
Bioactive Peptide of Marine Origin for the Prevention and Treatment of Non-Communicable Diseases


Abstract
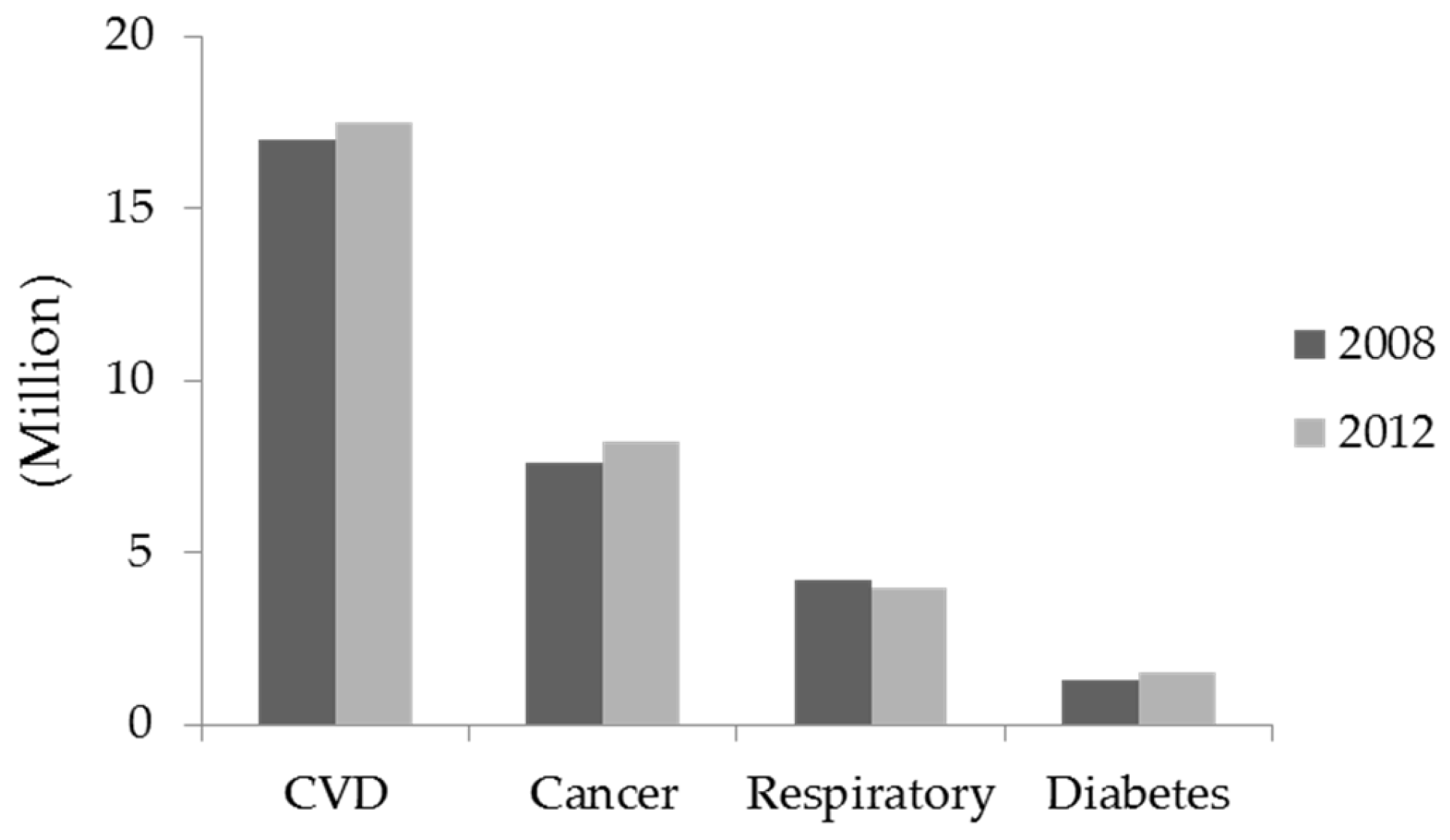
:1. Introduction
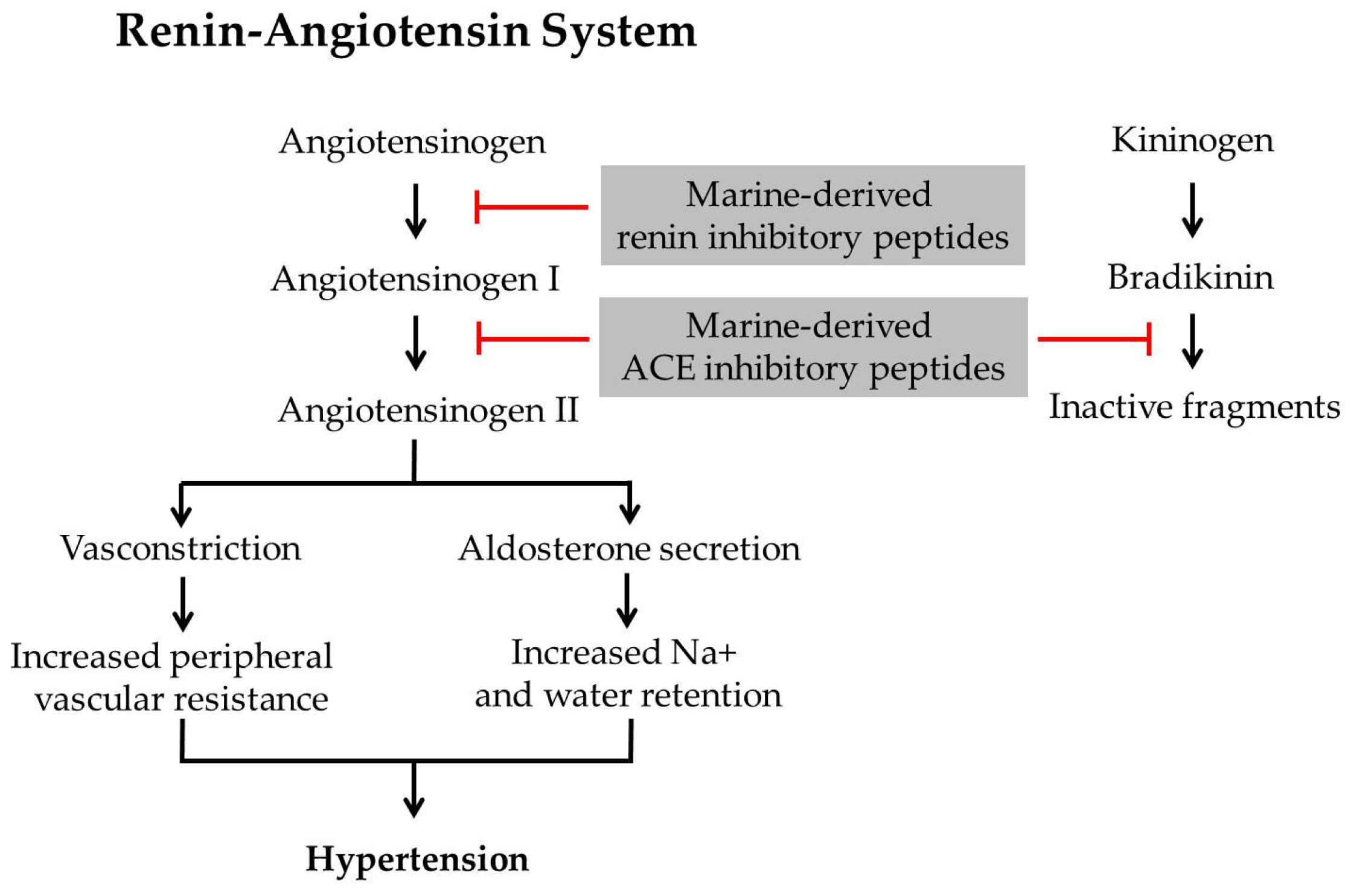
2. Marine-Derived Cardio Protective Peptides
2.1. Marine-Derived Renin Inhibitory Peptides
2.2. Marine-Derived ACE Inhibitory Peptides
3. Marine-Derived Anti-Cancer Peptides
3.1. Anti-Cancer Peptides Derived from Sponges
3.2. Anti-Cancer Peptides Derived from Fish
3.3. Anti-Cancer Peptides Derived from Urochordata
3.4. Anti-Cancer Peptides Derived from Mollusks
3.5. Anti-Cancer Peptides Derived from Cyanobacteria
4. Marine-Derived Antioxidant Peptides
5. Anti-Diabetic and Hypocholesterolemic Effects of Marine-Derived Bioactive Peptides
6. Future Perspectives of Marine-Derived Bioactive Peptides
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACE | Angiotensin converting enzymes |
| CVD | Cardiovascular disease |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| FAO | Food and Agriculture Organization |
| FOSHU | foods for specified health uses |
| IC50 | half maximal inhibitory concentration |
| IL-6 | interleukin-6 |
| IL-1β | interleukin-1β |
| MCP | marine collagen peptide |
| MW | Molecular weight |
| NCD | Non communicable diseases |
| PUFA | polyunsaturated fatty acids |
| RAS | renin-angiotensin system |
| ROS | reactive oxygen species |
| SHR | spontaneously hypertensive rat |
| SBP | systolic blood pressure |
| TNF-α | tumor necrosis factor-α |
| WHO | World Health Organization |
References
- World Health Organization. Global Status Report on Noncommuniacble Diseases; World Health Organization: Genève, Switzerland, 2014; p. 280. [Google Scholar]
- World Health Organization. Global Status Report on Noncommunicable Diseases; World Health Organization: Genève, Switzerland, 2010; p. 162. [Google Scholar]
- Bhandari, G.P.; Angdembe, M.R.; Dhimal, M.; Neupane, S.; Bhusal, C. State of non-communicable diseases in Nepal. BMC Public Health 2014, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. Looking Beyond the Terrestrial: The Potential of Seaweed Derived Bioactives to Treat Non-Communicable Diseases. Mar. Drugs 2016, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Wagner, K.-H.; Brath, H. A global view on the development of non communicable diseases. Prev. Med. 2012, 54, S38–S41. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, K.; Cheung, B.W.Y.; Schröder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Hecker, K.D.; Bonanome, A.; Coval, S.M.; Binkoski, A.E.; Hilpert, K.F.; Griel, A.E.; Etherton, T.D. Bioactive compounds in foods: Their role in the prevention of cardiovascular disease and cancer. Am. J. Med. 2002, 113 (Suppl. 9), 71S–88S. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2010, 84, 14–21. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.-K. Biological activities and health benefit effects of natural pigments derived from marine algae. J. Funct. Foods 2011, 3, 255–266. [Google Scholar] [CrossRef]
- Yusuf, S.; Wood, D.; Ralston, J.; Reddy, K.S. The World Heart Federation’s vision for worldwide cardiovascular disease prevention. Lancet 2015, 386, 399–402. [Google Scholar] [CrossRef]
- Nichols, M.; Townsend, N.; Scarborough, P.; Rayner, M. Cardiovascular disease in Europe 2014: Epidemiological update. Eur. Heart J. 2014, 35, 2950–2959. [Google Scholar] [CrossRef] [PubMed]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Himaya, S.; Ngo, D.-H.; Ryu, B.; Kim, S.-K. An active peptide purified from gastrointestinal enzyme hydrolysate of Pacific cod skin gelatin attenuates angiotensin-1 converting enzyme (ACE) activity and cellular oxidative stress. Food Chem. 2012, 132, 1872–1882. [Google Scholar] [CrossRef]
- Kher, V. Renin inhibition—Benefit beyond hypertension control. J. Assoc. Phys. India 2009, 57, 518–521. [Google Scholar]
- Allikmets, K. Aliskiren—An orally active renin inhibitor. Review of pharmacology, pharmacodynamics, kinetics, and clinical potential in the treatment of hypertension. Vasc. Health Risk Manag. 2007, 3, 809. [Google Scholar] [PubMed]
- Fitzgerald, C.N.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Aluko, R.E.; Hossain, M.; Rai, D.K.; Hayes, M. Potential of a renin inhibitory peptide from the red seaweed Palmaria palmata as a functional food ingredient following confirmation and characterization of a hypotensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2014, 62, 8352–8356. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Maekawa, K.; Chen, J.-R. Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J. Nutr. Biochem. 2004, 15, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, C.; Gallagher, E.; Doran, L.; Auty, M.; Prieto, J.; Hayes, M. Increasing the health benefits of bread: Assessment of the physical and sensory qualities of bread formulated using a renin inhibitory Palmaria palmata protein hydrolysate. LWT Food Sci. Technol. 2014, 56, 398–405. [Google Scholar] [CrossRef]
- Suetsuna, K.; Osajika, K. The inhibitroy activity of angiotensin-1 converting enzyme of basic peptides from sardine and hair tail meat. Bull. Jpn. Soc. Sci. Fish. 1986, 52, 1981–1984. [Google Scholar] [CrossRef]
