
Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus




Abstract
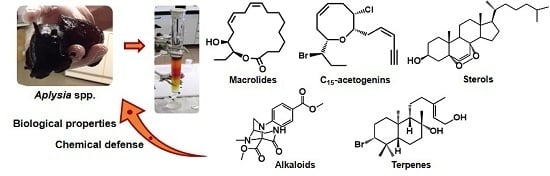
:
{kind=link}
{kind=link}
{kind=link}
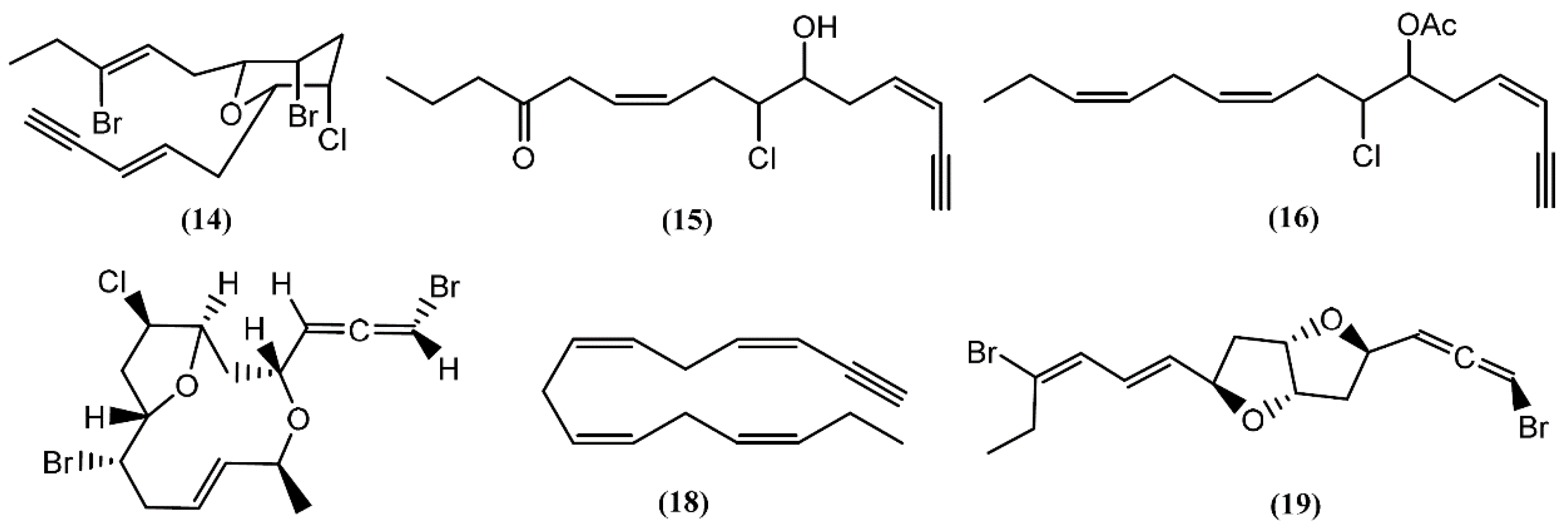{kind=link}
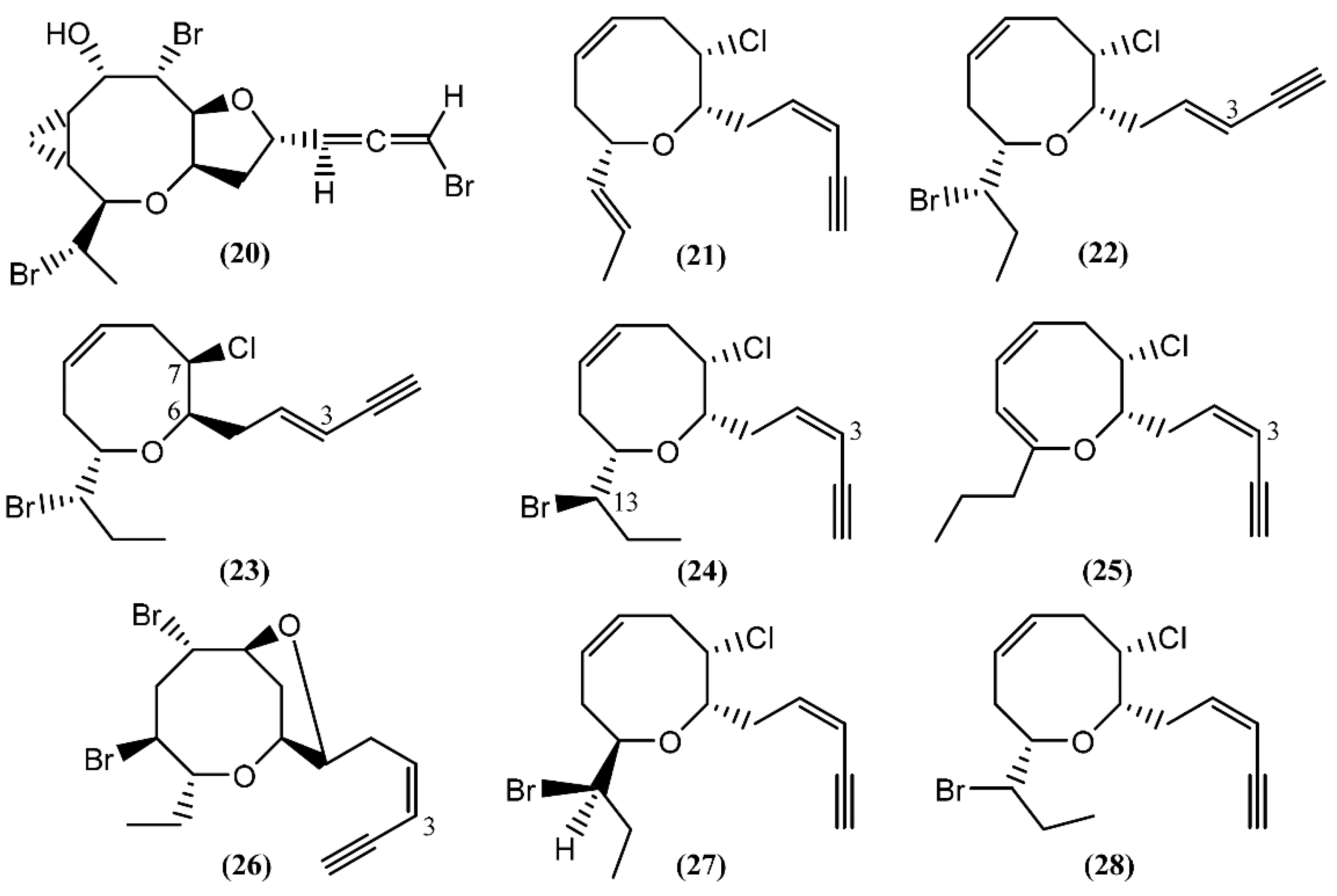{kind=link}
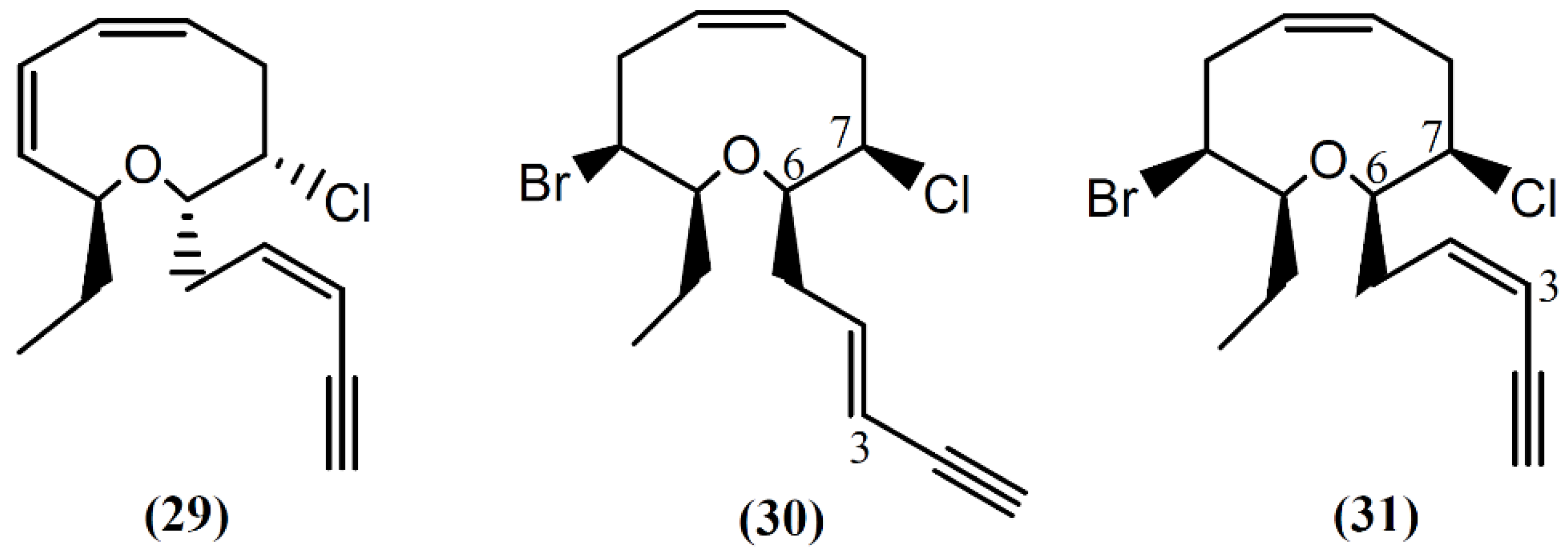{kind=link}
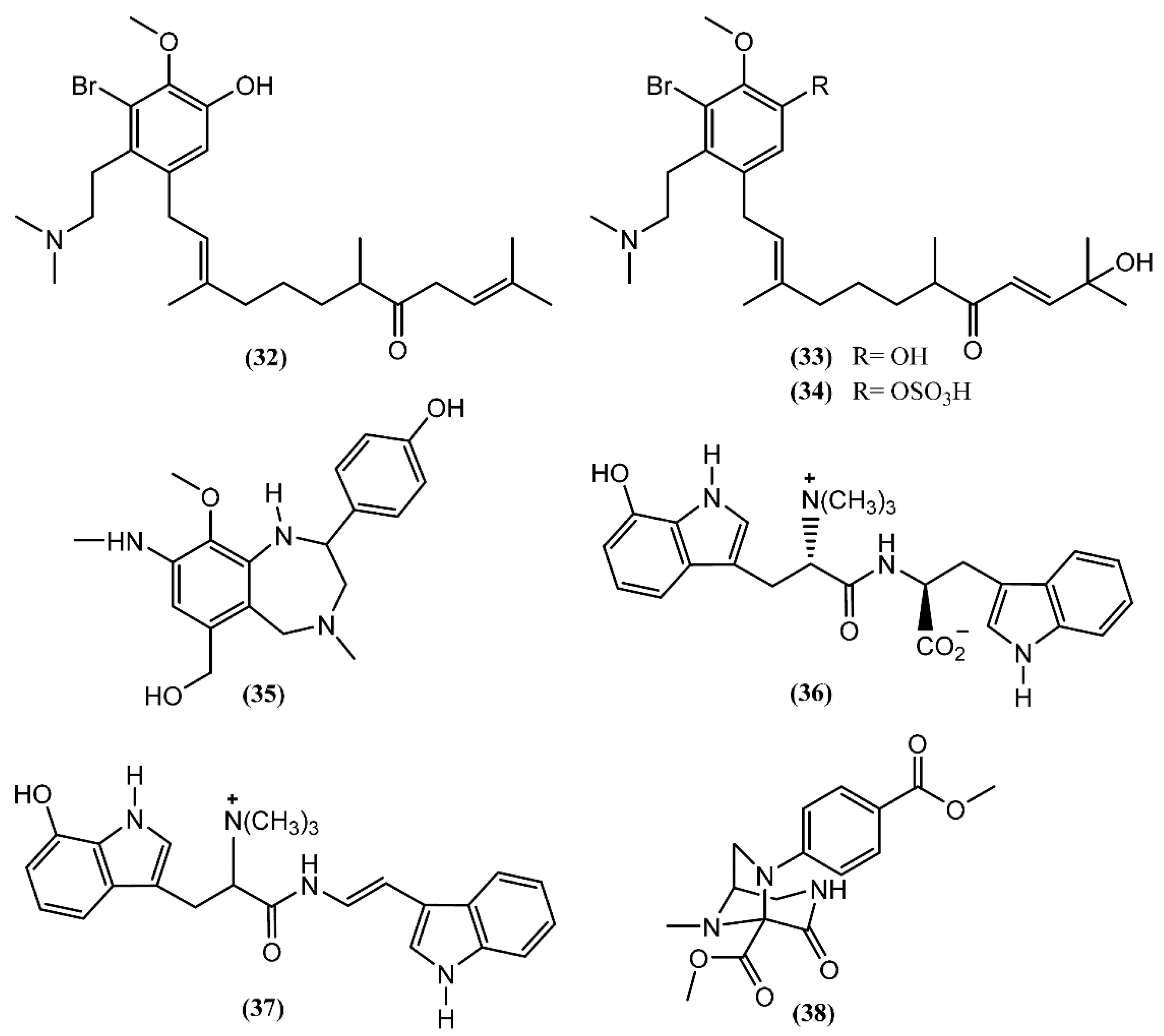{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
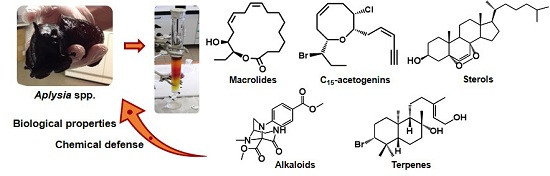
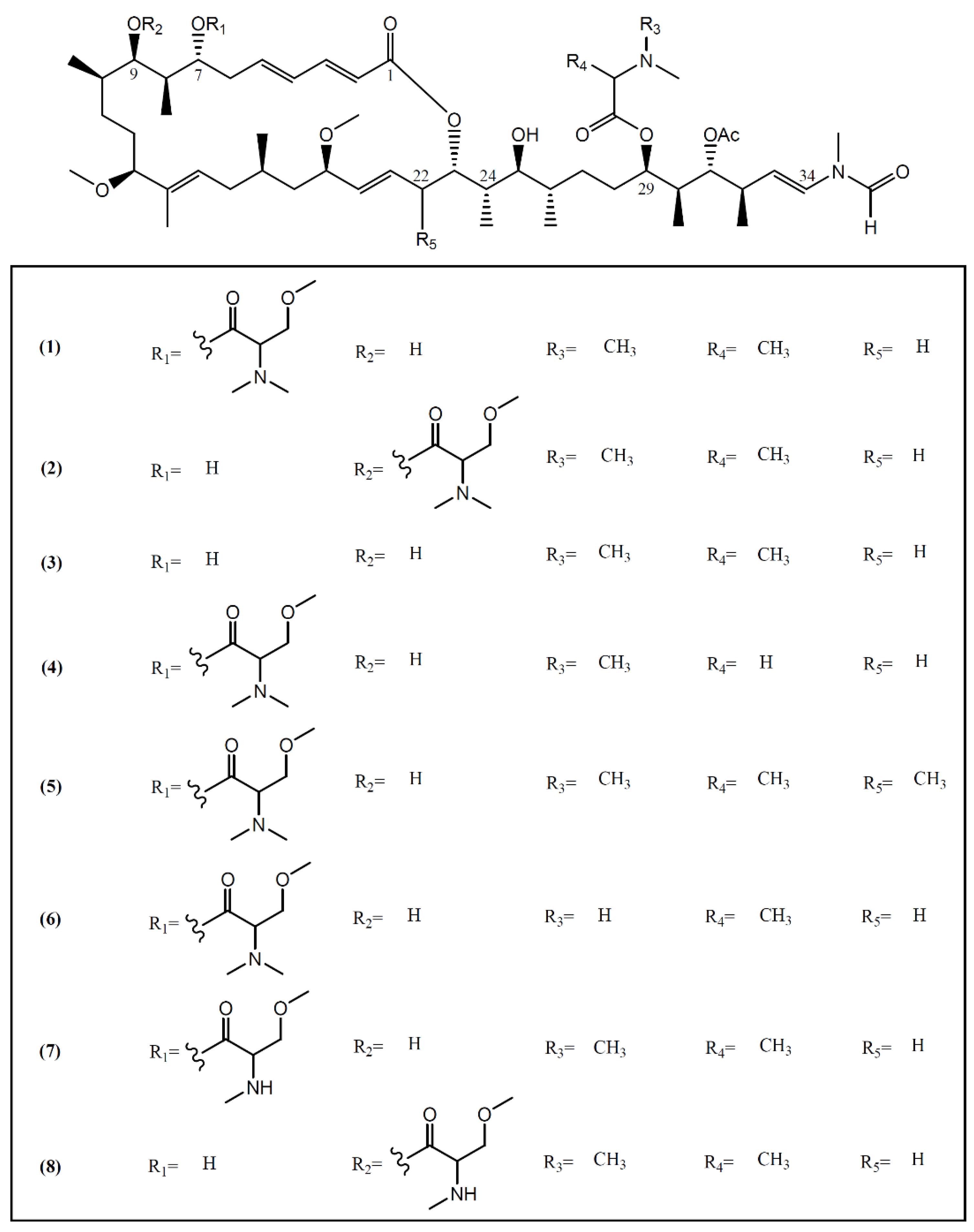
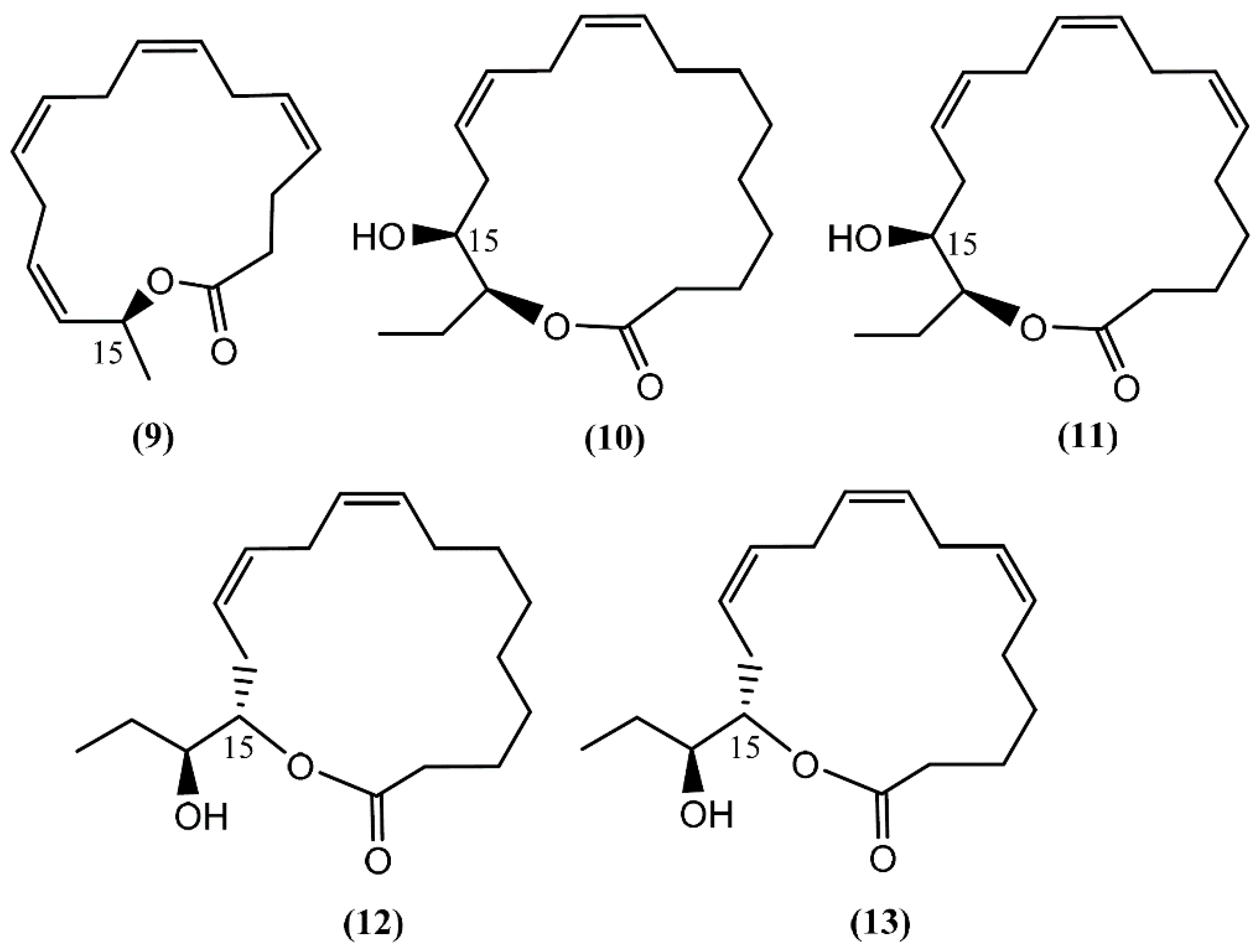
2. Macrolides
3. C15-Acetogenins
4. Alkaloids
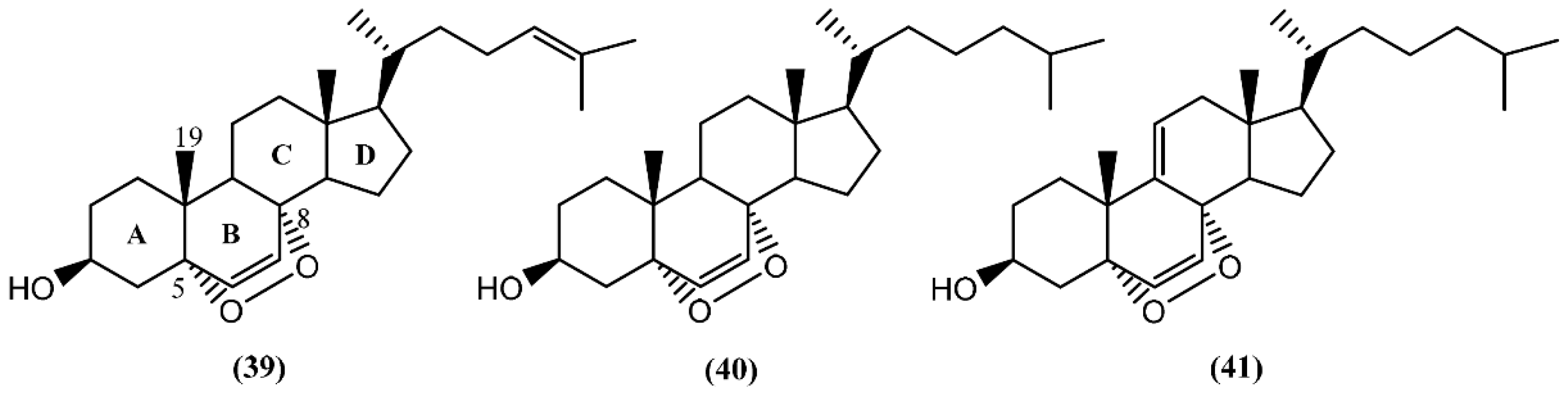
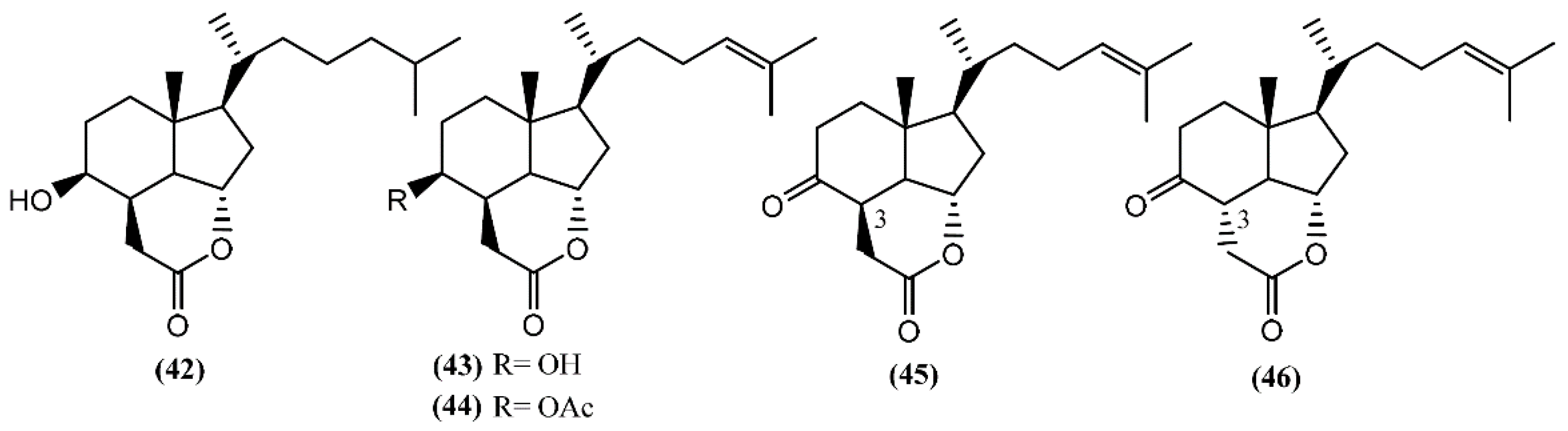
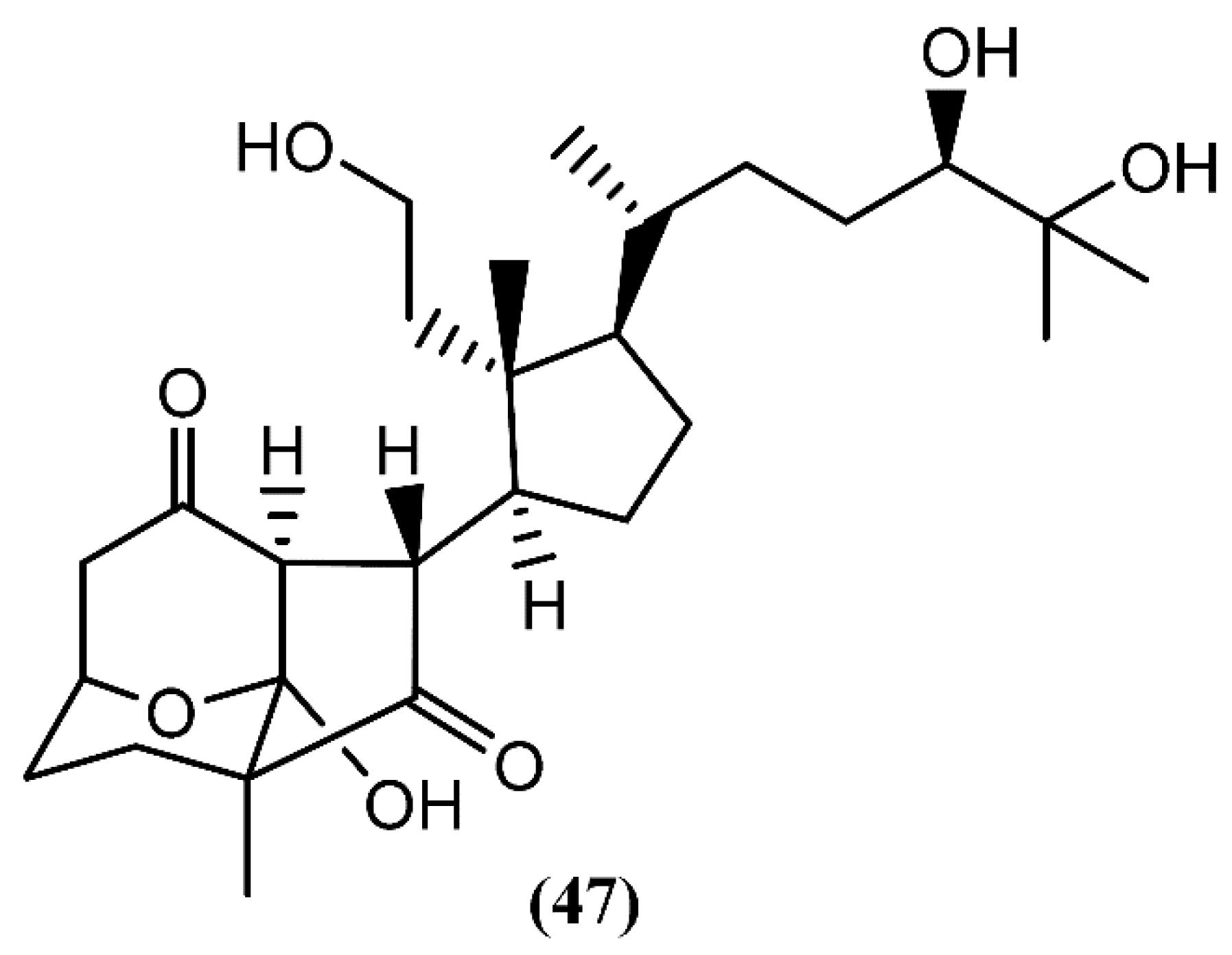
5. Sterols and Degraded Sterols
6. Terpenoids
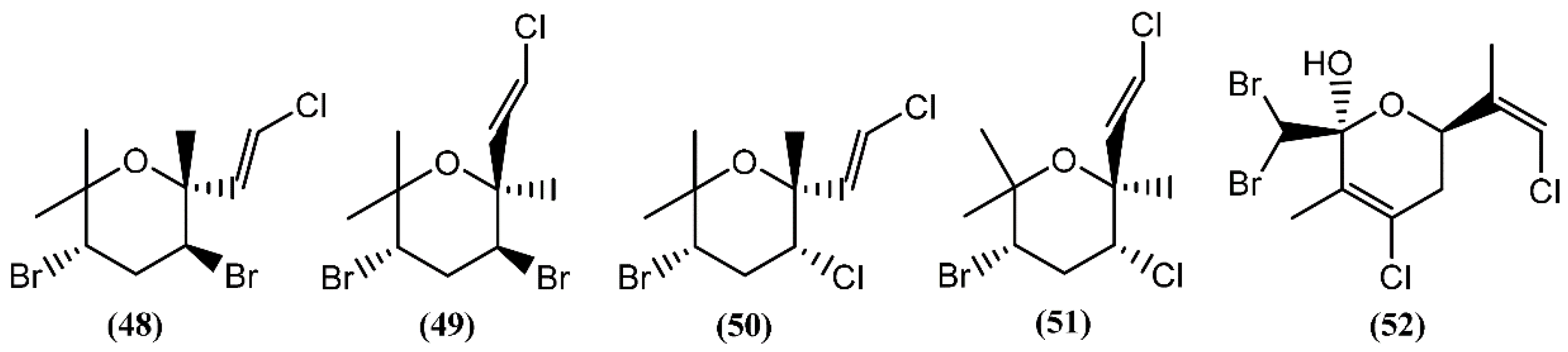
6.1. Monoterpenes
6.2. Sesquiterpenes
6.2.1. Chamigrane Skeleton Sesquiterpenes
6.2.2. Bisabolane and Cuparane Skeleton Sesquiterpenes
6.2.3. Laurane Skeleton Sesquiterpenes
6.2.4. Brasilane and Omphalane Skeleton Sesquiterpenes
6.2.5. Eudesmane and Snyderane Skeleton Sesquiterpenes
