Sequencing and Characterization of Novel PII Signaling Protein Gene in Microalga Haematococcus pluvialis

Abstract
:1. Introduction
2. Results
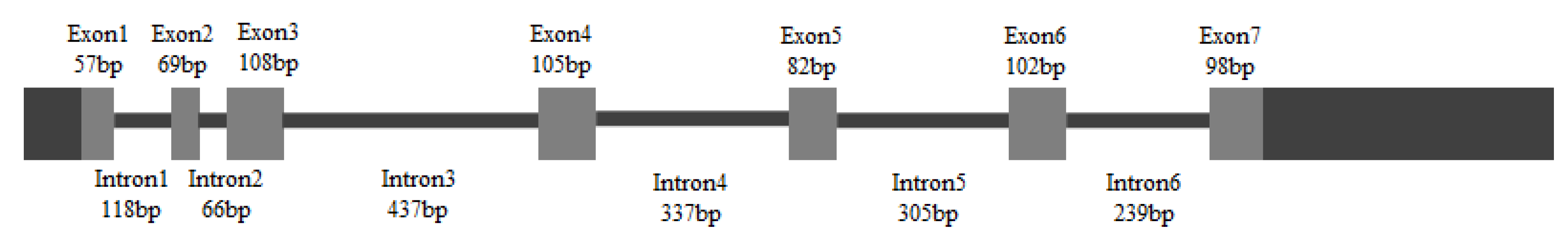
2.1. Cloning and Characterization of HpGLB1
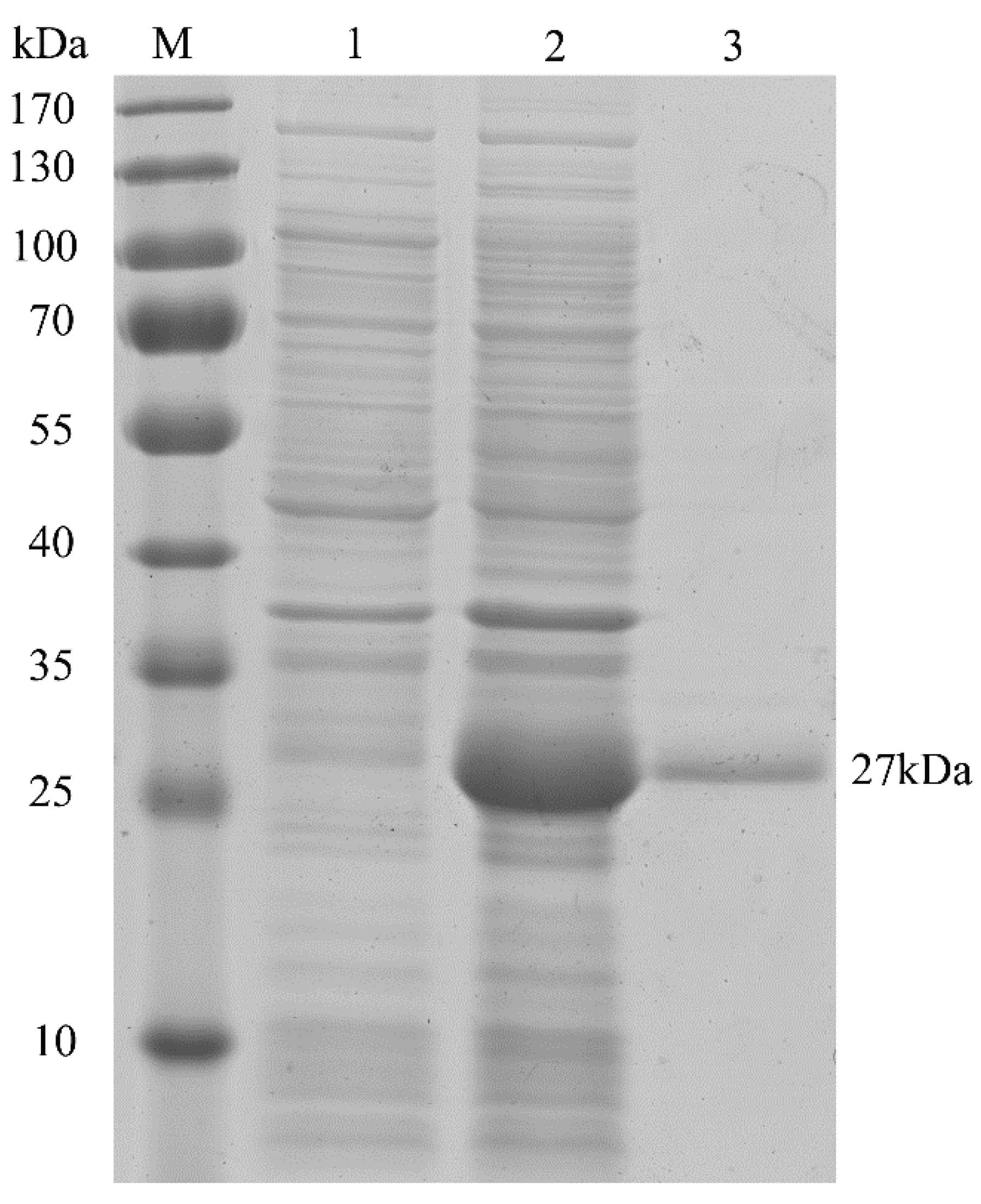
2.2. Characterization of HpPII Protein
2.3. Multiple Sequence Alignment and Structural Prediction
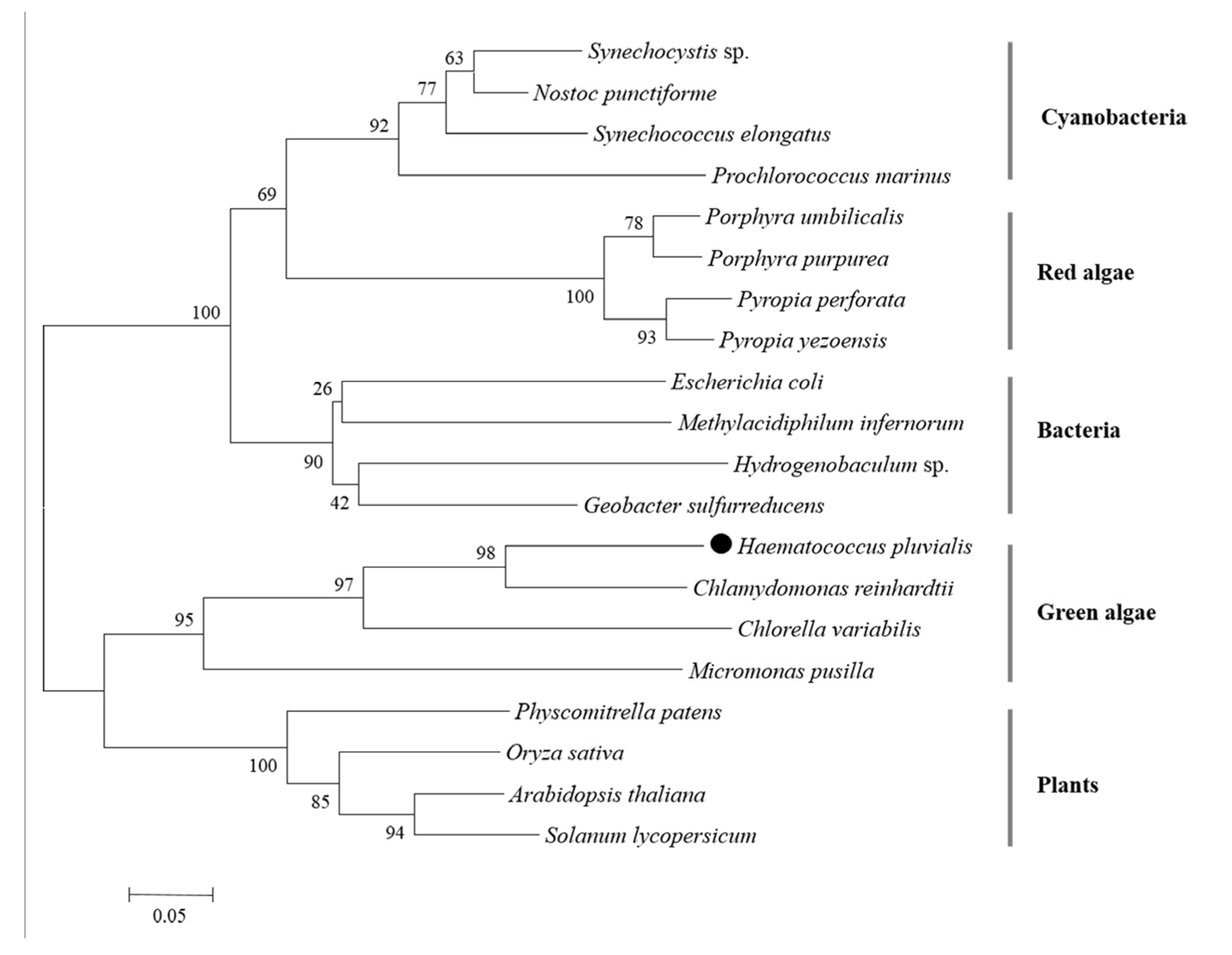
2.4. Phylogenetic Analysis
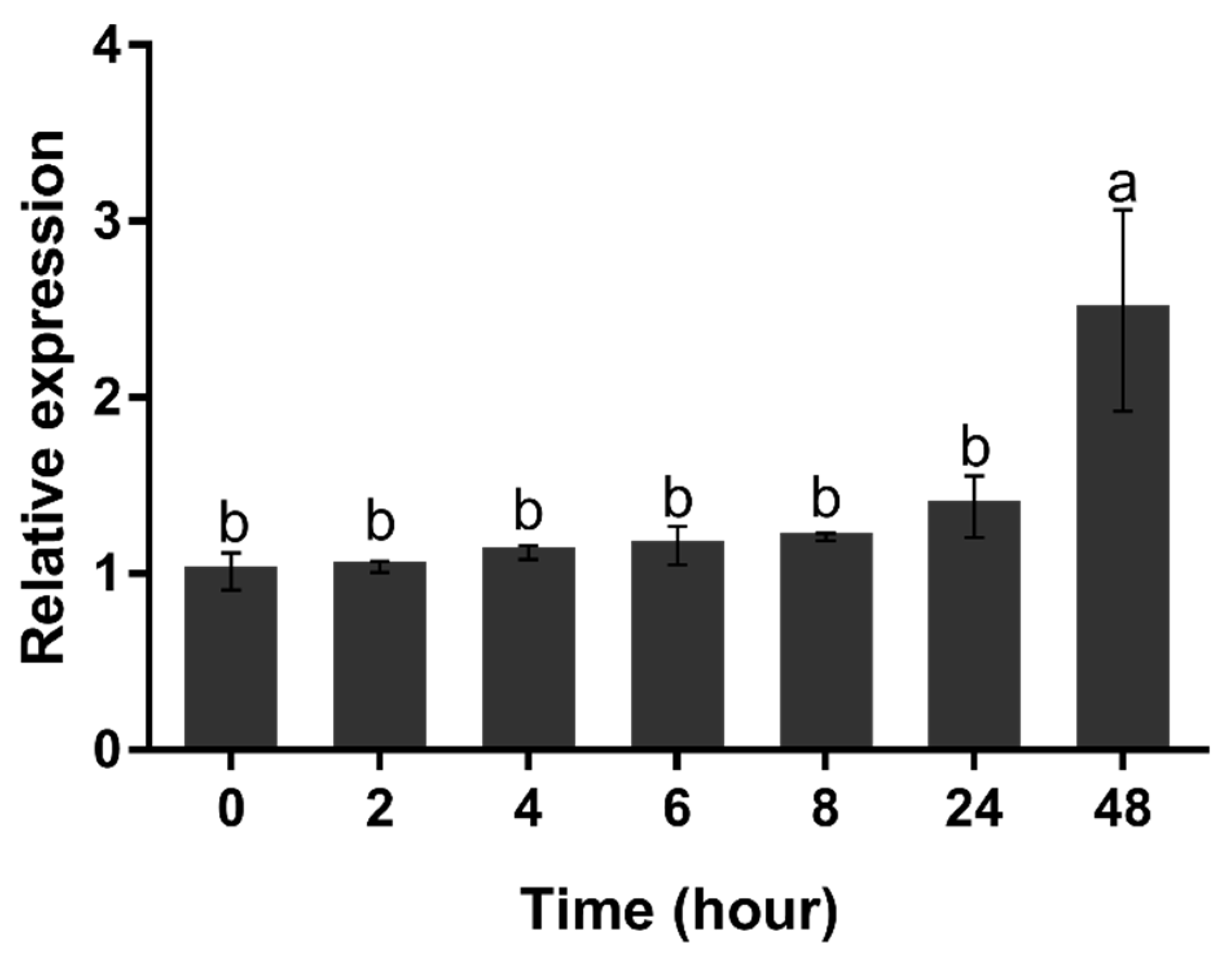
2.5. Transcription Analysis of HpGLB1 under Nitrogen Deprivation Condition
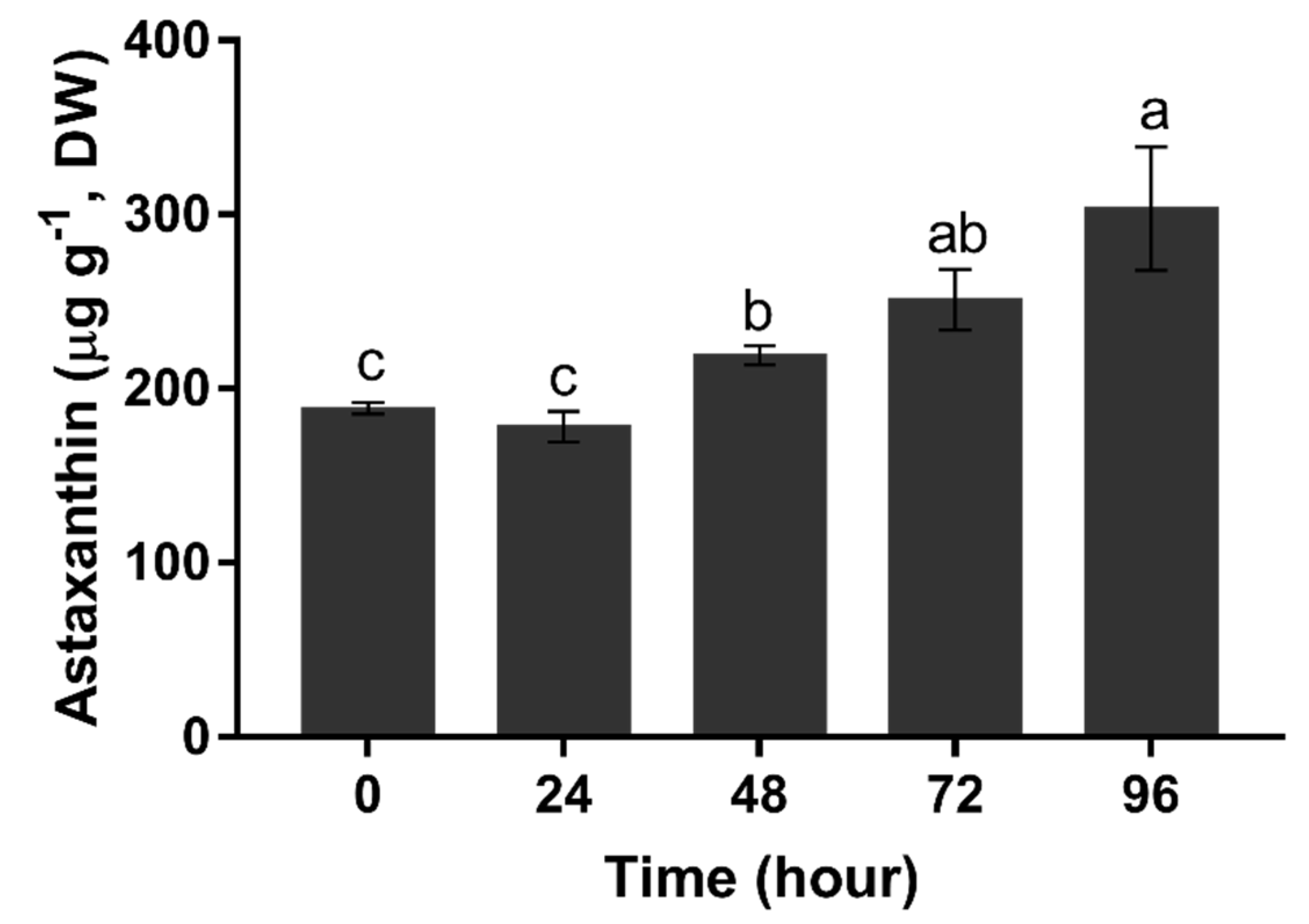
2.6. Astaxanthin Accumulation Pattern under Nitrogen Deprivation Condition
3. Discussion
4. Materials and Methods
4.1. Microalga Strain and Culture Conditions
4.2. RNA Isolation and cDNA Synthesis
4.3. Gene Cloning and Rapid Amplification of cDNA Ends (RACE)
4.4. Bioinformatics Analysis
4.5. Plasmid Construction
4.6. Expression and Purification of HpPII
4.7. Expression Analysis of HpGLB1 by qRT-PCR
4.8. Astaxanthin Extraction and Quantification
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mulders, K.J.M.; Weesepoel, Y.; Bodenes, P.; Lamers, P.P.; Vincken, J.P.; Martens, D.E.; Gruppen, H.; Wijffels, R.H. Nitrogen-depleted Chlorella zofingiensis produces astaxanthin, ketolutein and their fatty acid esters: A carotenoid metabolism study. J. Appl. Phycol. 2015, 27, 125–140. [Google Scholar] [CrossRef]