- Kim, S.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Wijesekara, I.; Kim, S.K. Angiotensin-I-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and Characterization of Angiotensin I-Converting Enzyme Inhibitors from Sour Milk. J. Dairy Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Ariyoshi, Y. Angiotensin-converting enzyme inhibitors derived from food proteins. Trends Food Sci. Technol. 1993, 4, 139–144. [Google Scholar] [CrossRef]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Nomura, A.; Noda, N.; Maruyama, S. Purification of angiotensin I-converting enzyme inhibitors in pelagic thresher Alopias pelagicus muscle hydrolysate and viscera extracts. Fish. Sci. 2002, 68, 954–956. [Google Scholar] [CrossRef]
- Ikeda, A.; Ichino, H.; Kiguchiya, S.; Chigwechokha, P.; Komatsu, M.; Shiozaki, K. Evaluation and Identification of Potent Angiotensin-I Converting Enzyme Inhibitory Peptide Derived from Dwarf Gulper Shark (Centrophorus atromarginatus). J. Food Process. Preserv. 2015, 39, 107–115. [Google Scholar] [CrossRef]
- Wu, H.; He, H.-L.; Chen, X.-L.; Sun, C.-Y.; Zhang, Y.-Z.; Zhou, B.-C. Purification and identification of novel angiotensin-I-converting enzyme inhibitory peptides from shark meat hydrolysate. Proc. Biochem. 2008, 43, 457–461. [Google Scholar] [CrossRef]
- Jian, P.W.; Aluko, R.E.; Nakai, S. Structural Requirements of Angiotensin I-Converting Enzyme Inhibitory Peptides: Quantitative Structure-Activity Relationship Study of Di- and Tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar]
- Sato, M.; Hosokawa, T.; Yamaguchi, T.; Nakano, T.; Muramoto, K.; Kahara, T.; Funayama, K.; Kobayashi, A.; Nakano, T. Angiotensin I-converting enzyme inhibitory peptides derived from wakame (Undaria pinnatifida) and their antihypertensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2002, 50, 6245–6252. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K. Purification and identification of angiotensin I-converting enzyme inhibitors from the red alga Porphyra yezoensis. J. Mar. Biotechnol. 1998, 6, 163–167. [Google Scholar] [PubMed]
- Suetsuna, K.; Chen, J.-R. Identification of Antihypertensive Peptides from Peptic Digest of Two Microalgae, Chlorella vulgaris and Spirulina platensis. Mar. Biotechnol. 2001, 3, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Kohama, Y.; Matsumoto, S.; Oka, H.; Teramoto, T.; Okabe, M.; Mimura, T. Isolation of angiotensin-converting enzyme inhibitor from tuna muscle. Biochem. Biophys. Res. Commun. 1988, 155, 332–337. [Google Scholar] [CrossRef]
- Matsumura, N.; Fujii, M.; Takeda, Y.; Sugita, K.; Shimizu, T. Angiotensin I-converting enzyme inhibitory peptides derived from bonito bowels autolysate. Biosci. Biotechnol. Biochem. 1993, 57, 695–697. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Kim, Y.-T.; Byun, H.-G.; Nam, K.-S.; Joo, D.-S.; Shahidi, F. Isolation and characterization of antioxidative peptides from gelatin hydrolysate of Alaska pollack skin. J. Agric. Food Chem. 2001, 49, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Ono, S.; Hosokawa, M.; Miyashita, K.; Takahashi, K. Isolation of Peptides with Angiotensin I-converting Enzyme Inhibitory Effect Derived from Hydrolysate of Upstream Chum Salmon Muscle. J. Food Sci. 2003, 68, 1611–1614. [Google Scholar] [CrossRef]
- Enari, H.; Takahashi, Y.; Kawarasaki, M.; Tada, M.; Tatsuta, K. Identification of angiotensin I-converting enzyme inhibitory peptides derived from salmon muscle and their antihypertensive effect. Fish. Sci. 2008, 74, 911–920. [Google Scholar] [CrossRef]
- Ngo, D.-H.; Kang, K.-H.; Ryu, B.; Vo, T.-S.; Jung, W.-K.; Byun, H.-G.; Kim, S.-K. Angiotensin-I converting enzyme inhibitory peptides from antihypertensive skate (Okamejei kenojei) skin gelatin hydrolysate in spontaneously hypertensive rats. Food Chem. 2015, 174, 37–43. [Google Scholar] [CrossRef] [PubMed]
- García-Moreno, P.J.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Production and identification of angiotensin I-converting enzyme (ACE) inhibitory peptides from Mediterranean fish discards. J. Funct. Foods 2015, 18, 95–105. [Google Scholar] [CrossRef]
- He, H.L.; Chen, X.L.; Sun, C.Y.; Zhang, Y.Z.; Zhou, B.C. Analysis of novel angiotensin 1 converting enzyme inhibitory peptides from protease-hydrolyzed marine shrimp Acetes chinensis. J. Pept. Sci. 2006, 12, 726–733. [Google Scholar]
- Wang, Y.-K.; He, H.-L.; Chen, X.-L.; Sun, C.-Y.; Zhang, Y.-Z.; Zhou, B.-C. Production of novel angiotensin I-converting enzyme inhibitory peptides by fermentation of marine shrimp Acetes chinensis with Lactobacillus fermentum SM 605. Appl. Microbiol. Biotechnol. 2008, 79, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Zhang, C.; Hong, P.; Ji, H.; Hao, J. Purification and identification of an ACE inhibitory peptide from the peptic hydrolysate of Acetes chinensis and its antihypertensive effects in spontaneously hypertensive rats. Int. J. Food Sci. Technol. 2010, 45, 959–965. [Google Scholar] [CrossRef]
