |
|---|---|---|
| μ-Actin | Santa Cruz Biotechnology, Inc. | 1:1000 |
| Bax | Cell Signaling TECHNOLOGY | 1:1000 |
| Bcl2 | Cell Signaling TECHNOLOGY | 1:1000 |
| GAPDH | Santa Cruz Biotechnology, Inc | 1:1000 |
| HO-1 | Santa Cruz Biotechnology, Inc | 1:1000 |
| IL-1β | Santa Cruz Biotechnology, Inc | 1:1000 |
| NF-κB | Cell Signaling TECHNOLOGY | 1:1000 |
| Nrf2 | Enzo Life Science | 1:1000 |
| Nitrotyrosine | Cell Signaling TECHNOLOGY | 1:1000 |
| TNF-α | Santa Cruz Biotechnology, Inc. | 1:1000 |
4.7. Measurement of TNF-α, IL-6 and IL-1β Production
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ichihashi, M.; Ueda, M.; Budiyanto, A.; Bito, T.; Oka, M.; Fukunage, M.; Tsuru, K.; Horikawa, T. UV-induced skin damage. Toxicology 2003, 189, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Datta, S.C.; Talwar, H.S.; Wang, Z.Q.; Varani, J.; Kang, S.; Voorhees, J.J. Molecular basis of sun-induced premature skin aging and retinoid antagonism. Nature 1996, 379, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Valacchi, G.; Sticozzi, C.; Pecorelli, A.; Cervellati, F.; Cervellati, C.; Maioli, E. Cutaneous responses to environmental stressors. Ann. N. Y. Acad. Sci. 2012, 1271, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Wondark, G.T.; Myron, K.; Jacobson, J.; Jacobson, E. Endogenous UVA-photo sensitizers: Mediators of skin photodamage and novel targets for skin photoprotection. Photochem. Photobiol. Sci. 2006, 5, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Soeur, J.; Eilstein, J.; Lereanx, G.; Jones, C.; Marrot, L. Skin resistance to oxidative stress induced by resveratrol from Nrf2 activation to GSH biosynthesis. Free Radic. Biol. Med. 2015, 78, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Hyberson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative stress in health and disease: The therapeutic potential of Nrf2 activation. Mol. Aspects Med. 2011, 32, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, M.B.; Anz, S.; Ben Sasson, S.; Bianco-Peled, H.; Kohen, R. Can nitroxides evoke the keap1-Nrf2-ARE pathway in skin? Free Radic. Biol. Med. 2014, 77, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.M.; Chen, H.C.; Lin, J.J.; Shih, I.C.; Wen, K.C. Michelia alba extract attenuates UVB-induced expression of matrix metalloproteinases via MAP kinase pathway in human dermal fibroblasts. Food Chem. Toxicol. 2012, 50, 4260–4269. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Hyun, Y.J.; Cho, S.J.; Kang, H.K.; Yoo, E.S.; Koh, Y.S.; Lee, N.H.; Ko, M.H.; Hyun, J.W. An ethanol extract derived from Bonnemaisonia hamifera scavenges ultraviolet B (UVB) radiation-induced reactive oxygen species and attenuates UVB-induced cell damage in human keratinocytes. Mar. Drugs 2012, 10, 2826–2845. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.; Farwanah, H.; Willrodt, A.H.; Huebner, A.J.; Sandhoff, K.; Roop, D.; Hohl, D.; Bloch, W.; Werner, S. Nrf2 links epidermal barriers function with antioxidant defense. EMBO Mol. 2012, 4, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Hirota, A.; Kawachi, Y.; Yamamoto, M.; Koga, T.; Hamada, K.; Otsnka, F. Acceleration of UVB-induced photoaging in Nrf2 gene-deficient mice. Exp. Dermatol. 2011, 20, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.V.; Walsh, N.A. Antioxidant and antiproliferative activities of extracts from a variety of edible seaweeds. Food Chem. Toxicol. 