- Tran, N.P.; Park, J.K.; Lee, C.G. Proteomics analysis of proteins in green alga Haematococcus lacustris (chlorophyceae) expressed under combined stress of nitrogen starvation and high irradiance. Enzyme Microb. Technol. 2009, 45, 241–246. [Google Scholar] [CrossRef]
- Longworth, J.; Wu, D.Y.; Huete-Ortega, M.; Wright, P.C.; Vaidyanathan, S. Proteome response of Phaeodactylum tricornutum, during lipid accumulation induced by nitrogen depletion. Algal Res. 2016, 18, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Commichau, F.M.; Forchhammer, K.; Stulke, J. Regulatory links between carbon and nitrogen metabolism. Curr. Opin. Microbiol. 2006, 9, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Chellamuthu, V.R.; Ermilova, E.; Lapina, T.; Luddecke, J.; Minaeva, E.; Herrmann, C.; Hartmann, M.D.; Forchhammer, K. A widespread glutamine-sensing mechanism in the plant kingdom. Cell 2014, 159, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Uhrig, R.G.; Ng, K.K.S.; Moorhead, G.B.G. PII in higher plants: A modern role for an ancient protein. Trends Plant Sci. 2009, 14, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Ninfa, A.J.; Jiang, P. PII signal transduction proteins: Sensors of alpha-ketoglutarate that regulate nitrogen metabolism. Curr. Opin. Microbiol. 2005, 8, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Huergo, L.F.; Chandra, G.; Merrick, M. PII signal transduction proteins: Nitrogen regulation and beyond. FEMS Microbiol. Rev. 2013, 37, 251–283. [Google Scholar] [CrossRef] [PubMed]