- Nii, Y.; Fukuta, K.; Yoshimoto, R.; Sakai, K.; Ogawa, T. Determination of antihypertensive peptides from an izumi shrimp hydrolysate. Biosci. Biotechnol. Biochem. 2008, 72, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Jia, A.; Zhang, Y.; Zhu, H.; Zhang, C.; Sun, Z.; Liu, C. Purification and characterization of angiotensin I converting enzyme inhibitory peptides from jellyfish Rhopilema esculentum. Food Res. Int. 2013, 50, 339–343. [Google Scholar] [CrossRef]
- Du, L.; Fang, M.; Wu, H.; Xie, J.; Wu, Y.; Li, P.; Zhang, D.; Huang, Z.; Xia, Y.; Zhou, L. A novel angiotensin I-converting enzyme inhibitory peptide from Phascolosoma esculenta water-soluble protein hydrolysate. J. Funct. Foods 2013, 5, 475–483. [Google Scholar] [CrossRef]
- Suetsuna, K. Identification of antihypertensive peptides from peptic digest of the short-necked clam Tapes philippinarum and the pearl oyster Pinctada fucata martensii. Fish. Sci. 2002, 68, 233–235. [Google Scholar] [CrossRef]
- Fujita, H.; Yamagami, T.; Ohshima, K. Effects of an ACE-inhibitory agent, katsuobushi oligopeptide, in the spontaneously hypertensive rat and in borderline and mildly hypertensive subjects. Nutr. Res. 2001, 21, 1149–1158. [Google Scholar] [CrossRef]
- Fujita, H.; Yoshikawa, M. LKPNM: A prodrug-type ACE-inhibitory peptide derived from fish protein. Immunopharmacology 1999, 44, 123–127. [Google Scholar] [CrossRef]
- Kouno, K.; Hirano, S.-I.; Kuboki, H.; Kasai, M.; Hatae, K. Effects of Dried Bonito (Katsuobushi) and Captopril, an Angiotensin I-Converting Enzyme Inhibitor, on Rat Isolated Aorta: A Possible Mechanism of Antihypertensive Action. Biosci. Biotechnol. Biochem. 2005, 69, 911–915. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Jeon, J.-K.; Byun, H.-G. Antihypertensive effect of novel angiotensin I converting enzyme inhibitory peptide from chum salmon (Oncorhynchus keta) skin in spontaneously hypertensive rats. J. Funct. Foods 2014, 7, 381–389. [Google Scholar] [CrossRef]
- Lee, S.-H.; Qian, Z.-J.; Kim, S.-K. A novel angiotensin I converting enzyme inhibitory peptide from tuna frame protein hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Food Chem. 2010, 118, 96–102. [Google Scholar] [CrossRef]
- Ko, S.-C.; Kang, N.; Kim, E.-A.; Kang, M.C.; Lee, S.-H.; Kang, S.-M.; Lee, J.-B.; Jeon, B.-T.; Kim, S.-K.; Park, S.-J. A novel angiotensin I-converting enzyme (ACE) inhibitory peptide from a marine Chlorella ellipsoidea and its antihypertensive effect in spontaneously hypertensive rats. Proc. Biochem. 2012, 47, 2005–2011. [Google Scholar] [CrossRef]
- Roy, M.; Mukherjee, A.; Sarkar, R.; Mukherjee, S.; Biswas, J. In search of natural remediation for cervical cancer. Anti-Cancer Agents Med. Chem. 2015, 15, 57–65. [Google Scholar] [CrossRef]
- He, G.; Karin, M. NF-κB and STAT3—Key players in liver inflammation and cancer. Cell Res. 2011, 21, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.; Cameron, D.; Waters, R.; Barrett-Lee, P.; Ellis, P.; Russell, S.; Bliss, J.; Hopwood, P.; Investigators, T.T. Comparison of patient reported quality of life and impact of treatment side effects experienced with a taxane-containing regimen and standard anthracycline based chemotherapy for early breast cancer: 6 year results from the UK TACT trial (CRUK/01/001). Eur. J. Cancer 2014, 50, 2375–2389. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Kalimuthu, S. Introduction to Anticancer Drugs from Marine Origin. In Handbook of Anticancer Drugs from Marine Origin; Springer: Berlin, Germany, 2015; pp. 1–13. [Google Scholar]
- Zheng, L.-H.; Wang, Y.-J.; Sheng, J.; Wang, F.; Zheng, Y.; Lin, X.-K.; Sun, M. Antitumor peptides from marine organisms. Mar. Drugs 2011, 9, 1840–1859. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H. Marine peptides: Bioactivities and applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Crews, P.; Manes, L.V.; Boehler, M. Jasplakinolide, a cyclodepsipeptide from the marine sponge, Jaspis sp. Tetrahedron Lett. 1986, 27, 2797–2800. [Google Scholar] [CrossRef]
- Takeuchi, H.; Ara, G.; Sausville, A.E.; Teicher, B. Jasplakinolide: Interaction with radiation and hyperthermia in human prostate carcinoma and Lewis lung carcinoma. Cancer Chemother. Pharmacol. 1998, 42, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.J.; Morinaka, B.I.; Amagata, T.; Tenney, K.; Bray, W.M.; Gassner, N.C.; Lokey, R.S.; Crews, P. New Structures and Bioactivity Properties of Jasplakinolide (Jaspamide) Analogues from Marine Sponges. J. Med. Chem. 2010, 53, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.; Prado, M.P.; Konno, K.; Naoki, H.; Freitas, J.C.; Machado-Santelli, G.M. Cytoskeleton alterations induced by Geodia corticostylifera depsipeptides in breast cancer cells. Peptides 2006, 27, 2047–2057. [Google Scholar] [CrossRef] [PubMed]
- Freitas, V.M.; Rangel, M.; Bisson, L.F.; Jaeger, R.G.; Machado-Santelli, G.M. The geodiamolide H, derived from Brazilian sponge Geodia corticostylifera, regulates actin cytoskeleton, migration and invasion of breast cancer cells cultured in three-dimensional environment. J. Cell. Physiol. 2008, 216, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, J.; Rechenmacher, F.; Kessler, H. N-Methylation of Peptides and Proteins: An Important Element for Modulating Biological Functions. Angew. Chem. Int. Ed. 2013, 52, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.; Pham, N.B.; Fechner, G.A.; Hooper, J.N.; Quinn, R.J. Potent cytotoxic peptides from the Australian marine sponge Pipestela candelabra. Mar. Drugs 2014, 12, 3399–3415. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Aoki, S.; Ohyabu, N.; Kurosu, M.; Wang, W.; Kitagawa, I. Arenastatin A, a potent cytotoxic depsipeptide from the okinawan marine sponge Dysidea arenaria. Tetrahedron Lett. 1994, 35, 7969–7972. [Google Scholar] [CrossRef]
