Optimization of Collagenase Production by Pseudoalteromonas sp. SJN2 and Application of Collagenases in the Preparation of Antioxidative Hydrolysates

Abstract
:1. Introduction
2. Results
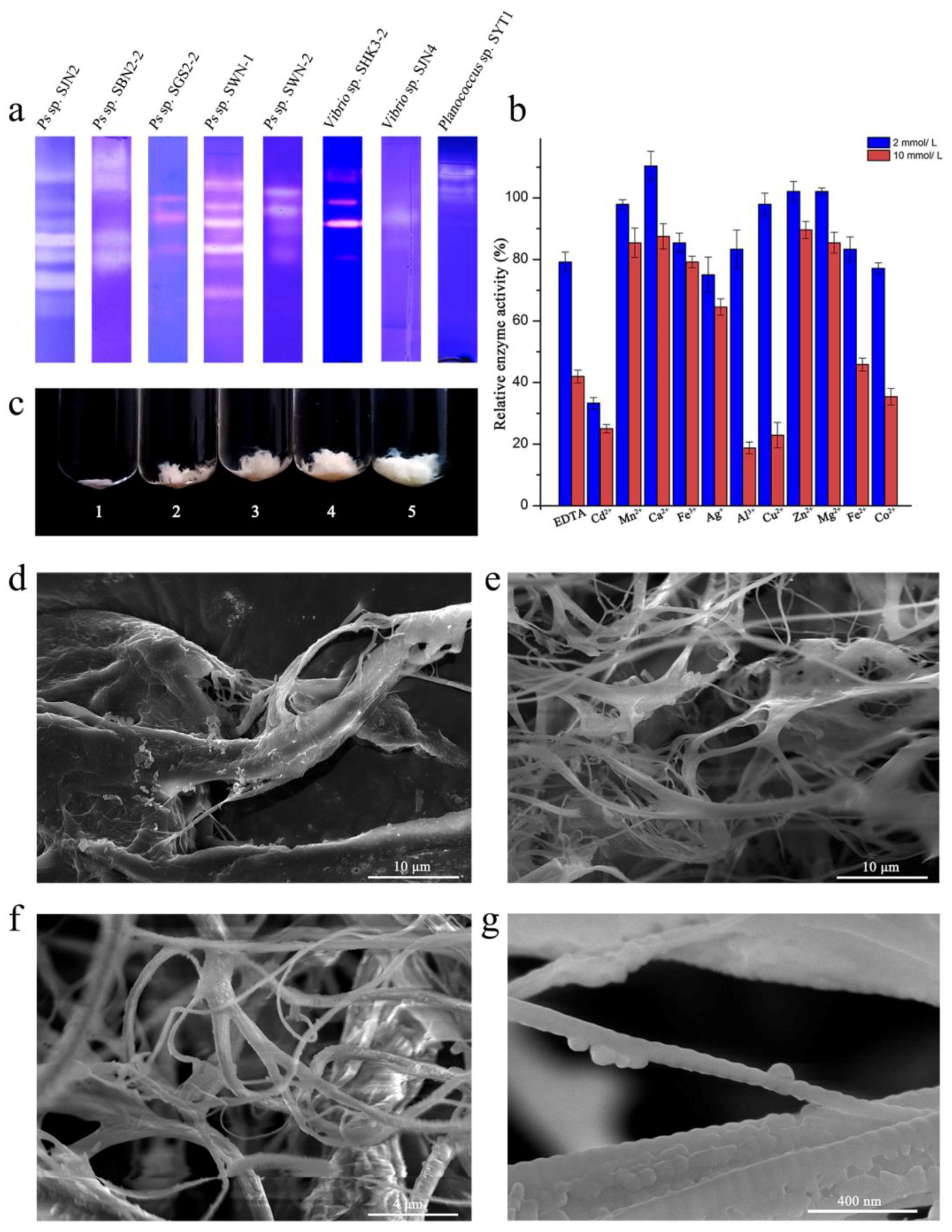
2.1. Purification and Enzymatic Properties of Ps sp. SJN2 Collagenases
2.2. Catalytic Efficiency of Collagenases from Ps sp. SJN2
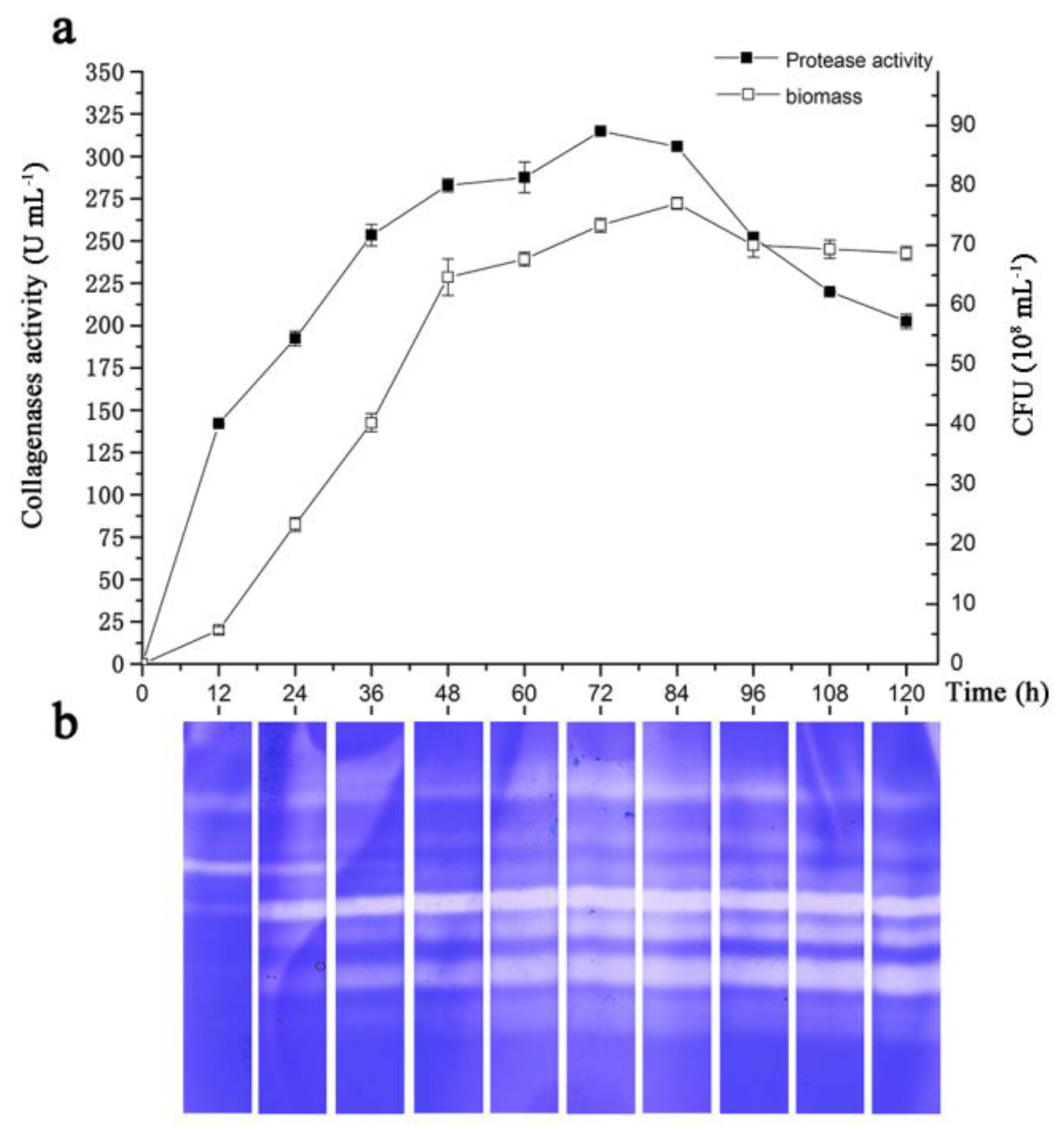
2.3. Initial Screening of Significant Fermentation Conditions
2.4. Further Optimization by Response Surface Methodology
− 24.74A2 − 25.18B2 − 48.46C2
2.5. Experimental Validation of the Model
2.6. Antioxidant Activity of the Hydrolysates
3. Materials and Methods
3.1. Microorganism
3.2. Fermentation
3.3. Assay of Collagenase Activity and Substrate Immersing Zymography
3.4. Screening of Significant Variables by Using Plackett–Burman Designs
3.5. Investigation of the Shifts in the Trends of the Significant Variables Using the Path of Steepest Ascent
3.6. Optimization by Response Surface Methodology
3.7. Statistical Analysis
3.8. Validation of the Optimization Model
3.9. Purification of Ps sp. SJN2 Collagenases
3.10. Preparation of Collagen from Fishery By-Products
3.11. Enzymatic Hydrolysis and Properties of Ps sp. SJN2 Collagenase
3.12. DPPH Radical Scavenging and ORAC Assay
4. Conclusions
Supplementary Materials
Acknowledgments
Conflicts of interest
References
- Zhao, G.Y.; Chen, X.L.; Zhao, H.L.; Xie, B.B.; Zhou, B.C.; Zhang, Y.Z. Hydrolysis of insoluble collagen by deseasin mcp-01 from deep-sea Pseudoalteromonas sp. Sm9913: Collagenolytic characters, collagen-binding ability of c-terminal polycystic kidney disease domain, and implication for its novel role in deep-sea sedimentary particulate organic nitrogen degradation. J. Biol. Chem. 2008, 283, 36100–36107. [Google Scholar] [PubMed]
- Silva, T.H.; Moreira-Silva, J.; Marques, A.L.; Domingues, A.; Bayon, Y.; Reis, R.L. Marine origin collagens and its potential applications. Mar. Drugs 2014, 12, 5881–5901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.; Vazquez, J.A.; Perez-Martin, R.I.; Sotelo, C.G. Hydrolysates of fish skin collagen: An opportunity for valorizing fish industry byproducts. Mar. Drugs 2017, 15, 131. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K. Collagenolytic proteases from bacteria. Appl. Microbiol. Biotechnol. 2004, 63, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.S.; Correia, A.; Esteves, A.C. Bacterial collagenases—A review. Crit. Rev. Microbiol. 2016, 42, 106–126. [Google Scholar] [CrossRef] [PubMed]