- Helfmann, S.; Lu, W.; Litz, C.; Andrade, S.L.A. Cooperative binding of MgATP and MgADP in the trimeric PII protein GlnK2 from Archaeoglobus fulgidus. J. Mol. Biol. 2010, 402, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, E.C.; Araujo, L.M.; Ribeiro, R.R.; Chubatsu, L.S.; Scarduelli, M.; Rodrigues, T.E.; Monteiro, R.A.; Pedrosa, F.O.; Souza, E.M.; Huergo, L.F. Influence of the ADP/ATP ratio, 2-oxoglutarate and divalent ions on Azospirillum brasilense PII protein signalling. Microbiology 2012, 158, 1656–1663. [Google Scholar] [CrossRef] [PubMed]
- Burillo, S.; Luque, I.; Fuentes, I.; Contreras, A. Interactions between the nitrogen signal transduction protein PII and N-acetyl glutamate kinase in organisms that perform oxygenic photosynthesis. J. Bacteriol. 2004, 186, 3346–3354. [Google Scholar] [CrossRef] [PubMed]
- Bourrellier, A.B.F.; Valot, B.; Guillot, A.; Ambard-Bretteville, F.; Vidal, J.; Hodges, M. Chloroplast acetyl-CoA carboxylase activity is 2-oxoglutarate-regulated by interaction of PII with the biotin carboxyl carrier subunit. Proc. Natl. Acad. Sci. USA 2010, 107, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, E.C.M.; Rodrigues, T.E.; Muller-Santos, M.; Pedrosa, F.O.; Souza, E.M.; Forchhammer, K.; Huergo, L.F. The bacterial signal transduction protein GlnB regulates the committed step in fatty acid biosynthesis by acting as a dissociable regulatory subunit of acetyl-CoA carboxylase. Mol. Microbiol. 2015, 95, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Minaeva, E.; Ermilova, E. Sequencing and expression analysis of the gene encoding PII signal protein in Chlorella variabilis NC64A. J. Plant Biochem. Biotechnol. 2015, 2. [Google Scholar] [CrossRef]