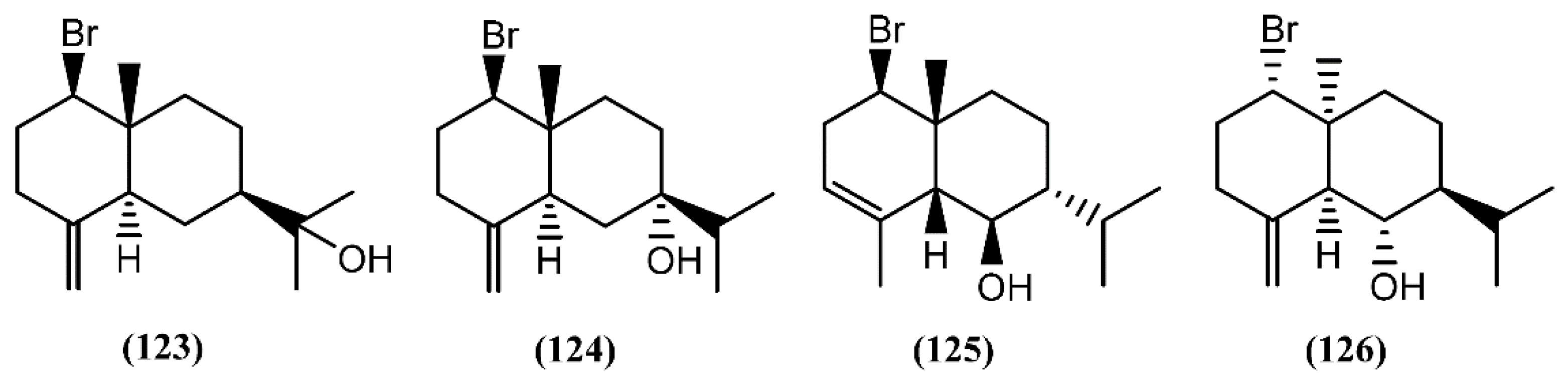
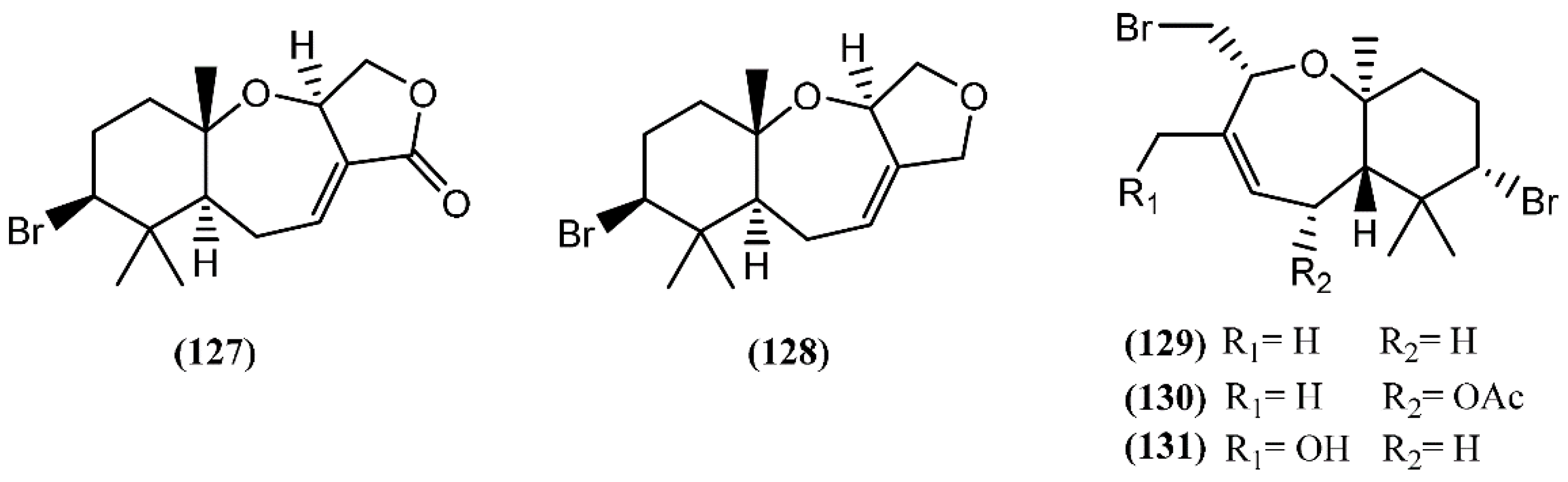
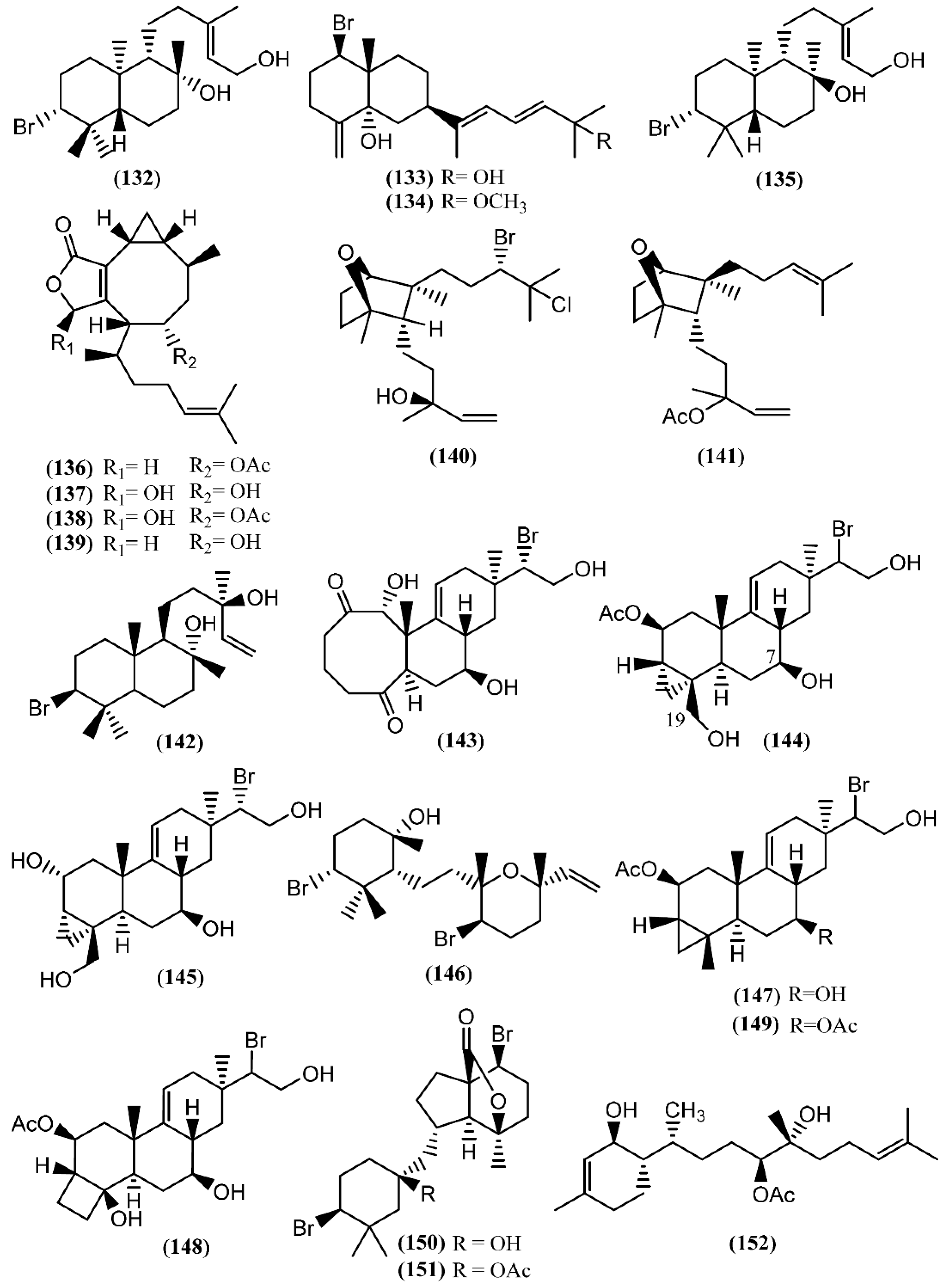
6.3. Diterpenes
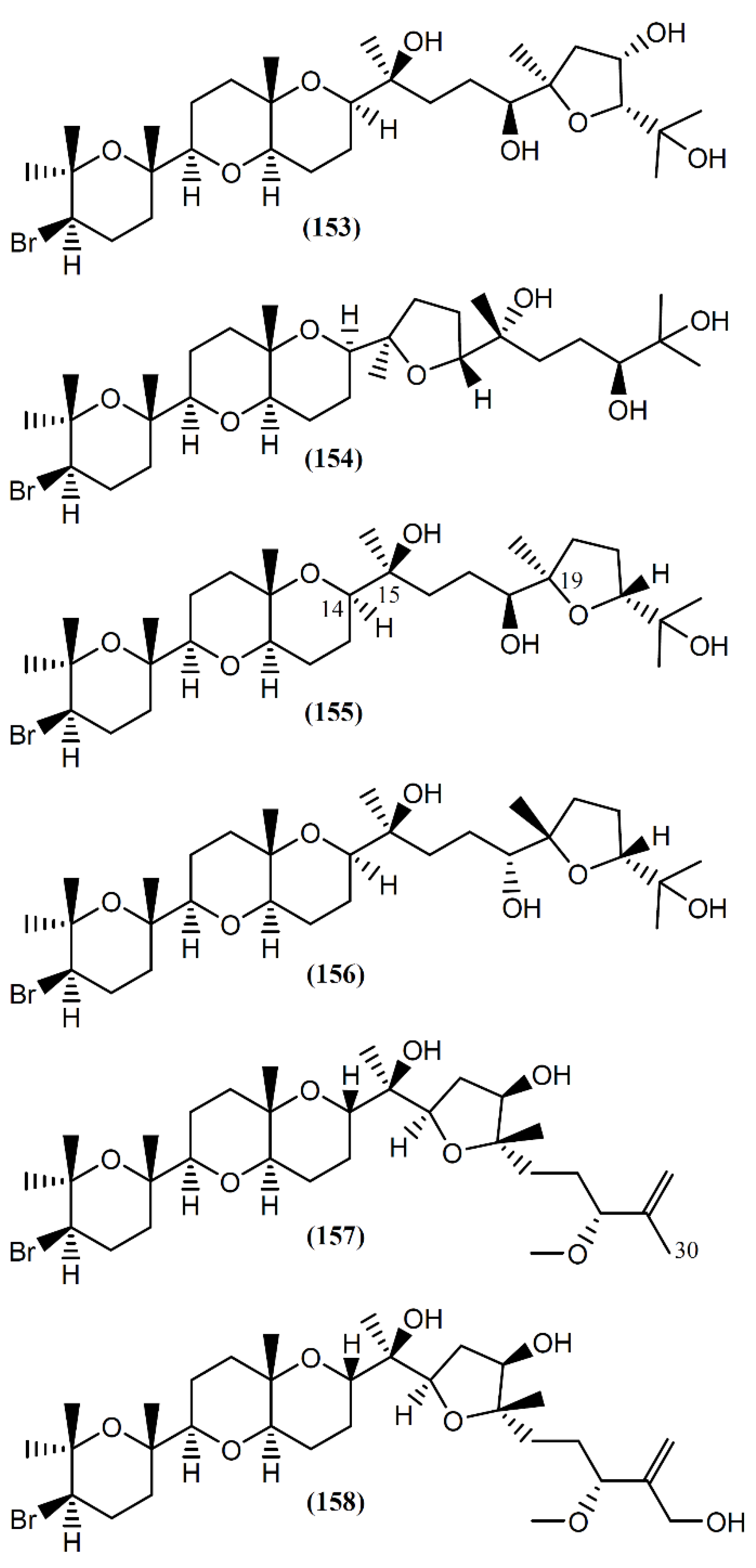
6.4. Triterpenes
7. Other Metabolites
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Lebar, M.D.; Heimbegner, J.L.; Baker, B.J. Cold-water marine natural products. Nat. Prod. Rep. 2007, 24, 774–797. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hu, G.; Yu, J.; Zhu, X.; Lin, Y.; Chen, S.; Yuan, J. Statistical research on the bioactivity of new marine natural products discovered during the 28 years from 1985 to 2012. Mar. Drugs 2015, 13, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.P.; Yuan, J.; Sun, L.; She, Z.G.; Wu, J.H.; Lan, X.J.; Zhu, X.; Lin, Y.C.; Chen, S.P. Statistical research on marine natural products based on data obtained between 1985 and 2008. Mar. Drugs 2011, 9, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K. Molluscan biological and chemical diversity: Secondary metabolites and medicinal resources produced by marine