- Silva, M.A.D.; Bierhalz, A.C.K.; Kieckbusch, T.G. Alginate and pectin composite films crosslinked with Ca2+ ions: Effect of the plasticizer concentration. Carbohydr. Polym. 2009, 77, 736–742. [Google Scholar] [CrossRef]
- Murakami, N.; Tamura, S.; Koyama, K.; Sugimoto, M.; Maekawa, R.; Kobayashi, M. New analogue of arenastatin A, a potent cytotoxic spongean depsipeptide, with anti-tumor activity. Bioorg. Med. Chem. Lett. 2004, 14, 2597–2601. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Tan, R. Isolation and Structure of Phakellistatin 14 from the Western Pacific Marine Sponge Phakellia sp. 1. J. Nat. Prod. 2005, 68, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-L.; Yi, Y.-H.; Wu, H.-M.; Xu, Q.-Z.; Tang, H.-F.; Zhou, D.-Z.; Lin, H.-W.; Wang, Z.-H. Isolation and Structure of the Cytotoxic Cycloheptapeptide Phakellistatin 13. J. Nat. Prod. 2003, 66, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Tan, R.; Ichihara, Y.; Williams, M.D.; Doubek, D.L.; Tackett, L.P.; Schmidt, J.M.; Cerny, R.L.; Boyd, M.R.; Hooper, J.N. Antineoplastic agents, 325. Isolation and structure of the human cancer cell growth inhibitory cyclic octapeptides phakellistatin 10 and 11 from Phakellia sp. J. Nat. Prod. 1995, 58, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Xu, J.-P.; Dorsaz, A.-C.; Williams, M.D.; Boyd, M.R.; Cerny, R.L. Isolation and structure of the human cancer cell growth inhibitory cyclic decapeptides phakellistatins 7, 8 and 9 1, 2. Bioorg. Med. Chem. Lett. 1995, 5, 1339–1344. [Google Scholar] [CrossRef]
- Pelay-Gimeno, M.; Meli, A.; Tulla-Puche, J.; Albericio, F. Rescuing biological activity from synthetic phakellistatin 19. J. Med. Chem. 2013, 56, 9780–9788. [Google Scholar] [CrossRef] [PubMed]
- Zhan, K.-X.; Jiao, W.-H.; Yang, F.; Li, J.; Wang, S.-P.; Li, Y.-S.; Han, B.-N.; Lin, H.-W. Reniochalistatins A–E, cyclic peptides from the marine sponge Reniochalina stalagmitis. J. Nat. Prod. 2014, 77, 2678–2684. [Google Scholar] [CrossRef] [PubMed]
- Edrada-Ebel, R.; Jaspars, M. The 9th European Conference on Marine Natural Products. Mar. Drugs 2015, 13, 7150–7249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ernst, E.; Consortium, C.C. Shark Cartilage: Concerted Action for Complementary and Alternative Medicine Assessment in the Cancer Field (CAM-Cancer); NAFKAM: Tromsø, Norway, 2013; pp. 1–53. [Google Scholar]
- Ulbricht, C.; Hammerness, P.; Barrette, E.-P.; Boon, H.; Szapary, P.; Sollars, D.; Smith, M.; Tsouronis, C.; Bent, S. Shark cartilage monograph: A clinical decision support tool. J. Herb. Pharmacother. 2002, 2, 71–93. [Google Scholar] [CrossRef]
- Dupont, E.; Falardeau, P.; Mousa, S.A.; Dimitriadou, V.; Pepin, M.-C.; Wang, T.; Alaoui-Jamali, M.A. Antiangiogenic and antimetastatic properties of Neovastat (AE-941), an orally active extract derived from cartilage tissue. Clin. Exp. Metastasis 2002, 19, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Gingras, D.; Boivin, D.; Deckers, C.; Gendron, S.; Barthomeuf, C.; Béliveau, R. Neovastat—A novel antiangiogenic drug for cancer therapy. Anti-Cancer Drugs 2003, 14, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Dredge, K. AE-941 (AEterna). Curr. Opin. Investig. Drugs 2004, 5, 668–677. [Google Scholar] [PubMed]
- Zheng, L.; Ling, P.; Wang, Z.; Niu, R.; Hu, C.; Zhang, T.; Lin, X. A novel polypeptide from shark cartilage with potent anti-angiogenic activity. Cancer Biol. Ther. 2007, 6, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-Y.; Lin, W.-J.; Wu, J.-L.; Her, G.M.; Hui, C.-F. Epinecidin-1 peptide induces apoptosis which enhances antitumor effects in human leukemia U937 cells. Peptides 2009, 30, 2365–2373. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-J.; Chien, Y.-L.; Pan, C.-Y.; Lin, T.-L.; Chen, J.-Y.; Chiu, S.-J.; Hui, C.-F. Epinecidin-1, an antimicrobial peptide from fish (Epinephelus coioides) which has an antitumor effect like lytic peptides in human fibrosarcoma cells. Peptides 2009, 30, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-C.; Li-Chan, E.C.; Jao, C.-L. Antiproliferative activity of peptides prepared from enzymatic hydrolysates of tuna dark muscle on human breast cancer cell line MCF-7. Food Chem. 2011, 126, 617–622. [Google Scholar] [CrossRef]
- Song, R.; Wei, R.-B.; Luo, H.-Y.; Yang, Z.-S. Isolation and identification of an antiproliferative peptide derived from heated products of peptic hydrolysates of half-fin anchovy (Setipinna taty). J. Funct. Foods 2014, 10, 104–111. [Google Scholar] [CrossRef]