- Ermilova, E.; Lapina, T.; Zalutskaya, Z.; Minaeva, E.; Fokina, O.; Forchhammer, K. PII signal transduction protein in Chlamydomonas reinhardtii: Localization and expression pattern. Protist 2013, 164, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Gwak, Y.; Hwang, Y.S.; Wang, B.B.; Kim, M.; Jeong, J.; Lee, C.G.; Hu, Q.; Han, D.X.; Jin, E. Comparative analyses of lipidomes and transcriptomes reveal a concerted action of multiple defensive systems against photooxidative stress in Haematococcus pluvialis. J. Exp. Bot. 2014, 65, 4317–4334. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.T.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef]
- Regnier, P.; Bastias, J.; Rodriguez-Ruiz, V.; Caballero-Casero, N.; Caballo, C.; Sicilia, D.; Fuentes, A.; Maire, M.; Crepin, M.; Letourneur, D.; et al. Astaxanthin from Haematococcus pluvialis prevents oxidative stress on human endothelial cells without toxicity. Mar. Drugs 2015, 13, 2857–2874. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.E.; Hwang, S.K.; Chang, W.S.; Kim, B.W.; Lee, J.; Sim, S.J. Enhanced autotrophic astaxanthin production from Haematococcus pluvialis under high temperature via heat stress-driven Haber-Weiss reaction. Appl. Microbiol. Biotechnol. 2015, 99, 5203–5215. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.Q.; Meng, C.X.; Chen, Y.C.; Ahmed, F.; Mangott, A.; Schenk, P.M.; Li, Y. Comparison of astaxanthin accumulation and biosynthesis gene expression of three Haematococcus pluvialis strains upon salinity stress. J. Appl. Phycol. 2015, 27, 1853–1860. [Google Scholar] [CrossRef]
- Scibilia, L.; Girolomoni, L.; Berteotti, S.; Alboresi, A.; Ballottari, M. Photosynthetic response to nitrogen starvation and high light in Haematococcus pluvialis. Algal Res. 2015, 12, 170–181. [Google Scholar] [CrossRef]
- Liang, C.W.; Zhao, F.Q.; Meng, C.X.; Tan, C.P.; Qin, S. Molecular cloning, characterization and evolutionary analysis of phytoene desaturase (pds) gene from Haematococcus pluvialis. World J. Microbiol. Biotechnol. 2006, 22, 59–64. [Google Scholar] [CrossRef]
- Cui, H.L.; Yu, X.N.; Wang, Y.; Cui, Y.L.; Li, X.Q.; Liu, Z.P.; Qin, S. Gene cloning and expression profile of a novel carotenoid hydroxylase (CYP97C) from the green alga Haematococcus pluvialis. J. Appl. Phycol. 2014, 26, 91–103. [Google Scholar] [CrossRef]
- Wang, J.X.; Sommerfeld, M.; Hu, Q. Cloning and expression of isoenzymes of superoxide dismutase in Haematococcus pluvialis (chlorophyceae) under oxidative stress. J. Appl. Phycol. 2011, 23, 995–1003. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Tao, M.; Li, Z.; Hu, Z.L. Cloning and characterization of selenoprotein thioredoxin reductase gene in Haematococcus pluvialis. Algal Res. 2016, 17, 97–104. [Google Scholar] [CrossRef]