molluscs. Biol. Rev. Camb. Philos. Soc. 2010, 85, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tuinman, A.A.; Boettner, F.E.; Kizu, H.; Schmidt, J.M.; Baczynskyj, L.; Tomer, K.B.; Bontems, R.J. The isolation and structure of a remarkable marine animal antineoplastic constituent: Dolastatin 10. J. Am. Chem. Soc. 1987, 109, 6883–6885. [Google Scholar] [CrossRef]
- Luesch, H.; Moore, R.E.; Paul, V.J.; Mooberry, S.L.; Corbett, T.H. Isolation of dolastatin 10 from the marine cyanobacterium Symploca species VP642 and total stereochemistry and biological evaluation of its analogue symplostatin 1. J. Nat. Prod. 2001, 64, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine-sourced anti-cancer and cancer pain control agents in clinical and late preclinical development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.; Janik, J.E.; Younes, A. Brentuximab Vedotin (SGN-35). Clin. Cancer Res. 2011, 17, 6428–6436. [Google Scholar] [CrossRef] [PubMed]
- Derby, C.D. Escape by inking and secreting: Marine molluscs avoid predators through a rich array of chemicals and mechanisms. Biol. Bull. 2007, 213, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Calin-Jageman, R.J.; Fischer, T.M. Behavioral adaptation of the Aplysia siphon-withdrawal response is accompanied by sensory adaptation. Behav. Neurosci. 2007, 121, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Love-Chezem, T.; Aggio, J.F.; Derby, C.D. Defense through sensory inactivation: Sea hare ink reduces sensory and motor responses of spiny lobsters to food odors. J. Exp. Biol. 2013, 216, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Berriman, J.S.; Kay, M.C.; Reed, D.C.; Rassweiler, A.; Goldstein, D.A.; Wright, W.G. Shifts in attack behavior of an important kelp forest predator within marine reserves. Mar. Ecol. Prog. Ser. 2015, 522, 193–201. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Mees, J.; Costello, M.J.; Hernandez, F.; Bailly, N.; Boury-Esnault, N.; Gofas, S.; Horton, T.; Klautau, M.; Kroh, A.; et al. World Register of Marine Species. Available online: http://www.marinespecies.org/aphia.php?p=taxlist (accessed on 27 January 2016).