- Shekhter, A.B.; Balakireva, A.V.; Kuznetsova, N.V.; Vukolova, M.N.; Litvitsky, P.F.; Zamyatnin, A.A. Collagenolytic enzymes and their applications in biomedicine. Curr. Med. Chem. 2017, 24, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ran, L.Y.; Su, H.N.; Zhao, G.Y.; Gao, X.; Zhou, M.Y.; Wang, P.; Zhao, H.L.; Xie, B.B.; Zhang, X.Y.; Chen, X.L.; et al. Structural and mechanistic insights into collagen degradation by a bacterial collagenolytic serine protease in the subtilisin family. Mol. Microbiol. 2013, 90, 997–1010. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Y.; Zhou, M.Y.; Zhao, H.L.; Chen, X.L.; Xie, B.B.; Zhang, X.Y.; He, H.L.; Zhou, B.C.; Zhang, Y.Z. Tenderization effect of cold-adapted collagenolytic protease mcp-01 on beef meat at low temperature and its mechanism. Food Chem. 2012, 134, 1738–1744. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, O.; Koide, T.; Kobayashi, R.; Nagata, K.; Okabe, A. Substrate recognition by the collagen-binding domain of clostridium histolyticum class I collagenase. J. Biol. Chem. 2001, 276, 8761–8770. [Google Scholar] [CrossRef] [PubMed]
- Bassetto, F.; Maschio, N.; Abatangelo, G.; Zavan, B.; Scarpa, C.; Vindigni, V. Collagenase from Vibrio alginolyticus cultures: Experimental study and clinical perspectives. Surg. Innov. 2016, 23, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Vijayaraghavan, P.; Vincent, S.G. Statistical optimization of fibrinolytic enzyme production by Pseudoalteromonas sp. Ind11 using cow dung substrate by response surface methodology. SpringerPlus 2014, 3, 60. [Google Scholar] [CrossRef] [PubMed]
- Haddar, A.; Fakhfakh-Zouari, N.; Hmidet, N.; Frikha, F.; Nasri, M.; Kamoun, A.S. Low-cost fermentation medium for alkaline protease production by Bacillus Mojavensis a21 using hulled grain of wheat and sardinella peptone. J. Biosci. Bioeng. 2010, 110, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.V.; Wee, Y.J.; Yun, J.S.; Ryu, H.W. Optimization of alkaline protease production by batch culture of Bacillus sp. Rky3 through plackett-burman and response surface methodological approaches. Bioresour. Technol. 2008, 99, 2242–2249. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Long, L.; Sun, X.; Wu, H.; Li, T.; Xiang, W. Optimization of medium using response surface methodology for lipid production by Scenedesmus sp. Mar. Drugs 2014, 12, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Beg, Q.K.; Lorenz, P. Bacterial alkaline proteases: Molecular approaches and industrial applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32. [Google Scholar] [PubMed]
- He, H.L.; Liu, D.; Ma, C.B. Review on the angiotensin-i-converting enzyme (ace) inhibitor peptides from marine proteins. Appl. Biochem. Biotechnol. 2013, 169, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.L.; Kim, J.S.; Wang, J.K. Optimization for the maximum bacteriocin production of Lactobacillus brevis df01 using response surface methodology. Food Sci. Biotechnol. 2012, 21, 653–659. [Google Scholar]
- Aleman, A.; Gimenez, B.; Montero, P.; Gomez-Guillen, M.C. Antioxidant activity of several marine skin gelatins. Lwt-Food Sci. Technol. 2011, 44, 407–413. [Google Scholar] [CrossRef]
- Liu, D.; Yang, X.; Huang, J.; Wu, R.; Wu, C.; He, H.; Li, H. In situ demonstration and characteristic analysis of the protease components from marine bacteria using substrate immersing zymography. Appl. Biochem. Biotechnol. 2015, 175, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Kleijnen, J.P.C.; Hertog, D.D.; Angün, E. Response surface methodology’s steepest ascent and step size revisited: Correction. Eur. J. Oper. Res. 2002, 170, 664–666. [Google Scholar] [CrossRef]
- Zhao, H.L.; Yang, J.; Chen, X.L.; Su, H.N.; Zhang, X.Y.; Huang, F.; Zhou, B.C.; Xie, B.B. Optimization of fermentation conditions for the production of the M23 protease pseudoalterin by deep-sea Pseudoalteromonas sp. CF6-2 with artery powder as an inducer. Molecules 2014, 19, 4779–4790. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Fang, Y.; Lv, M.; Wang, S.; Chen, L. Optimization of the production of organic solvent-stable protease by bacillus sphaericus ds11 with response surface methodology. Bioresour. Technol. 2010, 101, 7924–7929. [Google Scholar] [CrossRef] [PubMed]
- Sahan, T.; Ceylan, H.; Sahiner, N.; Aktas, N. Optimization of removal conditions of copper ions from aqueous solutions by Trametes versicolor. Bioresour. Technol. 2010, 101, 4520–4526. [Google Scholar] [CrossRef] [PubMed]