- Recht, L.; Topfer, N.; Batushansky, A.; Sikron, N.; Gibon, Y.; Fait, A.; Nikoloski, Z.; Boussiba, S.; Zarka, A. Metabolite profiling and integrative modeling reveal metabolic constraints for carbon partitioning under nitrogen starvation in the green algae Haematococcus pluvialis. J. Biol. Chem. 2014, 289, 30387–30403. [Google Scholar] [CrossRef] [PubMed]
- Fabregas, J.; Dominguez, A.; Alvarez, D.G.; Lamela, T.; Otero, A. Induction of astaxanthin accumulation by nitrogen and magnesium deficiencies in Haematococcus pluvialis. Biotechnol. Lett. 1998, 20, 623–626. [Google Scholar] [CrossRef]
- Boussiba, S.; Bing, W.; Yuan, J.P.; Zarka, A.; Chen, F. Changes in pigments profile in the green alga Haeamtococcus pluvialis exposed to environmental stresses. Biotechnol. Lett. 1999, 21, 601–604. [Google Scholar] [CrossRef]
- Forchhammer, K.; Tandeau de Marsac, N. The PII protein in the cyanobacterium Synechococcus sp. strain PCC 7942 is modified by serine phosphorylation and signals the cellular N-status. J. Bacteriol. 1994, 176, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Moorhead, G.B.G.; Ng, K.K.S. Structural basis for the regulation of N-acetylglutamate kinase by PII in Arabidopsis thaliana. J. Biol. Chem. 2007, 282, 35733–35740. [Google Scholar] [CrossRef] [PubMed]
- Forchhammer, K. Global carbon/nitrogen control by PII signal transduction in cyanobacteria: From signals to targets. FEMS Microbiol. Rev. 2004, 28, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Ferrario-Mery, S.; Bouvet, M.; Leleu, O.; Savino, G.; Hodges, M.; Meyer, C. Physiological