- Leal, M.C.; Puga, J.; Serôdio, J.; Gomes, N.C.; Calado, R. Trends in the discovery of new marine natural products from invertebrates over the last two decades—where and what are we bioprospecting? PLoS ONE 2012, 7, e30580. [Google Scholar] [CrossRef] [PubMed]
- Pennings, S.C.; Paul, V.J. Sequestration of dietary secondary metabolites by three species of sea hares: Location, specificity and dynamics. Mar. Biol. 1993, 117, 535–546. [Google Scholar] [CrossRef]
- Palaniveloo, K.; Vairappan, C.S. Chemical relationship between red algae genus Laurencia and sea hare Aplysia dactylomela Rang in the North Borneo Island. J. Appl. Phycol. 2014, 26, 1199–1205. [Google Scholar] [CrossRef]
- Yamamura, S.; Hirata, Y. Structures of aplysin and aplysinol, naturally occurring bromo-compounds. Tetrahedron 1963, 19, 1485–1496. [Google Scholar] [CrossRef]
- Kamiya, H.; Sakai, R.; Jimbo, M. Bioactive molecules from sea hares. In Molluscs: From Chemo-Ecological Study to Biotechnological Application; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin, Germany, 2006; pp. 215–234. [Google Scholar]
- Pereira, R.B.; Taveira, M.; Valentão, P.; Sousa, C.; Andrade, P.B. Fatty acids from edible sea hares: Anti-inflammatory capacity in LPS-stimulated RAW 264.7 cells involves iNOS modulation. RSC Adv. 2015, 5, 8981–8987. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Lobo-da-Cunha, A.; Taveira, M.; Ferreira, M.; Valentão, P.; Andrade, P.B. Digestive gland from Aplysia depilans Gmelin: Leads for inflammation treatment. Molecules 2015, 20, 15766–15780. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, J.G.; Daranas, A.H.; Norte, M.; Fernández, J.J. Marine macrolides, a promising source of antitumor compounds. Anticancer Agents Med. Chem. 2009, 9, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ma, S. The medicinal potential of promising marine macrolides with anticancer activity. Chem. Med. Chem. 2011, 6, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Ojika, M.; Ishigaki, T.; Yoshida, Y.; Ekimoto, H.; Arakawa, M. Aplyronine A, a potent antitumor substance and the congeners aplyronines B and C isolated from the sea hare Aplysia kurodai. J. Am. Chem. Soc. 1993, 115, 11020–11021. [Google Scholar]
- Ojika, M.; Kigoshi, H.; Suenaga, K.; Imamura, Y.; Yoshikawa, K.; Ishigaki, T.; Sakakura, A.; Mutou, T.; Yamada, K. Aplyronines D–H from the sea hare Aplysia kurodai: Isolation, structures, and cytotoxicity. Tetrahedron 2012, 68, 982–987. [Google Scholar] [CrossRef]
- Kita, M.; Kigoshi, H. Marine natural products that interfere with multiple cytoskeletal protein interactions. Nat. Prod. Rep. 2015, 32, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Watabe, S.; Ozaki, H.; Kigoshi, H.; Yamada, K.; Fusetani, N.; Karaki, H. Novel actin depolymerizing macrolide aplyronine A. J. Biochem. 1996, 120, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Ojika, M.; Kigoshi, H.; Suenaga, K. Aplyronine A, a potent antitumor macrolide of marine origin, and the congeners aplyronines B–H: Chemistry and biology. Nat. Prod. Rep. 2009, 26, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Kita, M.; Hirayama, Y.; Yoneda, K.; Yamagishi, K.; Chinen, T.; Usui, T.; Sumiya, E.; Uesugi, M.; Kigoshi, H. Inhibition of microtubule assembly by a complex of actin and antitumor macrolide aplyronine A. J. Am. Chem. Soc. 2013, 135, 18089–18095. [Google Scholar] [CrossRef] [PubMed]
- Kita, M.; Yoneda, K.; Hirayama, Y.; Yamagishi, K.; Saito, Y.; Sugiyama, Y.; Miwa, Y.; Ohno, O.; Morita, M.; Suenaga, K.; et al. Fluorescent aplyronine A: Intracellular accumulation and disassembly of actin cytoskeleton in tumor cells. ChemBioChem 2012, 13, 1754–1758. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, W.H. Carbocyclic oxylipins of marine origin. Chem. Rev. 1993, 93, 1807–1823. [Google Scholar] [CrossRef]
- Spinella, A.; Zubía, E.; Martînez, E.; Ortea, J.; Cimino, G. Structure and stereochemistry of aplyolides A-E, lactonized dihydroxy fatty acids from the skin of the marine mollusc Aplysia depilans. J. Org. Chem. 1997, 62, 5471–5475. [Google Scholar] [CrossRef]
- Stallard, M.O.; Faulkner, D.J. Chemical constituents of the digestive gland of the sea hare Aplysia californica I. Importance of diet. Comp. Biochem. Physiol. B 1974, 49, 25–35. [Google Scholar] [CrossRef]
- Kaul, P.N. Compounds from the sea with actions on the cardiovascular and central nervous systems. Fed. Proc. 1981, 40, 10–14. [Google Scholar] [PubMed]
- Ciavatta, M.L.; Gavagnin, M.; Puliti, R.; Cimino, G.; Martínez, E.; Ortea, J.; Mattia, C.A. Dactylallene: A novel dietary C15 bromoallene from the atlantic anaspidean mollusc Aplysia dactylomela. Tetrahedron 1997, 53, 17343–17350. [Google Scholar] [CrossRef]
- McDonald, F.J.; Campbell, D.C.; Vanderah, D.J.; Schmitz, F.J.; Washecheck, D.M.; Burks, J.E.; van der Helm, D. Marine natural products. Dactylyne, an acetylenic dibromochloro ether from the sea hare Aplysia dactylomela. J. Org. Chem. 1975, 40, 665–666. [Google Scholar] [CrossRef]
- Kaul, P.N.; Kulkarni, S.K. New drug metabolism inhibitor of marine origin. J. Pharm. Sci. 1978, 67, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.K.; Kaul, P.N. Substances of marine origin affecting pentobarbital pharmacokinetics. Indian J. Exp. Biol. 1980, 18, 270–272. [Google Scholar] [PubMed]
- Gao, L.-X.; Murai, A. Total synthesis of (−)-dactylyne and (−)-isodactylyne. Tetrahedron Lett. 1992, 33, 4349–4352. [Google Scholar] [CrossRef]
- Ioannou, E.; Nappo, M.; Avila, C.; Vagias, C.; Roussis, V. Metabolites from the sea hare Aplysia fasciata. J. Nat. Prod. 2009, 72, 1716–1719. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.A.; Li, G. Novel terpenoids from the sea hare Aplysia punctata. Can. J. Chem. 2002, 80, 1697–1707. [Google Scholar] [CrossRef]
- Kigoshi, H.; Shizuri, Y.; Niwa, H.; Yamada, K. Laurencenyne, a plausible precursor of various nonterpenoid C15-compounds, and neolaurencenyne from the red alga Laurencia okamurai. Tetrahedron Lett. 1981, 22, 4729–4732. [Google Scholar] [CrossRef]
- Okamoto, Y.; Nitanda, N.; Ojika, M.; Sakagami, Y. Aplysiallene, a new bromoallene as an Na+, K+-ATPase inhibitor from the sea hare, Aplysia kurodai. Biosci. Biotechnol. Biochem. 2001, 65, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kurosawa, E. A C15 non-terpenoid from the red alga Laurencia okamurai. Phytochemistry 1985, 24, 1999–2002. [Google Scholar] [CrossRef]
- Wang, J.; Pagenkopf, B.L. First total synthesis and structural reassignment of (−)-aplysiallene. Org. Lett. 2007, 9, 3703–3706. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Ebisawa, Y.; Higuchi, R. Aplyparvunin, a bioactive acetogenin from the sea hare Aplysia parvula. Tetrahedron Lett. 1995, 36, 6073–6074. [Google Scholar] [CrossRef]
- Manzo, E.; Ciavatta, M.L.; Gavagnin, M.; Puliti, R.; Mollo, E.; Guo, Y.-W.; Mattia, C.A.; Mozzarella, L.; Cimino, G. Structure and absolute stereochemistry of novel C15-halogenated acetogenins from the anaspidean mollusc Aplysia dactylomela. Tetrahedron 2005, 61, 7456–7460. [Google Scholar] [CrossRef]
- Falshaw, C.P.; King, T.J.; Imre, S.; Islimyeli, S.; Thomson, R.H. Laurenyne, a new acetylene from Laurencia obtusa: Crystal structure and absolute configuration. Tetrahedron Lett. 1980, 21, 4951–4954. [Google Scholar] [CrossRef]
- Takahashi, Y.; Daitoh, M.; Suzuki, M.; Abe, T.; Masuda, M. Halogenated metabolites from the new Okinawan red alga Laurencia yonaguniensis. J. Nat. Prod. 2002, 65, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kurosawa, E.; Furusaki, A.; Matsumoto, T. The structures of (3Z)-epoxyvenustin, (3Z)-venustin, and (3Z)-venustinene, new halogenated C15-nonterpenoids from the red alga Laurencia venusta Yamada. Chem. Lett. 1983, 5, 779–782. [Google Scholar] [CrossRef]
- McPhail, K.L.; Davies-Coleman, M.T. (3Z)-Bromofucin from a South African sea hare. Nat. Prod. Res. 2005, 19, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Coll, J.C.; Wright, A.D. Tropical Marine Algae. IV. Novel metabolites from the red alga Laurencia implicata (Rhodophyta, Rhodophyceae, Ceramiales, Rhodomelaceae). Aust. J. Chem. 1989, 42, 1685–1693. [Google Scholar] [CrossRef]
- Kinnel, R.B.; Dieter, R.K.; Meinwald, J.; van Engen, D.; Clardy, J.; Eisner, T.; Stallard, M.O.; Fenical, W. Brasilenyne and cis-dihydrorhodophytin: Antifeedant medium-ring haloethers from a sea hare Aplysia brasiliana. Proc. Natl. Acad. Sci. USA 1979, 76, 3576–3579. [Google Scholar] [CrossRef] [PubMed]
- Kigoshi, H.; Imamura, Y.; Yoshikawa, K.; Yamada, K. Three new cytotoxic alkaloids, aplaminone, neoaplaminone and neoaplaminone sulfate from the marine mollusc Aplysia kurodai. Tetrahedron Lett. 1990, 31, 4911–4914. [Google Scholar] [CrossRef]
- Yamada, K.; Ojika, M.; Kigoshi, H.; Suenaga, K. Cytotoxic substances from two species of Japanese sea hares: Chemistry and bioactivity. Proc. Jpn. Acad. Ser. B 2010, 86, 176–189. [Google Scholar] [CrossRef]
- Kigoshi, H.; Adachi, Y.; Yoshikawa, K.; Yamada, K. Absolute stereochemistry of aplaminone and neoaplaminone, cytotoxic bromodopamines from a marine mollusc: Enantioselective synthesis of debromoneoaplaminone. Tetrahedron Lett. 1992, 33, 4195–4198. [Google Scholar] [CrossRef]
- Ojika, M.; Yoshida, T.; Yamada, K. Aplysepine, a novel 1,4-benzodiazepine alkaloid from the sea hare Aplysia kurodai. Tetrahedron Lett. 1993, 34, 5307–5308. [Google Scholar] [CrossRef]
- Appleton, D.R.; Babcock, R.C.; Copp, B.R. Novel tryptophan-derived dipeptides and bioactive metabolites from the sea hare Aplysia dactylomela. Tetrahedron 2001, 57, 10181–10189. [Google Scholar] [CrossRef]
- Kuroda, T.; Kigoshi, H. Aplaminal: A novel cytotoxic aminal isolated from the sea hare Aplysia kurodai. Org. Lett. 2008, 10, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.B.; Liu, Z. Total synthesis of (−)-aplaminal. Org. Lett. 2008, 10, 4363–4365. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Tang, H.; Wang, P.; Gong, W.; Xue, M.; Zhang, H.; Liu, T.; Liu, B.; Yi, Y.; Zhang, W. Bioactive polyoxygenated steroids from the South China Sea Soft Coral, Sarcophyton sp. Mar. Drugs 2013, 11, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Yen, W.-H.; Chen, W.-F.; Cheng, C.-H.; Dai, C.-F.; Lu, M.-C.; Su, J.-H.; Su, Y.-D.; Chen, Y.-H.; Chang, Y.-C.; Chen, Y.-H.; et al. A new 5α,8α-epidioxysterol from the Soft Coral Sinularia gaweli. Molecules 2013, 18, 2895–2903. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Ibrahim, A.; Arafa, A.S. Anti-H5N1 virus metabolites from the Red Sea soft coral, Sinularia candidula. Tetrahedron Lett. 2013, 54, 2377–2381. [Google Scholar] [CrossRef]