- Oskouiea, S.F.G.; Tabandehb, F.; Yakhchalib, B.; Eftekhara, F. Response surface optimization of medium composition for alkaline protease production by Bacillus clausii. Biochem. Eng. J. 2008, 39, 37–42. [Google Scholar] [CrossRef]
- Ogawa, M. Biochemical properties of bone and scale collagens isolated from the subtropical fish black drum (Pogonia cromis) and sheepshead seabream (Archosargus probatocephalus). Food Chem. 2004, 88, 495–501. [Google Scholar] [CrossRef]
- Vazquez, J.A.; Blanco, M.; Massa, A.E.; Amado, I.R.; Perez-Martin, R.I. Production of fish protein hydrolysates from Scyliorhinus canicula discards with antihypertensive and antioxidant activities by enzymatic hydrolysis and mathematical optimization using response surface methodology. Mar. Drugs 2017, 15, 306. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Chen, L.; Liu, D.; Huang, J.; Zhang, J.; Xiao, X.; Lei, M.; Chen, Y.; He, H. Preparation of antioxidant peptides from salmon byproducts with bacterial extracellular proteases. Mar. Drugs 2017, 15, 4. [Google Scholar] [CrossRef] [PubMed]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Dávalos, A.; Carmen Gómezcordovés, A.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (orac−fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Component | Unit | Lower Level (−1) | Higher Level (+1) |
|---|---|---|---|---|
| X1 | Temperature | °C | 18.0 | 27.0 |
| X2 | pH | - | 7.5 | 8.5 |
| X3 | Inoculum | μL | 500.0 | 750.0 |
| X4 | Culture time | d | 4.0 | 6.0 |
| X5 | Corn meal | g·L−1 | 20.0 | 30.0 |
| X6 | Bran liquid | mL·L−1 | 10.0 | 15.0 |
| X7 | Soybean powder | g·L−1 | 20.0 | 30.0 |
| X8 | Dummy variable | - | - | - |
| X9 | Dummy variable | - | - | - |
| X10 | Dummy variable | - | - | - |
| X11 | Dummy variable | - | - | - |
| Run | Variables | Collagenase Activity U·mL−1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | ||
| 1 | −1 | +1 | −1 | +1 | +1 | −1 | +1 | +1 | +1 | −1 | −1 | 230.53 |
| 2 | −1 | −1 | −1 | +1 | −1 | +1 | +1 | −1 | +1 | +1 | +1 | 235.50 |
| 3 | −1 | +1 | +1 | −1 | +1 | +1 | +1 | −1 | −1 | −1 | +1 | 273.33 |
| 4 | +1 | −1 | +1 | +1 | −1 | +1 | +1 | +1 | −1 | −1 | −1 | 162.23 |
| 5 | +1 | +1 | +1 | −1 | −1 | −1 | +1 | −1 | +1 | +1 | −1 | 168.39 |
| 6 | +1 | −1 | +1 | +1 | +1 | −1 | −1 | −1 | +1 | −1 | +1 | 127.82 |
| 7 | +1 | −1 | −1 | −1 | +1 | −1 | +1 | +1 | −1 | +1 | +1 | 225.40 |
| 8 | +1 | +1 | −1 | −1 | −1 | +1 | −1 | +1 | +1 | −1 | +1 | 202.29 |