characterisation of Arabidopsis mutants affected in the expression of the putative regulatory protein PII. Planta 2005, 223, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Olguin, E.J.; Dorantes, E.; Castillo, O.S.; Hernandez-Landa, V.J. Anaerobic digestates from vinasse promote growth and lipid enrichment in Neochloris oleoabundans cultures. J. Appl. Phycol. 2015, 27, 1813–1822. [Google Scholar] [CrossRef]
- Xie, Y.P.; Ho, S.H.; Chen, C.N.N.; Chen, C.Y.; Ng, I.S.; Jing, K.J.; Chang, J.S.; Lu, Y.H. Phototrophic cultivation of a thermo-tolerant Desmodesmus sp. for lutein production: Effects of nitrate concentration, light intensity and fed-batch operation. Bioresour. Technol. 2013, 144, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Varela-Alvarez, E.; Andreakis, N.; Lago-Leston, A.; Pearson, G.A.; Serrao, E.A.; Procaccini, G.; Duarte, C.M.; Marba, N. Genomic DNA isolation from green and brown algae (Caulerpales and Fucales) for microsatellite library construction. J. Phycol. 2006, 42, 741–745. [Google Scholar] [CrossRef]
- Ng, I.S.; Chi, X.Q.; Wu, X.M.; Bao, Z.W.; Lu, Y.H.; Chang, J.S.; Ling, X.P. Cloning and expression of Cel8A from Klebsiella pneumoniae in Escherichia coli and comparison to cel gene of Cellulomonas uda. Biochem. Eng. J. 2013, 78, 53–58. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) | Sequence Information |
|---|---|---|
| GLB1-de-F | AGCGNTACGCN GGCACNGAGTT | Homology cloning primer |
| GLB1-de-R | ATGTCCTCCATGCCNCCCTCCAT | Homology cloning primer |