- Wang, M.; Nie, Y.; Peng, Y.; He, F.; Yang, J.; Wu, C.; Li, X. Purification, characterization and antitumor activities of a new protein from Syngnathus acus, an officinal marine fish. Mar. Drugs 2011, 10, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Júnior, A.C.; Costa, H.M.; Icimoto, M.Y.; Hirata, I.Y.; Marcondes, M.; Carvalho, L.B.; Oliveira, V.; Bezerra, R.S. Giant Amazonian fish pirarucu (Arapaima gigas): Its viscera as a source of thermostable trypsin. Food Chem. 2012, 133, 1596–1602. [Google Scholar] [CrossRef]
- Shai, Y.; Fox, J.; Caratsch, C.; Shih, Y.-L.; Edwards, C.; Lazarovici, P. Sequencing and synthesis of pardaxin, a polypeptide from the Red Sea Moses sole with ionophore activity. FEBS Lett. 1988, 242, 161–166. [Google Scholar] [CrossRef]
- Huang, T.-C.; Lee, J.-F.; Chen, J.-Y. Pardaxin, an antimicrobial peptide, triggers caspase-dependent and ROS-mediated apoptosis in HT-1080 cells. Mar. Drugs 2011, 9, 1995–2009. [Google Scholar] [CrossRef] [PubMed]
- Boivin, D.; Gendron, S.; Beaulieu, É.; Gingras, D.; Béliveau, R. The Antiangiogenic Agent Neovastat (Æ-941) Induces Endothelial Cell Apoptosis 1 Supported by Æterna Laboratories, Québec City, Québec, Canada. 1. Mol. Cancer Ther. 2002, 1, 795–802. [Google Scholar] [PubMed]
- Oren, Z.; Shai, Y. A Class of Highly Potent Antibacterial Peptides Derived from Pardaxin, A Pore-Forming Peptide Isolated from Moses Sole Fish Pardachirus marmoratus. Eur. J. Biochem. 1996, 237, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.-C.; Lin, L.-C.; Tzen, J.T.; Chen, J.-Y. Pardaxin-induced apoptosis enhances antitumor activity in HeLa cells. Peptides 2011, 32, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.-Y.; Lin, C.-N.; Chiou, M.-T.; Yu, C.Y.; Chen, J.-Y.; Chien, C.-H. The antimicrobial peptide pardaxin exerts potent anti-tumor activity against canine perianal gland adenoma. Oncotarget 2015, 6, 2290–2301. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Cui, Z.; Li, Y.-H.; Hsu, W.-H.; Lee, B.-H. In vitro and in vivo anticancer activity of pardaxin against proliferation and growth of oral squamous cell carcinoma. Mar. Drugs 2015, 14, 2. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.K.; Saviola, A.J.; Burns, P.D.; Mackessy, S.P. Apoptosis induction in human breast cancer (MCF-7) cells by a novel venom L-amino acid oxidase (Rusvinoxidase) is independent of its enzymatic activity and is accompanied by caspase-7 activation and reactive oxygen species production. Apoptosis 2015, 20, 1358–1372. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO). The State of World Fisheries and Aquaculture 2016; FAO: Roma, Italy, 2016; p. 200. [Google Scholar]
- Boutayeb, A.; Boutayeb, S. The burden of non communicable diseases in developing countries. Int. J. Equity Health 2005, 4, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahergorabi, R.; Jaczynski, J. Isoelectric solubilization/precipitation as a means to recover protein and lipids from seafood by-products. In Seafood Processing By-Products; Springer: New York, NY, USA, 2014; pp. 101–123. [Google Scholar]
- Schwartsmann, G.; da Rocha, A.B.; Berlinck, R.G.; Jimeno, J. Marine organisms as a source of new anticancer agents. Lancet Oncol. 2001, 2, 221–225. [Google Scholar] [CrossRef]
- Cain, J.M.; Liu, P.; Alberts, D.E.; Gallion, H.H.; Laufman, L.; O’Sullivan, J.; Weiss, G.; Bickers, J.N. Phase II trial of didemnin-B in advanced epithelial ovarian cancer. Investig. New Drugs 1992, 10, 23–24. [Google Scholar] [CrossRef]
- Urdiales, J.; Morata, P.; De Castro, I.N.; Sánchez-Jiménez, F. Antiproliferative effect of dehydrodidemnin B (DDB), a depsipeptide isolated from Mediterranean tunicates. Cancer Lett. 1996, 102, 31–37. [Google Scholar] [CrossRef]
- Suarez-Jimenez, G.-M.; Burgos-Hernandez, A.; Ezquerra-Brauer, J.-M. Bioactive peptides and depsipeptides with anticancer potential: Sources from marine animals. Mar. Drugs 2012, 10, 963–986. [Google Scholar] [CrossRef] [PubMed]
- Kucuk, O.; Young, M.L.; Habermann, T.M.; Wolf, B.C.; Jimeno, J.; Cassileth, P.A. Phase II Trial of Didemnin B in Previously Treated Non-Hodgkin’s Lymphoma: An Eastern Cooperative Oncology Group (ECOG) Study. Am. J. Clin. Oncol. 2000, 23, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, A.B.; Lopes, R.M.; Schwartsmann, G. Natural products in anticancer therapy. Curr. Opin. Pharmacol. 2001, 1, 364–369. [Google Scholar] [CrossRef]
- Cooper, E.L.; Albert, R. Tunicates: A vertebrate ancestral source of antitumor compounds. In Handbook of Anticancer Drugs from Marine Origin; Springer: Berlin, Germany, 2015; pp. 383–395. [Google Scholar]
- Broggini, M.; Marchini, S.; Galliera, E.; Borsotti, P.; Taraboletti, G.; Erba, E.; Sironi, M.; Jimeno, J.; Faircloth, G.; Giavazzi, R. Aplidine, a new anticancer agent of marine origin, inhibits vascular endothelial growth factor (VEGF) secretion and blocks VEGF-VEGFR-1 (flt-1) autocrine loop in human leukemia cells MOLT-4. Leukemia 2003, 17, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Kitagaki, J.; Shi, G.; Miyauchi, S.; Murakami, S.; Yang, Y. Cyclic depsipeptides as potential cancer therapeutics. Anti-Cancer Drugs 2015, 26, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Vervoort, H.; Fenical, W.; Epifanio, R.D.A. Tamandarins A and B: New cytotoxic depsipeptides from a Brazilian ascidian of the family Didemnidae. J. Org. Chem. 2000, 65, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Míguez, A.; Gutiérrez-Jácome, A.; Pérez-Pérez, M.; Pérez-Rodríguez, G.; Catalán-García, S.; Fdez-Riverola, F.; Lourenço, A.; Sánchez, B. From amino acid sequence to bioactivity: The biomedical potential of antitumor peptides. Protein Sci. 2016, 25, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Bowden, B.; Gravalos, D.G. Cyclic Hepta-Peptide Derivative from Colonial Ascidians, Lissoclinum sp. U.S. Patent No. US 20040033940 A1, 3 April 2004. [Google Scholar]