2006, 44, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Andreguetti, D.; Stein, E.M.; Pereira, C.M.; Pinto, E.; Colepicpolo, P. Antioxidant properties and UV absorbance pattern of mycosporine-like amino acids analogs synthesized in an environmentally friendly manner. J. Biochem. Mol. Toxicol. 2013, 27, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Park, S.J.; Kim, I.H.; Choi, Y.H.; Nam, T.J. Protective effect of porphyra 334 on UVA-induced photoaging in human skin fibroblasts. Int. J. Mol. Med. 2014, 34, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Pi, S.H.; Lee, S.I.; Kim, E.C. The anti-inflammatory role of heme oxygenase-1 in LPS and cytokine-stimulated inducible nitric oxide synthase and nitric oxide production in human periodontal ligament cells. J. Periodontal. 2009, 80, 2045–2055. [Google Scholar] [CrossRef] [PubMed]
- Jais, A.; Einwallner, E.; Sharif, O.; Gossens, K.; Lu, T.T.; Soyal, S.M.; Medgyesi, D.; Neureiter, D.; Paier-Pourani, J.; Dalgaard, K.; et al. Heme oxygenase-1 drives mataflammation and insulin resistance in mouse and man. Cell 2014, 3, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NF-κB inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.H.; Qu, J.; Shen, X. NF-κB/p65 antagonizes Nrf2-ARE pathway by deriving CBP from Nrf2 and facilitating recruitment of HDAC3 to Mafk. Biochim. Biophys. Acta 2008, 1783, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Greten, F.R. NF-κB: Linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 2005, 5, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, H.; Yamamoto, M. Nrf2-keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Sandireddy, R.; Yerra, V.G.; Areti, A.; Komirishetty, P.; Kumar, A. Neuroinflammation and oxidative stress in diabetic neuropathy : Futuristic strategies based on these targets. Int. J. Endocrionol. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J. NF-κB and Nrf2 as potential chemopreventive targets of some anti-inflammatory and anti-oxidative phytonutrients with anti-inflammatory and anti-oxidative activities. Asia Pac. J. Clin. Nutr. 2008, 17, 269–272. [Google Scholar] [PubMed]
- Zhao, Y.F.; Zhang, Q.; Xi, J. Y.; Li, Y.H.; Ma, C.G.; Xiao, B. G. Multitarget intervention of Fasudil in the neuroprotection of dopaminergic neurons in MPTP-mouse model of Parkinson’s disease. J. Neurol. Sci. 2015, in press. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, J.; Kwon, M.-J.; Nam, T.-J. Nrf2 and NF-κB Signaling Pathways Contribute to Porphyra-334-Mediated Inhibition of UVA-Induced Inflammation in Skin Fibroblasts. Mar. Drugs 2015, 13, 4721-4732. https://doi.org/10.3390/md13084721
Ryu J, Kwon M-J, Nam T-J. Nrf2 and NF-κB Signaling Pathways Contribute to Porphyra-334-Mediated Inhibition of UVA-Induced Inflammation in Skin Fibroblasts. Marine Drugs. 2015; 13(8):4721-4732. https://doi.org/10.3390/md13084721
Chicago/Turabian StyleRyu, Jina, Mi-Jin Kwon, and Taek-Jeong Nam. 2015. "Nrf2 and NF-κB Signaling Pathways Contribute to Porphyra-334-Mediated Inhibition of UVA-Induced Inflammation in Skin Fibroblasts" Marine Drugs 13, no. 8: 4721-4732. https://doi.org/10.3390/md13084721
APA StyleRyu, J., Kwon, M. -J., & Nam, T. -J. (2015). Nrf2 and NF-κB Signaling Pathways Contribute to Porphyra-334-Mediated Inhibition of UVA-Induced Inflammation in Skin Fibroblasts. Marine Drugs, 13(8), 4721-4732. https://doi.org/10.3390/md13084721