| GSP1 | GATTACGCCAAGCTTACTATGTCCACCTGGGAGCGTGAC | 3’-RACE primer |
| GSP2 | GATTACGCCAAGCTTCGCTCCCAGGTGGACATAGTGAC | 5’-RACE primer |
| GLB1-F | ATGATTGGCACAGTGCTGCAGAA | Gene cloning primer |
| GLB1-R | CTAACCACTAGCCTCCGGCTTG | Gene cloning primer |
| GLB1-B-F | GGAATTCATGATTGGCACAGTGCTGCAGAA | Gene cloning primer |
| GLB1- B-R | CCCAAGCTTCTAACCACTAGCCTCCGGCTTG | Gene cloning primer |
| GLB1-Q-F | CGCCTGGCATTTGTCATTG | Real-time gene primer |
| GLB1-Q-R | AAACTCAGTGCCAGCATAGCG | Real-time gene primer |
| 18S-F | CAAAGCAAGCCTACGCTCT | Real-time gene primer |
| 18S-R | ATACGAATGCCCCCGACT | Real-time gene primer |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, R.; Li, Y.; Lu, Y. Sequencing and Characterization of Novel PII Signaling Protein Gene in Microalga Haematococcus pluvialis. Mar. Drugs 2017, 15, 304. https://doi.org/10.3390/md15100304
Ma R, Li Y, Lu Y. Sequencing and Characterization of Novel PII Signaling Protein Gene in Microalga Haematococcus pluvialis. Marine Drugs. 2017; 15(10):304. https://doi.org/10.3390/md15100304
Chicago/Turabian StyleMa, Ruijuan, Yan Li, and Yinghua Lu. 2017. "Sequencing and Characterization of Novel PII Signaling Protein Gene in Microalga Haematococcus pluvialis" Marine Drugs 15, no. 10: 304. https://doi.org/10.3390/md15100304
APA StyleMa, R., Li, Y., & Lu, Y. (2017). Sequencing and Characterization of Novel PII Signaling Protein Gene in Microalga Haematococcus pluvialis. Marine Drugs, 15(10), 304. https://doi.org/10.3390/md15100304