- Kalemkerian, P.G.; Ou, X.; Adil, R.M.; Rosati, R.; Khoulani, M.M.; Madan, K.S.; Pettit, R.G. Activity of dolastatin 10 against small-cell lung cancer in vitro and in vivo: Induction of apoptosis and bcl-2 modification. Cancer Chemother. Pharmacol. 1999, 43, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B 2004, 803, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Margolin, K.; Longmate, J.; Synold, T.W.; Gandara, D.R.; Weber, J.; Gonzalez, R.; Johansen, M.J.; Newman, R.; Baratta, T.; Doroshow, J.H. Dolastatin-10 in Metastatic Melanoma: A Phase II and Pharmokinetic Trial of the California Cancer Consortium. Investig. New Drugs 2001, 19, 335–340. [Google Scholar] [CrossRef]
- Turner, T.; Jackson, W.H.; Pettit, G.R.; Wells, A.; Kraft, A.S. Treatment of human prostate cancer cells with dolastatin 10, a peptide isolated from a marine shell-less mollusc. Prostate 1998, 34, 175–181. [Google Scholar] [CrossRef]
- Zheng, L.; Lin, X.; Wu, N.; Liu, M.; Zheng, Y.; Sheng, J.; Ji, X.; Sun, M. Targeting cellular apoptotic pathway with peptides from marine organisms. BBA Rev. Cancer 2013, 1836, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, X.; Zhao, J.; Xiao, L.; Liu, H.; Wang, C.; Cheng, L.; Wu, N.; Lin, X. Induction of apoptosis, G0/G1 phase arrest and microtubule disassembly in K562 leukemia cells by Mere15, a novel polypeptide from Meretrix meretrix Linnaeus. Mar. Drugs 2012, 10, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, J.; Wu, N.; Liu, M.; Wang, C.; Zhang, Y.; Wang, F.; Liu, H.; Lin, X. Mere15, a novel polypeptide from Meretrix meretrix, inhibits adhesion, migration and invasion of human lung cancer A549 cells via down-regulating MMPs. Pharm. Biol. 2013, 51, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Janmaat, M.L.; Rodriguez, J.A.; Jimeno, J.; Kruyt, F.A.; Giaccone, G. Kahalalide F induces necrosis-like cell death that involves depletion of ErbB3 and inhibition of Akt signaling. Mol. Pharmacol. 2005, 68, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Waters, A.L.; Valeriote, F.A.; Hamann, M.T. An efficient and cost-effective approach to kahalalide F N-terminal modifications using a nuisance algal bloom of Bryopsis pennata. Biochim. Biophys. Acta 2015, 1850, 1849–1854. [Google Scholar] [CrossRef] [PubMed]
- Staats, P.S.; Yearwood, T.; Charapata, S.G.; Presley, R.W.; Wallace, M.S.; Byas-Smith, M.; Fisher, R.; Bryce, D.A.; Mangieri, E.A.; Luther, R.R. Intrathecal ziconotide in the treatment of refractory pain in patients with cancer or AIDS: A randomized controlled trial. JAMA 2004, 291, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Olivier Brenet, M.; Sabine de Bourmont, M.; Florence Dixmerias, M.; Nadia Buisset, M.; Nathalie Lebrec, M.; Dominique Monnin, M. Ziconotide adverse events in patients with cancer pain: A multicenter observational study of a slow titration, multidrug protocol. Pain Phys. 2012, 15, 395–403. [Google Scholar]
- Mundt, S.; Kreitlow, S.; Nowotny, A.; Effmert, U. Biochemical and pharmacological investigations of selected cyanobacteria. Int. J. Hyg. Environ. Health 2001, 203, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, C.C.; Thimmaiah, M.; Shaala, L.A.; Hau, A.M.; Malmo, J.M.; Ishmael, J.E.; Youssef, D.T.; McPhail, K.L. Cyclic depsipeptides, grassypeptolides D and E and Ibu-epidemethoxylyngbyastatin 3, from a Red Sea Leptolyngbya cyanobacterium. J. Nat. Prod. 2011, 74, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Do Rosário Martins, M.; Costa, M. Marine Cyanobacteria Compounds with Anticancer Properties: Implication of Apoptosis. In Handbook of Anticancer Drugs from Marine Origin; Kim, S.-K., Ed.; Springer: Cham, Germany, 2015; pp. 621–647. [Google Scholar]
- Shaala, L.A.; Youssef, D.T.; McPhail, K.L.; Elbandy, M. Malyngamide 4, a new lipopeptide from the Red Sea marine cyanobacterium Moorea producens (formerly Lyngbya majuscula). Phytochem. Lett. 2013, 6, 183–188. [Google Scholar] [CrossRef]
- Wrasidlo, W.; Mielgo, A.; Torres, V.A.; Barbero, S.; Stoletov, K.; Suyama, T.L.; Klemke, R.L.; Gerwick, W.H.; Carson, D.A.; Stupack, D.G. The marine lipopeptide somocystinamide A triggers apoptosis via caspase 8. Proc. Natl. Acad. Sci. USA 2008, 105, 2313–2318. [Google Scholar] [CrossRef] [PubMed]