- Felder, S.; Kehraus, S.; Neu, E.; Bierbaum, G.; Schaberle, T.F.; Konig, G.M. Salimyxins and enhygrolides: Antibiotic, sponge-related metabolites from the obligate marine myxobacterium Enhygromyxa salina. ChemBioChem 2013, 14, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.; Quiñoá, E.; Castedo, L.; Riguera, R. Epidioxy sterols from the tunicates Dendrodoa grossularia and Ascidiella aspersa and the gastropoda Aplysia depilans and Aplysia punctata. J. Nat. Prod. 1986, 49, 905–909. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Nakanishi, Y.; Shimokawa, T.; Hashiguchi, S.; Hayashi, A. Structure elucidation of oxygenated sterols from eggs of sea hare.Aplysia Juliana. Chem. Lett. 1992, 21, 1713–1714. [Google Scholar] [CrossRef]
- Mun, B.; Wang, W.; Kim, H.; Hahn, D.; Yang, I.; Won, D.H.; Kim, E.-H.; Lee, J.; Han, C.; Kim, H.; et al. Cytotoxic 5α,8α-epidioxy sterols from the marine sponge Monanchora sp. Arch. Pharm. Res. 2015, 38, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.E.; Capper, A.; Della Togna, G.; Paul, V.J.; Romero, L.I.; Johns, T.; Cubilla-Rios, L.; Capson, T.L. Ecology- and bioassay-guided drug discovery for treatments of tropical parasitic disease: 5alpha, 8alpha-epidioxycholest-6-en-3beta-ol isolated from the mollusk Dolabrifera dolabrifera shows significant activity against Leishmania donovani. Nat. Prod. Commun. 2013, 8, 1537–1540. [Google Scholar] [PubMed]
- Miyamoto, T.; Higuchi, R.; Komori, T.; Fujioka, T.; Mihashi, K. Isolation and structures of aplykurodins A and B, two new isoprenoids from the marine mollusk Aplysia kurodai. Tetrahedron Lett. 1986, 27, 1153–1156. [Google Scholar] [CrossRef]
- Spinella, A.; Gavagnin, M.; Crispino, A.; Cimino, G.; Martinez, E.; Ortea, J.; Sodano, G. 4-acetylaplykurodin B and aplykurodinone B, two ichthyotoxic degraded sterols from the mediterranean mollusk Aplysia fasciata. J. Nat. Prod. 1992, 55, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Izzo, I.; Meduri, G.; Avallone, E.; De Riccardis, F.; Sodano, G. Studies towards the synthesis of aplykurodins-Synthesis of 17,18-dihydro-3,9-di-epi-aplykurodinone B. Eur. J. Org. Chem. 2000, 2000, 439–448. [Google Scholar] [CrossRef]
- Shoji, N.; Umeyama, A.; Shin, K.; Takeda, K.; Arihara, S.; Kobayashi, J.; Takei, M. Two unique pentacyclic steroids with cis C/D ring junction from Xestospongia bergquistii Fromont, powerful inhibitors of histamine release. J. Org. Chem. 1992, 57, 2996–2997. [Google Scholar] [CrossRef]
- Ortega, M.J.; Zubía, E.; Salvá, J. 3-epi-aplykurodinone B, a new degraded sterol from Aplysia fasciata. J. Nat. Prod. 1997, 60, 488–489. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, A.; Kita, M.; Kigoshi, H. Aplysiasecosterol A: A 9,11-secosteroid with an unprecedented tricyclic γ-diketone structure from the sea hare Aplysia kurodai. Angew. Chem. Int. Ed. Engl. 2015, 54, 7073–7076. [Google Scholar] [CrossRef] [PubMed]
- Fraga, B.M. Natural sesquiterpenoids. Nat. Prod. Rep. 2010, 27, 1681–1708. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, H.; Sakai, R.; Jimbo, M. Bioactive molecules from sea hares. Prog. Mol. Subcell. Biol. 2006, 43, 215–239. [Google Scholar] [PubMed]
- Pennings, S.C.; Weiss, A.M.; Paul, V.J. Secondary metabolites of the cyanobacterium Microcoleus lyngbyaceus and the sea hare Stylocheilus longicauda: Palatability and toxicity. Mar. Biol. 1996, 126, 735–743. [Google Scholar] [CrossRef]
- Stallard, M.O.; Faulkner, D.J. Chemical constituents of the digestive gland of the sea hare Aplysia californica. II. Importance of diet. Comp. Biochem. Physiol. B. 1974, 49, 37–41. [Google Scholar] [PubMed]
- Wessels, M.; Konig, G.M.; Wright, A.D. New natural product isolation and comparison of the secondary metabolite content of three distinct samples of the sea hare Aplysia dactylomela from Tenerife. J. Nat. Prod. 2000, 63, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, T.; Uchida, H.; Inouye, Y.; Ishitsuka, M.; Yamamoto, H.; Kakisawa, H. Novel cytotoxic monoterpenes having a halogenated tetrahydropyran from Aplysia kurodai. J. Org. Chem. 1987, 52, 4597–4600. [Google Scholar] [CrossRef]
- Jung, M.E.; Lew, W. Efficient total synthesis of the cytotoxic halogenated monoterpene aplysiapyranoid D. J. Org. Chem. 1991, 56, 1347–1349. [Google Scholar] [CrossRef]
- Jung, M.E.; D’Amico, D.C.; Lew, W. Efficient total synthesis of the cytotoxic halogenated monoterpene aplysiapyranoid A. Tetrahedron Lett. 1993, 34, 923–926. [Google Scholar] [CrossRef]
- Jung, M.E.; Fahr, B.T.; D’Amico, D.C. Total syntheses of the cytotoxic marine natural product, aplysiapyranoid C. J. Org. Chem. 1998, 63, 2982–2987. [Google Scholar] [CrossRef]
- Grkovic, T.; Appleton, D.R.; Copp, B.R. Chemistry and chemical ecology of some of the common opisthobranch molluscs found on the shores of New Zealand. Chem. N. Z. 2005, 69, 12–15. [Google Scholar]
- Stierle, D.B.; Wing, R.M.; Sims, J.J. Marine natural products. Costatone and costatolide, new halogenated monoterpenes from the red seaweed, Plocamium costatum. Tetrahedron Lett. 1976, 17, 4455–4458. [Google Scholar] [CrossRef]
- Kazlauskas, R.; Murphy, P.T.; Quinn, R.J.; Wells, R.J.; Schönhlzer, P.; F-Konzern, J.S.; Roche, F.H.-L. Two polyhalogenated monoterpenes from the red alga Plocamium costatum. Tetrahedron Lett. 1976, 17, 4451–4454. [Google Scholar] [CrossRef]
- Motti, C.A.; Thomas-Hall, P.; Hagiwara, K.A.; Simmons, C.J.; Willis, R.; Wright, A.D. Accelerated identification of halogenated monoterpenes from Australian specimens of the red algae Plocamium hamatum and Plocamium costatum. J. Nat. Prod. 2014, 77, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Williard, P.G.; Grab, L.A. Total synthesis of costatone: A monoterpene from the red seaweed: Plocarnium costatum. Tetrahedron Lett. 1984, 25, 5009–5012. [Google Scholar] [CrossRef]
- Quiñoa, E.; Castedo, L.; Riguera, R. The halogenated monoterpenes of Aplysia punctata. A comparative study. Comp. Biochem. Physiol. B 1989, 92, 99–101. [Google Scholar] [CrossRef]
- Miyamoto, T.; Higuchi, R.; Marubayashi, N.; Komori1, T. Studies on the constituents of marine opisthobranchia, IV. Two new polyhalogenated monoterpenes from the sea hare Aplysia kurodai. Liebigs Ann. Chem. 1988, 1988, 1191–1193. [Google Scholar] [CrossRef]
- Watanabe, K.; Umeda, K.; Kurita, Y.; Takayama, C.; Miyakado, M. Two insecticidal monoterpenes, telfairine and aplysiaterpenoid A, from the red alga Plocamium telfairiae: Structure elucidation, biological activity, and molecular topographical consideration by a semiempirical molecular orbital study. Pestic. Biochem. Physiol. 1990, 37, 275–286. [Google Scholar] [CrossRef]
- Faulkner, D.J.; Stallard, M.O. 7-chloro-3,7-dimethyl-1,4,6-tribromo-1-octen-3-ol, a novel monoterpene alcohol from Aplysia californica. Tetrahedron Lett. 1973, 14, 1171–1174. [Google Scholar] [CrossRef]
- Katayama, A.; Ina, K.; Nozaki, H.; Nakayama, M. Structural elucidation of kurodainol, a novel halogenated monoterpene from sea hare Aplysia Kurodai. Agric. Biol. Chem. 1982, 46, 859–860. [Google Scholar] [CrossRef]
- Ortega, M.J.; Zubía, E.; Salvá, J. New polyhalogenated monoterpenes from the sea hare Aplysia punctata. J. Nat. Prod. 1997, 60, 482–484. [Google Scholar] [CrossRef] [PubMed]
- Conolly, J.D.; Hill, R.A. Mono and sesquiterpenoids. In Dictionary of Terpenoids, 1st ed.; Champman and Hall: London, UK, 1991; Volume 1, pp. 1–153. [Google Scholar]
- Dias, T.; Brito, I.; Moujir, L.; Paiz, N.; Darias, J.; Cueto, M. Cytotoxic sesquiterpenes from Aplysia dactylomela. J. Nat. Prod. 2005, 68, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Desoti, V.C.; Lazarin-Bidóia, D.; Sudatti, D.B.; Pereira, R.C.; Alonso, A.; Ueda-Nakamura, T.; Dias Filho, B.P.; Nakamura, C.V.; De Silva, S.O. Trypanocidal action of (−)-elatol involves an oxidative stress triggered by mitochondria dysfunction. Mar. Drugs 2012, 10, 1631–1646. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.O.; Veiga-Santos, P.; Ueda-Nakamura, T.; Dias Filho, B.P.; Sudatti, D.B.; Bianco, É.M.; Pereira, R.C.; Nakamura, C.V. Effect of elatol, isolated from red seaweed Laurencia dendroidea, on Leishmania amazonensis. Mar. Drugs 2010, 8, 2733–2743. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Souza, C.B.; Lhullier, C.; Falkenberg, M.; Schenkel, E.P.; Ribeiro-Do-Valle, R.M.; Siqueira, J.M. Anti-tumor effects of elatol, a marine derivative compound obtained from red algae Laurencia microcladia. J. Pharm. Pharmacol. 2012, 64, 1146–1154. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Daitoh, M.; Suzuki, M.; Abe, T.; Masuda, M. Antibacterial halogenated metabolites from the Malaysian Laurencia species. Phytochemistry 2001, 58, 291–297. [Google Scholar] [CrossRef]
- Bianco, E.M.; Pires, L.; Santos, G.K.N.; Dutra, K.A.; Reis, T.N.V.; Vasconcelos, E.R.T.P.P.; Cocentino, A.L.M.; Navarro, D.M.A.F. Larvicidal activity of seaweeds from northeastern Brazil and of a halogenated sesquiterpene against the dengue mosquito (Aedes aegypti). Ind. Crops Prod. 2013, 43, 270–275. [Google Scholar] [CrossRef]
- Desoti, V.C.; Lazarin-Bidóia, D.; Sudatti, D.B.; Pereira, R.C.; Ueda-Nakamura, T.; Nakamura, C.V.; de Oliveira Silva, S. Additional evidence of the trypanocidal action of (−)-elatol on amastigote forms through the involvement of reactive oxygen species. Mar. Drugs 2014, 12, 4973–4983. [Google Scholar] [CrossRef] [PubMed]
- König, G.M.; Wright, A.D. Laurencia rigida: Chemical investigations of its antifouling dichloromethane extract. J. Nat. Prod. 1997, 60, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S.; Anangdan, S.P.; Matsunaga, S. Diet-derived halogenated metabolite from the sea hare Aplysia parvula. Malays. J. Sci. 2009, 28, 269–273. [Google Scholar]
- Díaz-Marrero, A.R.; De La Rosa, J.M.; Brito, I.; Darias, J.; Cueto, M. Dactylomelatriol, a biogenetically intriguing omphalane-derived marine sesquiterpene. J. Nat. Prod. 2012, 75, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Vairappan, C.S. Potent antibacterial activity of halogenated metabolites from Malaysian red algae, Laurencia majuscula (Rhodomelaceae, Ceramiales). Biomol. Eng. 2003, 20, 255–259. [Google Scholar] [CrossRef]
- Da Silva Machado, F.L.; Pacienza-Lima, W.; Rossi-Bergmann, B.; De Souza Gestinari, L.M.; Fujii, M.T.; Campos De Paula, J.; Costa, S.S.; Lopes, N.P.; Kaiser, C.R.; Soares, A.R. Antileishmanial sesquiterpenes from the Brazilian red alga Laurencia dendroidea. Planta Med. 2011, 77, 733–735. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, M.-E.F.; Moustfa, A.Y.; Mohamed, A.E.-H.H.; Alhammady, M.A.; Elbehairi, S.E.I.; Ohta, S.; Paré, P.W. New cytotoxic halogenated sesquiterpenes from the Egyptian sea hare, Aplysia oculifera. Tetrahedron Lett. 2014, 55, 1711–1714. [Google Scholar] [CrossRef]
- Kaiser, C.R.; Pitombo, L.F.; Pinto, A.C. C-13 and H-1 NMR assignments of the chamigrenes prepacifenol and dehydroxyprepacifenol epoxides. Spectrosc. Lett. 1998, 31, 573–585. [Google Scholar] [CrossRef]