| 9 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 215.47 |
| 10 | −1 | −1 | +1 | −1 | +1 | +1 | −1 | +1 | +1 | +1 | −1 | 218.72 |
| 11 | +1 | +1 | −1 | +1 | +1 | +1 | −1 | −1 | −1 | +1 | −1 | 147.17 |
| 12 | −1 | +1 | +1 | +1 | −1 | −1 | −1 | +1 | −1 | +1 | +1 | 173.70 |
| Source | df | Collagenase Activity of Fermented Broth a | ||||
|---|---|---|---|---|---|---|
| Sum of Squares | Coefficient Estimate | Mean Square | F-Value | p-value Probability > F | ||
| Model a | 7 | 18,796.53 | - | 2685.22 | 9.770 | 0.0219 b |
| X1 | 1 | 8213.77 | −26.16 | 8213.77 | 29.880 | 0.0054 b |
| X2 | 1 | 8.79 | 0.86 | 8.79 | 0.032 | 0.8668 |
| X3 | 1 | 1455.41 | −11.01 | 1455.41 | 5.290 | 0.0829 |
| X4 | 1 | 4280.74 | −18.89 | 4280.74 | 15.570 | 0.0169 b |
| X5 | 1 | 356.35 | 5.45 | 356.35 | 1.300 | 0.3184 |
| X6 | 1 | 799.01 | 8.16 | 799.01 | 2.910 | 0.1634 |
| X7 | 1 | 3682.46 | 17.52 | 3682.46 | 13.400 | 0.0216 b |
| Residual | 4 | 1099.59 | - | 274.90 | - | - |
| Corrected Total | 11 | 19,896.12 | - | - | - | - |
| Item | Temperature (°C) | Culture time (d) | Soybean Concentration (g·L−1) | Collagenase Activity (U·mL−1) |
|---|---|---|---|---|
| Slope a | −26.16 | −18.89 | 17.52 | - |
| Base b | 18.00 | 4.00 | 30.00 | - |
| Average c | 22.50 | 5.00 | 25.00 | - |
| Ratio d | 0.017 | 0.034 | 0.034 | - |
| △ e | −2.00 | −0.64 | 3.00 | - |
| Base | 18.00 | 4.00 | 30.00 | 224.66 ± 11.53 |
| Base+△ | 16.00 | 3.36 | 33.00 | 279.18 ± 8.71 |
| Base+2△ | 14.00 | 2.72 | 36.00 | 206.22 ± 26.47 |
| Base+3△ | 12.00 | 2.08 | 39.00 | 195.09 ± 9.65 |
| Variables | Component | Unit | +1.68 Level | +1 Level | 0 Level | −1 Level | −1.68 Level |
|---|---|---|---|---|---|---|---|
| A | Temperature | °C | 19.36 | 18.00 | 16.00 | 14.00 | 12.64 |
| B | Culture time | d | 4.44 | 4.00 | 3.36 | 2.72 | 2.28 |
| C | Soybean powder | g·L−1 | 38.04 | 36.00 | 33.00 | 30.00 | 27.96 |
| Run | Variables | Collagenase Activity (U·mL−1) | ||
|---|---|---|---|---|
| A | B | C | ||
| 1 | +1.68 | 0 | 0 | 251.13 |
| 2 | +1 | −1 | +1 | 221.71 |
| 3 | 0 | −1.68 | 0 | 187.44 |
| 4 | +1 | −1 | −1 | 160.45 |
| 5 | −1 | +1 | −1 | 123.73 |
| 6 | 0 | 0 | 0 | 278.35 |
| 7 | 0 | 0 | 0 | 282.97 |
| 8 | +1 | +1 | −1 | 197.21 |
| 9 | +1 | +1 | +1 | 302.56 |
| 10 | 0 | +1.68 | 0 | 243.74 |
| 11 | −1.68 | 0 | 0 | 182.53 |
| 12 | 0 | 0 | 0 | 278.07 |
| 13 | 0 | 0 | 0 | 287.85 |