- Marquez, B.L.; Watts, K.S.; Yokochi, A.; Roberts, M.A.; Verdier-Pinard, P.; Jimenez, J.I.; Hamel, E.; Scheuer, P.J.; Gerwick, W.H. Structure and absolute stereochemistry of hectochlorin, a potent stimulator of actin assembly. J. Nat. Prod. 2002, 65, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Chanda, S.K.; Raya, R.M.; DeJesus, P.D.; Orth, A.P.; Walker, J.R.; Belmonte, J.C.I.; Schultz, P.G. A functional genomics approach to the mode of action of apratoxin A. Nat. Chem. Biol. 2006, 2, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, M.H.; Zhang, X.C.; Chu, X.M. Molecular immune mechanism of c-phycocyanin from Spirulina platensis induces apoptosis in HeLa cells in vitro. Biotechnol. Appl. Biochem. 2006, 43, 155–164. [Google Scholar] [PubMed]
- Thangam, R.; Suresh, V.; Princy, W.A.; Rajkumar, M.; SenthilKumar, N.; Gunasekaran, P.; Rengasamy, R.; Anbazhagan, C.; Kaveri, K.; Kannan, S. C-Phycocyanin from Oscillatoria tenuis exhibited an antioxidant and in vitro antiproliferative activity through induction of apoptosis and G0/G1 cell cycle arrest. Food Chem. 2013, 140, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Lau, N.-S.; Matsui, M.; Abdullah, A.A.-A. Cyanobacteria: Photoautotrophic microbial factories for the sustainable synthesis of industrial products. BioMed Res. Int. 2015, 2015, 754934. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.H.; Wijesekara, I.; Vo, T.S.; Van Ta, Q.; Kim, S.K. Marine food-derived functional ingredients as potential antioxidants in the food industry: An overview. Food Res. Int. 2011, 44, 523–529. [Google Scholar] [CrossRef]
- Nazeer, R.; Prabha, K.D.; Kumar, N.S.; Ganesh, R.J. Isolation of antioxidant peptides from clam, Meretrix casta (Chemnitz). J. Food Sci. Technol. 2013, 50, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.; Mendis, E.; Byun, H.-G.; Kim, S.-K. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem. 2005, 16, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Zhong, Y. Bioactive Peptides. J. AOAC Int. 2008, 91, 914–931. [Google Scholar] [PubMed]
- Je, J.-Y.; Park, P.-J.; Kim, S.-K. Antioxidant activity of a peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. Food Res. Int. 2005, 38, 45–50. [Google Scholar] [CrossRef]
- Jun, S.-Y.; Park, P.-J.; Jung, W.-K.; Kim, S.-K. Purification and characterization of an antioxidative peptide from enzymatic hydrolysate of yellowfin sole (Limanda aspera) frame protein. Eur. Food Res. Technol. 2004, 219, 20–26. [Google Scholar]
- Mendis, E.; Rajapakse, N.; Kim, S.-K. Antioxidant properties of a radical-scavenging peptide purified from enzymatically prepared fish skin gelatin hydrolysate. J. Agric. Food Chem. 2005, 53, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.; Ryu, B.; Ngo, D.-H.; Kim, S.-K. Peptide Isolated From Japanese Flounder Skin Gelatin Protects Against Cellular Oxidative Damage. J. Agric. Food Chem. 2012, 60, 9112–9119. [Google Scholar] [CrossRef] [PubMed]
- Guérard, F.; Decourcelle, N.; Sabourin, C.; Floch-Laizet, C.; Le Grel, L.; Le Floc’H, P.; Gourlay, F.; Le Delezir, R.; Jaouen, P.; Bourseau, P. Recent developments of marine ingredients for food and nutraceutical applications: A review. J. Sci. Halieut. Aquat. 2010, 2, 21–27. [Google Scholar]
- Zhu, C.-F.; Peng, H.-B.; Liu, G.-Q.; Zhang, F.; Li, Y. Beneficial effects of oligopeptides from marine salmon skin in a rat model of type 2 diabetes. Nutrition 2010, 26, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.-F.; Li, G.-Z.; Peng, H.-B.; Zhang, F.; Chen, Y.; Li, Y. Effect of marine collagen peptides on markers of metabolic nuclear receptors in type 2 diabetic patients with/without hypertension. Biomed. Environ. Sci. 2010, 23, 113–120. [Google Scholar] [CrossRef]
- Zhu, C.-F.; Li, G.-Z.; Peng, H.-B.; Zhang, F.; Chen, Y.; Li, Y. Treatment with marine collagen peptides modulates glucose and lipid metabolism in Chinese patients with type 2 diabetes mellitus. Appl. Physiol. Nutr. Metab. 2010, 35, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Chance, W.T.; Tao, Z.; Sheriff, S.; Balasubramaniam, A. WRYamide, A NPY-based tripeptide that antagonizes feeding in rats. Brain Res. 1998, 803, 39–43. [Google Scholar] [CrossRef]
- Vernaleken, A.; Veyhl, M.; Gorboulev, V.; Kottra, G.; Palm, D.; Burckhardt, B.-C.; Burckhardt, G.; Pipkorn, R.; Beier, N.; van Amsterdam, C. Tripeptides of RS1 (RSC1A1) inhibit a monosaccharide-dependent exocytotic pathway of Na+-d-glucose cotransporter SGLT1 with high affinity. J. Biol. Chem. 2007, 282, 28501–28513. [Google Scholar] [CrossRef] [PubMed]
- Chevrier, G.; Mitchell, P.L.; Rioux, L.-E.; Hasan, F.; Jin, T.; Roblet, C.R.; Doyen, A.; Pilon, G.; St-Pierre, P.; Lavigne, C. Low-molecular-weight peptides from salmon protein prevent obesity-linked glucose intolerance, inflammation, and dyslipidemia in LDLR−/−/ApoB100/100 mice. J. Nutr. 2015, 145, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: The link between insulin resistance, obesity and diabetes. Trends Immunol. 2004, 25, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Shimizu, K.; Kaneko, H.; Shibayama, F.; Morikawa, K.; Kanamaru, Y.; Otsuka, A.; Hirahashi, T.; Kato, T. A novel protein c-phycocyanin plays a crucial role in the hypocholesterolemic action of Spirulina platensis concentrate in rats. J. Nutr. 2005, 135, 2425–2430. [Google Scholar] [PubMed]