- Ireland, C.; Stallard, M.O.; Faulkner, D.J.; Finer, J.; Clardy, J. Some chemical constituents of the digestive gland of the sea hare Aplysia californica. J. Org. Chem. 1976, 41, 2461–2465. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, D.J.; Stallard, M.O.; Ireland, C. Prefacifenol epoxide, a halogenated sesquiterpene diepoxide. Tetrahedron Lett. 1974, 15, 3571–3574. [Google Scholar] [CrossRef]
- Pitombo, L.F.; Kaiser, C.R.; Pinto, A.C. Occurrence of chamigrenes in Aplysia dactylomela from Brazilian waters. Bol. Soc. Chil. Quim. 1996, 41, 433–436. [Google Scholar]
- Sims, J.J.; Fenical, W.; Wing, R.M.; Radlick, P. Marine natural products. I. Pacifenol, a rare sesquiterpene containing bromine and chlorine from the red alga, Laurencia pacifica. J. Am. Chem. Soc. 1971, 93, 3774–3775. [Google Scholar] [CrossRef]
- Kaiser, C.R.; Pitombo, L.F.; Pinto, A.C. Complete 1H and 13C NMR assignments of chamigrenes from Aplysia dactylomela. Magn. Reson. Chem. 2001, 39, 147–149. [Google Scholar] [CrossRef]
- San-Martín, A.; Rovirosa, J.; Astudillo, L.; Sepulveda, B.; Ruiz, D.; San-Martin, C. Biotransformation of the marine sesquiterpene pacifenol by a facultative marine fungus. Nat. Prod. Res. 2008, 22, 1627–1632. [Google Scholar] [CrossRef] [PubMed]
- Jongaramruong, J.; Blackman, A.J.; Skelton, B.W.; White, A.H. Chemical relationships between the sea hare Aplysia parvula and the red seaweed Laurencia filiformis from Tasmania. Aust. J. Chem. 2002, 55, 275–280. [Google Scholar] [CrossRef]
- Gil, B.; Ferrándiz, M.L.; Sanz, M.J.; Terencio, M.C.; Ubeda, A.; Rovirosa, J.; San-Martin, A.; Alcaraz, M.J.; Payá, M. Inhibition of inflammatory responses by epitaondiol and other marine natural products. Life Sci. 1995, 57, 25–30. [Google Scholar] [CrossRef]
- Pietra, F. Evolution of the secondary metabolite versus evolution of the species. Pure Appl. Chem. 2003, 74, 2207–2211. [Google Scholar] [CrossRef]
- McPhail, K.L.; Davies-Coleman, M.T.; Copley, R.C.B.; Eggleston, D.S. New halogenated sesquiterpenes from South African specimens of the circumtropical sea hare Aplysia dactylomela. J. Nat. Prod. 1999, 62, 1618–1623. [Google Scholar] [CrossRef]
- Kimura, J; Kamada, N.; Tsujimoto, Y. Fourteen chamigrane derivatives from a red alga, Laurencia nidifica. Bull. Chem. Soc. Jpn. 1999, 72, 289–292. [Google Scholar] [CrossRef]
- Fraga, B.M. Natural sesquiterpenoids. Nat. Prod. Rep. 1997, 14, 145–162. [Google Scholar] [CrossRef]
- González, A.G.; Darias, J.; Estèvez, E. Chemotherapeutic activity of polyhalogenated terpenes from Spanish algae. Planta Med. 1982, 44, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Konig, G.M. Terpenoids from marine organisms: Unique structures and their pharmacological potential. Phytochem. Rev. 2006, 5, 115–141. [Google Scholar] [CrossRef]
- Lhullier, C.; Falkenberg, M.; Ioannou, E.; Quesada, A.; Papazafiri, P.; Horta, P.A.; Schenkel, E.P.; Vagias, C.; Roussis, V. Cytotoxic halogenated metabolites from the Brazilian red alga Laurencia catarinensis. J. Nat. Prod. 2010, 73, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Brito, I.; Dias, T.; Díaz-Marrero, A.R.; Darias, J.; Cueto, M. Aplysiadiol from Aplysia dactylomela suggested a key intermediate for a unified biogenesis of regular and irregular marine algal bisabolene-type metabolites. Tetrahedron 2006, 62, 9655–9660. [Google Scholar] [CrossRef]
- Hollenbeak, K.H.; Schmitz, F.J.; Hossain, M.B.; Van Der Helm, D. Deodactol, antineoplastic sesquiterpenoid from the sea hare Aplysia dactylomela. Tetrahedron 1979, 35, 541–545. [Google Scholar] [CrossRef]
- Harinantenaina, L.; Kurata, R.; Asakawa, Y. Chemical constituents of Malagasy liverworts, part III: Sesquiterpenoids from Bazzania decrescens and Bazzania madagassa. Chem. Pharm. Bull. 2005, 53, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Guella, G.; Öztunç, A.; Mancini, I.; Pietra, F. Stereochemical features of sesquiterpene metabolites as a distinctive trait of red seaweeds in the genus Laurencia. Tetrahedron Lett. 1997, 38, 8261–8264. [Google Scholar] [CrossRef]
- Ichiba, T.; Higa, T. New cuparene-derived sesquiterpenes with unprecedented oxygenation pattern from the sea hare Aplysia dactylomela. J. Org. Chem. 1986, 51, 3364–3366. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Ishii, T.; Lee, T.K.; Suzuki, M.; Zhaoqi, Z. Antibacterial activities of a new brominated diterpene from Borneon Laurencia spp. Mar. Drugs 2010, 8, 1743–1749. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Yamashita, Y.; Ohta, T. New cytotoxic and antibacterial compounds isolated from the sea hare, Aplysia kurodai. Mar. Drugs 2005, 3, 22–28. [Google Scholar] [CrossRef]
- Kim, M.M.; Mendis, E.; Kim, S.K. Laurencia okamurai extract containing laurinterol induces apoptosis in melanoma cells. J. Med. Food 2008, 11, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liang, H.; Su, A.; He, J.; Yu, H. Effect of aplysin on the proliferation and apoptosis in human gastric cancer cell SGC-7901. Chin. Pharmacol. Bull. 2010, 26, 333–337. [Google Scholar]
- Gong, A.; Ge, N.; Yao, W.; Lu, L.; Liang, H. Aplysin enhances temozolomide sensitivity in glioma cells by increasing miR-181 level. Cancer Chemother. Pharmacol. 2014, 74, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Gong, A.-J.; Gong, L.-L.; Yao, W.-C.; Ge, N.; Lu, L.-X.; Liang, H. Aplysin induces apoptosis in glioma cells through HSP90/AKT pathway. Exp. Biol. Med. 2015, 240, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Ge, N.; Liang, H.; Liu, Y.; Ma, A.-G.; Han, L. Protective effect of aplysin on hepatic injury in ethanol-treated rats. Food Chem. Toxicol. 2013, 62, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Copley, R.C.; Davies-Coleman, M.T.; Edmonds, D.R.; Faulkner, D.J.; McPhail, K.L. Absolute stereochemistry of ibhayinol from a South African sea hare. J. Nat. Prod. 2002, 65, 580–582. [Google Scholar] [CrossRef] [PubMed]
- König, G.M.; Wright, A.D. Sesquiterpene content of the antibacterial dichloromethane extract of the marine red alga Laurencia obtusa. Planta Med. 1997, 63, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Lake, R.J.; Munro, M.H.G. Sesquiterpenes from the marine red alga Laurencia distichophylla. Phytochemistry 1984, 23, 1951–1954. [Google Scholar] [CrossRef]
- Gewali, M.B.; Ronald, R.C. Synthesis of allolaurinterol. J. Org. Chem. 1982, 47, 2792–2795. [Google Scholar] [CrossRef]
- Stallard, M.O.; Fenical, W.; Kittredge, J.S. The brasilenols, rearranged sesquiterpene alcohols isolated from the marine opisthobranch Aplysia brasiliana. Tetrahedron 1978, 34, 2077–2081. [Google Scholar] [CrossRef]
- Tori, M.; Nakashima, K.; Asakawa, Y. Sesquiterpenes and a phenolic compound from the liverwort Omphalanthus filiformis. Phytochemistry 1995, 38, 651–653. [Google Scholar] [CrossRef]
- Dieter, R.K.; Kinnel, R.; Meinwald, J.; Eisner, T. Brasudol and isobrasudol: Two bromosesquiterpenes from a sea hare Aplysia brasiliana. Tetrahedron Lett. 1979, 20, 1645–1648. [Google Scholar] [CrossRef]
- Brennan, M.R.; Erickson, K.L. Austradiol acetate and austradiol diacetate, 4,6-dihydroxy-(+)-selinane derivatives from an Australian Laurencia sp. J. Org. Chem. 1982, 47, 3917–3921. [Google Scholar] [CrossRef]
- Sun, H.H.; Waraszkiewicz, S.M.; Erickson, K.L. Sesquiterpenoid alcohols from the Hawaiian marine alga Laurencia nidifica. Tetrahedron Lett. 1976, 17, 585–588. [Google Scholar] [CrossRef]
- Suzuki, T.; Kikuchi, H.; Kurosawa, E. Six new sesquiterpenoids from the red alga Laurencia nipponica Yamada. Bull. Chem. Soc. Jpn. 1982, 55, 1561–1563. [Google Scholar] [CrossRef]
- Baker, B.; Ratnapala, L.; Mahindaratne, M.P.D.; de Silva, E.D.; Tillekeratne, L.M.V.; Jeong, J.H.; Scheuer, P.J.; Seff, K. Lankalapuol A and B: Two cis-eudesmanes from the sea hare Aplysia dactylomela. Tetrahedron 1988, 44, 4695–4701. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Anangdan, S.P.; Lee, T.K. Additional halogenated secondary metabolites from the sea hare Aplysia dactylomela. Malays. J. Sci. 2007, 26, 57–64. [Google Scholar]
- Pettit, G.R.; Herald, C.L.; Allen, M.S.; von Dreele, R.B.; Vanell, L.D.; Kao, J.P.; Blake, W. The isolation and structure of aplysistatin. J. Am. Chem. Soc. 1977, 99, 262–263. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.N.; de Nys, R.; Steinberg, P.D. Effects of algal diet on the performance and susceptibility to predation of the sea hare Aplysia parvula. Mar. Ecol. Prog. Ser. 2002, 236, 241–254. [Google Scholar] [CrossRef]
- Vairappan, C.S.; Kamada, T.; Lee, W.-W.; Jeon, Y.-J. Anti-inflammatory activity of halogenated secondary metabolites of Laurencia snackeyi (Weber-van Bosse) Masuda in LPS-stimulated RAW 264.7 macrophages. J. Appl. Phycol. 2013, 25, 1805–1813. [Google Scholar] [CrossRef]
- Matsuda, H.; Tomiie, Y.; Yamamura, S.; Hirata, Y. The structure of aplysin-20. Chem. Commun. (Lond.) 1967, 17, 898b–899b. [Google Scholar] [CrossRef]
- Ojika, M.; Yoshida, Y.; Okumura, M.; Ieda, S.; Yamada, K. Aplysiadiol, a new brominated diterpene from the marine mollusc Aplysia kurodai. J. Nat. Prod. 1990, 53, 1619–1622. [Google Scholar] [CrossRef]
- Takahashi, Y.; Suzuki, M.; Abe, T.; Masuda, M. Anhydroaplysiadiol from Laurencia japonensis. Phytochemistry 1998, 48, 987–990. [Google Scholar] [CrossRef]
- Ojika, M.; Kigoshi, H.; Yoshikawa, K.; Yamada, K. A new bromo diterpene, epi-aplysin-20, and ent-isoconcinndiol from the marine mollusc Aplysia kurodai. Bull. Chem. Soc. Jpn. 1992, 65, 2300–2302. [Google Scholar] [CrossRef]
- Midland, S.L.; Wing, R.M.; Sims, J.J. New crenulides from the sea hare Aplysia vaccaria. J. Org. Chem. 1983, 48, 1906–1909. [Google Scholar] [CrossRef]
- Sun, H.H.; McEnroe, F.J.; Fenical, W. Acetoxycrenulide, a new bicyclic cyclopropane-containing diterpenoid from the brown seaweed Dictyota crenulata. J. Org. Chem. 1983, 48, 1903–1906. [Google Scholar] [CrossRef]
- Othmani, A.; Bouzidi, N.; Viano, Y.; Alliche, Z.; Seridi, H.; Blache, Y.; El Hatta, M.; Briand, J.-F.; Culioli, G. Anti-microfouling properties of compounds isolated from several Mediterranean Dictyota spp. J. Appl. Phycol. 2014, 26, 1573–1584. [Google Scholar] [CrossRef]
- Bouaicha, N.; Pesando, D.; Puel, D.; Tringali, C. Cytotoxic diterpenoids from the brown alga Dilophus ligulatus. J. Nat. Prod. 1993, 56, 1747–1752. [Google Scholar] [CrossRef] [PubMed]
- Ko, R.K.; Kang, M.C.; Kim, S.S.; Oh, T.H.; Kim, G.O.; Hyun, C.G.; Hyun, J.W.; Lee, N.H. Anti-melanogenesis constituents from the seaweed Dictyota coriacea. Nat. Prod. Commun. 2013, 8, 427–428. [Google Scholar] [PubMed]
- Estrada, D.M.; Ravelo, J.L.; Ruiz-Pérez, C.; Martín, J.D.; Solans, X. Dactylomelol, a new class of diterpene from the sea hare Aplysia dactylomela. Tetrahedron Lett. 1989, 30, 6219–6220. [Google Scholar] [CrossRef]