| 14 | 0 | 0 | −1.68 | 107.18 |
| 15 | −1 | +1 | +1 | 214.38 |
| 16 | 0 | 0 | +1.68 | 192.30 |
| 17 | −1 | −1 | +1 | 182.51 |
| 18 | −1 | −1 | −1 | 119.22 |
| 19 | 0 | 0 | 0 | 280.56 |
| 20 | 0 | 0 | 0 | 319.77 |
| Source | df | Collagenase Activity of Fermented Brotha a | |||
|---|---|---|---|---|---|
| Sum of Squares | Mean Square | F-Value | p-value Probability > F | ||
| Model | 9 | 75,803.27 | 8422.59 | 35.19 | 0.0001 b |
| A | 1 | 9356.35 | 9356.35 | 39.09 | 0.0001 b |
| B | 1 | 4528.07 | 4528.07 | 18.92 | 0.0014 b |
| C | 1 | 15,744.59 | 15,744.59 | 65.78 | 0.0001 b |
| AB | 1 | 824.79 | 824.79 | 3.45 | 0.0931 |
| AC | 1 | 20.07 | 20.07 | 0.084 | 0.7781 |
| BC | 1 | 638.14 | 638.14 | 2.67 | 0.1335 |
| A2 | 1 | 8823.57 | 8823.57 | 36.87 | 0.0001 b |
| B2 | 1 | 9139.01 | 9139.01 | 38.18 | 0.0001 b |
| C2 | 1 | 33,848.58 | 33,848.58 | 141.43 | 0.0001 b |
| Residual | 10 | 2393.38 | 239.34 | - | - |
| Lack of Fit | 5 | 1111.68 | 222.34 | 0.87 | 0.5601 |
| Pure Error | 5 | 1281.71 | 256.34 | - | - |
| Corrected Total | 19 | 78,196.66 | - | - | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Xiao, X.; Liu, D.; Wu, R.; Wu, C.; Zhang, J.; Huang, J.; Liao, B.; He, H. Optimization of Collagenase Production by Pseudoalteromonas sp. SJN2 and Application of Collagenases in the Preparation of Antioxidative Hydrolysates. Mar. Drugs 2017, 15, 377. https://doi.org/10.3390/md15120377
Yang X, Xiao X, Liu D, Wu R, Wu C, Zhang J, Huang J, Liao B, He H. Optimization of Collagenase Production by Pseudoalteromonas sp. SJN2 and Application of Collagenases in the Preparation of Antioxidative Hydrolysates. Marine Drugs. 2017; 15(12):377. https://doi.org/10.3390/md15120377
Chicago/Turabian StyleYang, Xinghao, Xiao Xiao, Dan Liu, Ribang Wu, Cuiling Wu, Jiang Zhang, Jiafeng Huang, Binqiang Liao, and Hailun He. 2017. "Optimization of Collagenase Production by Pseudoalteromonas sp. SJN2 and Application of Collagenases in the Preparation of Antioxidative Hydrolysates" Marine Drugs 15, no. 12: 377. https://doi.org/10.3390/md15120377
APA StyleYang, X., Xiao, X., Liu, D., Wu, R., Wu, C., Zhang, J., Huang, J., Liao, B., & He, H. (2017). Optimization of Collagenase Production by Pseudoalteromonas sp. SJN2 and Application of Collagenases in the Preparation of Antioxidative Hydrolysates. Marine Drugs, 15(12), 377. https://doi.org/10.3390/md15120377