- Colla, L.M.; Muccillo-Baisch, A.L.; Costa, J.A.V. Spirulina platensis effects on the levels of total cholesterol, HDL and triacylglycerols in rabbits fed with a hypercholesterolemic diet. Braz. Arch. Biol. Technol. 2008, 51, 405–411. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine Bioactives as Functional Food Ingredients: Potential to Reduce the Incidence of Chronic Diseases. Mar. Drugs 2011, 9, 1056–1110. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The future of peptide-based drugs. Chem. Biol. Drug Des. 2013, 81, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Chandrasekara, A. Millet grain phenolics and their role in disease risk reduction and health promotion: A review. J. Funct. Foods 2013, 5, 570–581. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Sun, L.; Zhang, Y.; Liu, G. Antihypertensive Effect of Long-Term Oral Administration of Jellyfish (Rhopilema esculentum) Collagen Peptides on Renovascular Hypertension. Mar. Drugs 2012, 10, 417–426. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Source | Extraction | Sequence | Inhibition (IC50) | References |
|---|---|---|---|---|
| Seaweed (Undaria pinnatifida) | Hot water extraction; Chromatography | Ile-Tyr | 2.7 μM | [18] |
| Enzymatic hydrolysis (Protease S); Chromatography | Ile-Trp | 1.5 μM | [30] | |
| Seaweed (P. yezoensis) | Chromatography | Ala-Lys-Tyr-Ser-Tyr | 1.52 μM | [31] |
| Microalgae (Spirulina platensis) | Enzymatic hydrolysis (Pepsin); Chromatography | Ile-Ala-Pro-Gly | 11.4 μM | [32] |
| Yellowfin tuna (Neothunnus macropterus) | Chromatography | Pro-Thr-His-Ile-Lys-Trp-Gly-Asp | 2 μM | [33] |
| Skipjack tuna (Katsuwonus pelamis) bowels | Chromatography | Leu-Arg-Pro | 1 μM | [34] |
| Alaska Pollack skin (Theragra chalcogramma) | Enzymatic hydrolysis (serial protease); Chromatography | Gly-Pro-Leu | 2.6 μM | [35] |
| Chum salmon (Oncorhynchus keta) muscle | Enzymatic hydrolysis (Thermolysin); Chromatography | Val-Trp | 2.5 μM | [36] |
| Pink salmon (Oncorhynchus gorbuscha) | Enzymatic hydrolysis (papain); Chromatography | Ile-Trp | 1.2 μM | [37] |
| Skate skin (Okamejei kenojei) | Enzymatic hydrolysis (alkalase/protease); Chromatography | Met-Val-Gly-Ser-Ala-Pro-Gly-Val-Leu | 3.09 μM | [38] |
| Small-spotted catshark (Scyliorhinus canicula) | Enzymatic hydrolysis (Trypsin, subtilisin); Chromatography | Val-Ala-Met-Pro-Phe | 0.44 μM | [39] |
| Pelagic thresher (Alopias pelagicus) muscle | Enzymatic hydrolysis (thermolysin); Chromatography | Ile-Lys-Trp | 0.54 μM | [26] |
| Marine shrimp (Acetes chinensis) | Enzymatic hydrolysis (Protease); Chromatography | Ile-Phe-Val-Pro-Ala-Phe | 3.4 μM | [40] |
| Fermentation; Chromatography | Asp-Pro | 2.15 μM | [41] | |
| Enzymatic hydrolysis (Pepsin); Chromatography | Leu-His-Pro | 3.4 μM | [42] | |
| Izumi shrimp (Plesionika izumiae Omori, 1971) | Enzymatic hydrolysis (Protease); Chromatography | Ser-Thr | 4.03 μM | [43] |
| Jellyfish (Rhopilema esculentum) | Enzymatic hydrolysis (pepsin, papain); ultrafiltration; Chromatography | Gln-Pro-Gly-Pro-Thr | 80.67 μM | [44] |
| Sipuncula (Phascolosoma esculenta) | Enzymatic hydrolysis (Pepsin); Chromatography | Ala-Trp-Leu-His-Pro-Gly-Ala-Pro-Lys-Val-Phe | 135 M | [45] |
| Pearl oyster (Pinctada fucata martensii) | Enzymatic hydrolysis (Pepsin); Chromatography | Ala-Leu-Ala-Pro-Glu | 167.5 μM | [46] |
| Name | Source | Anti-Cancer Activity | References |
|---|---|---|---|
| Neovastat (AE-941) | Spiny dogfish shark (Squalus acanthias) | Inhibition of metastatic activity on HUVEC, BAEC cells; inhibition of matrix metalloproteinase; Anti-angiogenic effects; Pro-apoptotic on BAEC cells | [78,90] |
| Pardaxin | Red Sea Moses sole (Pardachirus marmoratus) | Pro-apoptotic on HT1080 (IC50: 14.52–15.74 μg/mL), HeLa, OSCC cells | [89,91,92,93,94] |
| PG155 | Blue shark (Prionace glauca) | Anti-angiogenic effects on HUVECs | [81] |
| Syngnathusin | Pipefish (Syngnathus acus) | Pro-apoptotic on A549 (IC50: 84.9 μg/mL), and CCRF-CEM (IC50: 215.3 μg/mL), cells | [86] |
| Epinecidin-1 | Grouper (Epinephelus coioides) | Anti-angiogenic effects on A549, HA59T/VGH, HeLa, HepG2, and HT1080 cells Pro-apoptotic on U937 cells | [82,83] |
| PAB 1; PAB2 | Long tail tuna (Thunnus tonggol) | Pro-apoptotic on MCF-7 cells (IC50: 8.1; 8.8 μM) | [84] |
| YALRAH | Half-fin anchovy (Setipinna taty) | Pro-apoptotic on PC-3 cells (IC50: 11.1 μM) | [85] |
| Rusvinoxidase | Venom of Daboia russelii russelii | Pro-apoptotic on MCF-7 cells (IC50: 83 nM) | [95] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pangestuti, R.; Kim, S.-K. Bioactive Peptide of Marine Origin for the Prevention and Treatment of Non-Communicable Diseases. Mar. Drugs 2017, 15, 67. https://doi.org/10.3390/md15030067
Pangestuti R, Kim S-K. Bioactive Peptide of Marine Origin for the Prevention and Treatment of Non-Communicable Diseases. Marine Drugs. 2017; 15(3):67. https://doi.org/10.3390/md15030067
Chicago/Turabian StylePangestuti, Ratih, and Se-Kwon Kim. 2017. "Bioactive Peptide of Marine Origin for the Prevention and Treatment of Non-Communicable Diseases" Marine Drugs 15, no. 3: 67. https://doi.org/10.3390/md15030067
APA StylePangestuti, R., & Kim, S. -K. (2017). Bioactive Peptide of Marine Origin for the Prevention and Treatment of Non-Communicable Diseases. Marine Drugs, 15(3), 67. https://doi.org/10.3390/md15030067