- Fernández, J.J.; Souto, M.L.; Gil, L.V.; Norte, M. Isolation of naturally occurring dactylomelane metabolites as Laurencia constituents. Tetrahedron 2005, 61, 8910–8915. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Michaud, D.P.; Schmidt, P.G. Marine natural products: Parguerol, deoxyparguerol, and isoparguerol. New brominated diterpenes with modified pimarane skeletons from the sea hare Aplysia dactylomela. J. Am. Chem. Soc. 1982, 104, 6415–6423. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Yamashita, Y.; Yoshida, T.; Ohta, T. Parguerol and isoparguerol isolated from the sea hare, Aplysia kurodai, induce neurite outgrowth in PC-12 Cells. Mar. Drugs 2004, 2, 170–175. [Google Scholar] [CrossRef]
- Awad, N.E. Bioactive brominated diterpenes from the marine red alga Jania rubens (L.) Lamx. Phytother. Res. 2004, 18, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Herald, C.L.; Einck, J.J.; Vanell, L.D.; Brown, P.; Gust, D. Isolation and structure of angasiol. J. Org. Chem. 1978, 43, 4685–4686. [Google Scholar] [CrossRef]
- Atta-Ur-Rahmanc; Alvi, K.A.; Abbas, S.A.; Sultana, T.; Shameel, M.; Iqbal Choudhary, M.; Clardy, J.C. A diterpenoid lactone from Aplysia Juliana. J. Nat. Prod. 1991, 54, 886–888. [Google Scholar] [CrossRef]
- Jiménez-Romero, C.; Mayer, A.M.S.; Rodríguez, A.D. Dactyloditerpenol acetate, a new prenylbisabolane-type diterpene from Aplysia dactylomela with significant in vitro anti-neuroinflammatory activity. Bioorg. Med. Chem. Lett. 2014, 24, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S. Therapeutic implications of microglia activation by lipopolysaccharide and reactive oxygen species generation in septic shock and central nervous system pathologies: A review. Medicina 1998, 58, 377–385. [Google Scholar] [PubMed]
- Kim, S.-K.; Himaya, S.W.A. Triterpene glycosides from sea cucumbers and their biological activities. Adv. Food Nutr. Res. 2012, 65, 297–319. [Google Scholar] [PubMed]
- Sultana, N.; Saify, Z.S. Naturally occurring and synthetic agents as potential anti-inflammatory and immunomodulants. Antiinflamm. Antiallergy Agents Med. Chem. 2012, 11, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.-A.; Zhou, Q.; Guo, W.-Z.; Qiu, Y.; Wang, R.; Jin, M.; Zhang, W.; Li, K.; Yamori, T.; Dan, S.; et al. In vitro antitumor activity of stellettin B, a triterpene from marine sponge Jaspis stellifera, on human glioblastoma cancer SF295 cells. Mar. Drugs 2014, 12, 4200–4213. [Google Scholar] [CrossRef] [PubMed]
- Akl, M.R.; Foudah, A.I.; Ebrahim, H.Y.; Meyer, S.A.; El Sayed, K.A. The marine-derived sipholenol A-4-O-3′,4′-dichlorobenzoate inhibits breast cancer growth and motility in vitro and in vivo through the suppression of Brk and FAK signaling. Mar. Drugs 2014, 12, 2282–2304. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, H.; Tian, X.; Lin, H.; Wang, M.; Yao, M. Three new cytotoxic isomalabaricane triterpenes from the marine sponge Stelletta tenuis. Fitoterapia 2015, 106, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Manzo, E.; Gavagnin, M.; Bifulco, G.; Cimino, P.; Di Micco, S.; Ciavatta, M.L.; Guo, Y.W.; Cimino, G. Aplysiols A and B, squalene-derived polyethers from the mantle of the sea hare Aplysia dactylomela. Tetrahedron 2007, 63, 9970–9978. [Google Scholar] [CrossRef]
- Vera, B.; Rodríguez, A.D.; Avilés, E.; Ishikawa, Y. Aplysqualenols A and B: Squalene-derived polyethers with antitumoral and antiviral activity from the Caribbean Sea slug Aplysia dactylomela. Eur. J. Org. Chem. 2009, 2009, 5327–5336. [Google Scholar] [CrossRef] [PubMed]
- Ola, A.R.B.; Babey, A.M.; Motti, C.; Bowden, B.F. Aplysiols CE, brominated triterpene polyethers from the marine alga Chondria armata and a revision of the structure of aplysiol B. Aust. J. Chem. 2010, 63, 907–914. [Google Scholar] [CrossRef]
- Cen-Pacheco, F.; Mollinedo, F.; Villa-Pulgarín, J.A.; Norte, M.; Fernández, J.J.; Daranas, A.H. Saiyacenols A and B: The key to solve the controversy about the configuration of aplysiols. Tetrahedron 2012, 68, 7275–7279. [Google Scholar] [CrossRef]
- Blunt, J.W.; Hartshorn, M.P.; McLennan, T.J.; Munro, M.H.G.; Robinson, W.T.; Yorke, S.C. Thyrsiferol: A squalene-derived metabolite of Laurencia thyrsifera. Tetrahedron Lett. 1978, 19, 69–72. [Google Scholar] [CrossRef]
- Sakemi, S.; Higa, T.; Jefford, C.W.; Bernardinelli, G. Venustatriol. A new, anti-viral, triterpene tetracyclic ether from Laurencia venusta. Tetrahedron Lett. 1986, 27, 4287–4290. [Google Scholar] [CrossRef]
- Fernández, J.J.; Souto, M.L.; Norte, M. Evaluation of the cytotoxic activity of polyethers isolated from Laurencia. Bioorg. Med. Chem. 1998, 6, 2237–2243. [Google Scholar] [CrossRef]
- Little, R.D.; Nishiguchi, G.A. Synthetic efforts toward, and biological activity of, thyrsiferol and structurally-related analogues. Stud. Nat. Prod. Chem. 2008, 35, 3–56. [Google Scholar]
- Mahdi, F.; Falkenberg, M.; Ioannou, E.; Roussis, V.; Zhou, Y.-D.; Nagle, D.G. Thyrsiferol inhibits mitochondrial respiration and HIF-1 activation. Phytochem. Lett. 2011, 4, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Poddar, M.; Aminabhavi, T.M.; Patel, M.; Singh, N.; Noolvi, M.N. HIF inhibitors: New hope for cancer therapy. Lett. Drug Des. Discov. 2015, 12, 736–753. [Google Scholar] [CrossRef]
- González, I.C.; Forsyth, C.J. Total synthesis of thyrsiferyl 23-acetate, a specific inhibitor of protein phosphatase 2A and an anti-leukemic inducer of apoptosis. J. Am. Chem. Soc. 2000, 122, 9099–9108. [Google Scholar] [CrossRef]
- Janssens, V.; Goris, J. Protein phosphatase 2A: A highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem. J. 2001, 353, 417–439. [Google Scholar] [CrossRef] [PubMed]
- Kuniyoshi, M.; Yamada, K.; Higa, T. A biologically active diphenyl ether from the green alga Cladophora fascicularis. Experientia 1985, 41, 523–524. [Google Scholar] [CrossRef]
- Kamio, M.; Nguyen, L.; Yaldiz, S.; Derby, C.D. How to produce a chemical defense: Structural elucidation and anatomical distribution of aplysioviolin and phycoerythrobilin in the sea hare Aplysia californica. Chem. Biodivers. 2010, 7, 1183–1197. [Google Scholar] [CrossRef] [PubMed]
- Kamio, M.; Grimes, T.V.; Hutchins, M.H.; van Dam, R.; Derby, C.D. The purple pigment aplysioviolin in sea hare ink deters predatory blue crabs through their chemical senses. Anim. Behav. 2010, 80, 89–100. [Google Scholar] [CrossRef]
- Sakai, S.; Komura, Y.; Nishimura, Y.; Sugawara, T.; Hirata, T. Inhibition of mast cell degranulation by phycoerythrin and its pigment moiety phycoerythrobilin, prepared from Porphyra yezoensis. Food Sci. Technol. Res. 2011, 17, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Przeslawski, R.; Benkendorff, K.; Davis, A.R. A quantitative survey of mycosporine-like amino acids (MAAS) in intertidal egg masses from temperate rocky shores. J. Chem. Ecol. 2005, 31, 2417–2438. [Google Scholar] [CrossRef] [PubMed]
- Kamio, M.; Kicklighter, C.E.; Nguyen, L.; Germann, M.W.; Derby, C.D. Isolation and structural elucidation of novel mycosporine-like amino acids as alarm cues in the defensive ink secretion of the sea hare Aplysia californica. Helv. Chim. Acta 2011, 94, 1012–1018. [Google Scholar] [CrossRef]
- Singh, S.P.; Kumari, S.; Rastogi, R.P.; Singh, K.L.; Sinha, R.P. Mycosporine-like amino acids (MAAs): Chemical structure, biosynthesis and significance as UV-absorbing/screening compounds. Indian J. Exp. Biol. 2008, 46, 7–17. [Google Scholar] [PubMed]
- Kicklighter, C.E.; Kamio, M.; Nguyen, L.; Germann, M.W.; Derby, C.D. Mycosporine-like amino acids are multifunctional molecules in sea hares and their marine community. Proc. Natl. Acad. Sci. USA 2011, 108, 11494–11499. [Google Scholar] [CrossRef] [PubMed]
- Cubillos, V.M.; Burritt, D.J.; Lamare, M.D.; Peake, B.M. The relationship between UV-irradiance, photoprotective compounds and DNA damage in two intertidal invertebrates with contrasting mobility characteristics. J. Photochem. Photobiol. B 2015, 149, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.C.M.; Alonso-Varona, A.; Palomares, T.; Zubillaga, V.; Labidi, J.; Bulone, V. Exploiting mycosporines as natural molecular sunscreens for the fabrication of UV-absorbing green materials. ACS Appl. Mater. Interfaces 2015, 7, 16558–16564. [Google Scholar] [CrossRef] [PubMed]
- Colabella, F.; Moliné, M.; Libkind, D. UV sunscreens of microbial origin: Mycosporines and mycosporine-like aminoacids. Recent Pat. Biotechnol. 2015, 8, 179–193. [Google Scholar] [CrossRef]
- Carefoot, T.H.; Harris, M.; Taylor, B.E.; Donovan, D.; Karentz, D. Mycosporine-like amino acids: Possible UV protection in eggs of the sea hare Aplysia dactylomela. Mar. Biol. 1998, 130, 389–396. [Google Scholar] [CrossRef]
- Carefoot, T.H.; Karentz, D.; Pennings, S.C.; Young, C.L. Distribution of mycosporine-like amino acids in the sea hare Aplysia dactylomela: Effect of diet on amounts and types sequestered over time in tissues and spawn. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2000, 126, 91–104. [Google Scholar] [CrossRef]
- De La Coba, F.; Aguilera, J.; Figueroa, F.L.; De Gálvez, M.V.; Herrera, E. Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J. Appl. Phycol. 2009, 21, 161–169. [Google Scholar] [CrossRef]
- Andreguetti, D.; Stein, E.M.; Pereira, C.M.; Pinto, E.; Colepicolo, P. Antioxidant properties and UV absorbance pattern of mycosporine-like amino acids analogs synthesized in an environmentally friendly manner. J. Biochem. Mol. Toxicol. 2013, 27, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, S.; Khodabandeh, S.; Agh, N.; Bakhtiarian, M. Effects of salinity and ultraviolet radiation on the bioaccumulation of mycosporine-like amino acids in Artemia from Lake Urmia. Photochem. Photobiol. 2013, 89, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Ojika, M.; Yoshida, Y.; Nakayama, Y.; Yamada, K. Aplydilactone, a novel fatty acid metabolite from the marine mollusc Aplysia kurodai. Tetrahedron Lett. 1990, 31, 4907–4910. [Google Scholar] [CrossRef]
- Yamada, K.; Kigoshi, H. Bioactive compounds from the sea hares of two genera: Aplysia and Dolabella. Bull. Chem. Soc. Jpn. 1997, 70, 1479–1489. [Google Scholar] [CrossRef]

























© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, R.B.; Andrade, P.B.; Valentão, P. Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus. Mar. Drugs 2016, 14, 39. https://doi.org/10.3390/md14020039
Pereira RB, Andrade PB, Valentão P. Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus. Marine Drugs. 2016; 14(2):39. https://doi.org/10.3390/md14020039
Chicago/Turabian StylePereira, Renato B., Paula B. Andrade, and Patrícia Valentão. 2016. "Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus" Marine Drugs 14, no. 2: 39. https://doi.org/10.3390/md14020039
APA StylePereira, R. B., Andrade, P. B., & Valentão, P. (2016). Chemical Diversity and Biological Properties of Secondary Metabolites from Sea Hares of Aplysia Genus. Marine Drugs, 14(2), 39. https://doi.org/10.3390/md14020039