Perna canaliculus and the Intestinal Microbiome


Abstract
:1. Introduction
2. Intestinal Bacterial Cohort
3. Bioactive Molecules and Compounds from Green-Lipped Mussel
3.1. Mechanism of Intestinal Actions
3.2. Immunomodulation
3.3. Anti-Microbial Activities
3.4. Cardiovascular Effects
4. The Microbiome and Omega-3 Fatty Acids
4.1. Changes in the Intestinal Microbiome
4.2. SCFAs as Signaling Molecules for End-Organ Health
4.3. Gut Synthesis of Butyrate
5. Discussion
6. Conclusions
Author Contributions
Conflicts of Interest
References
- Schloss, P.D.; Girard, R.A.; Martin, T.; Edwards, J.; Thrash, J.C. Status of the archaeal and bacterial census: An update. mBio 2016, 7, e00201-16. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.; Rossello-Mora, R. After all, only millions? mBio 2016, 7, e00999-16. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Fernandez, L.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodriguez, J.M. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [PubMed]
- Trosvik, P.; de Muinck, E.J. Ecology of bacteria in the human gastrointestinal tract—Identification of keystone and foundation taxa. Microbiome 2015, 3, 44. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Hall, S.; Linnane, A.W. Live probiotic cultures and the gastrointestinal tract: Symbiotic preservation of tolerance whilst attenuating pathogenicity. Front. Cell. Infect. Microbiol. 2014, 4, 143. [Google Scholar] [CrossRef] [PubMed]
- Egert, M.; de Graaf, A.A.; Smidt, H.; de Vos, W.M.; Venema, K. Beyond diversity: Functional microbiomics of the human colon. Trends Microbiol. 2006, 14, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Hutkins, R.W.; Krumbeck, J.A.; Bindels, L.B.; Cani, P.D.; Fahey, G., Jr.; Goh, Y.J.; Hamaker, B.; Martens, E.C.; Mills, D.A.; Rastal, R.A.; et al. Prebiotics: Why definitions matter. Curr. Opin. Biotechnol. 2016, 37, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; de Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Kim, S.S. Probiotics and prebiotics: Present status and future perspectives on metabolic disorders. Nutrients 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed]
- Schilderink, R.; Verseijden, C.; de Jonge, W.J. Dietary inhibitors of histone deacetylases in intestinal immunity and homeostasis. Front. Immunol. 2013, 4, 226. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Nian, L.; Kwok, L.Y.; Sun, T.; Zhao, J. Reduction in fecal microbiota diversity and short-chain fatty acid producers in methicillin-resistant staphylococcus aureus infected individuals as revealed by pacbio single molecule, real-time sequencing technology. Eur. J. Clin. Microbiol. Infect. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104, S1–S63. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.; Stolarczyk, E.; Mouni, F.; Brassart, C.; Vidal, O.; Aissi, E.; Bouquelet, S.; Krzewinski, F. Characterisation of glutamine fructose-6-phosphate amidotransferase (ec 2.6.1.16) and n-acetylglucosamine metabolism in bifidobacterium. Arch. Microbiol. 2008, 189, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Koser, S.A.; Tribby, I.; Stuedell, J.T. Glucosamine utilization by some lactic acid bacteria. J. Infect. Dis. 1961, 108, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Coulson, S.; Butt, H.; Vecchio, P.; Gramotnev, H.; Vitetta, L. Green-lipped mussel extract (perna canaliculus) and glucosamine sulphate in patients with knee osteoarthritis: Therapeutic efficacy and effects on gastrointestinal microbiota profiles. Inflammopharmacology 2013, 21, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Rastall, R.A.; Gibson, G.R. Recent developments in prebiotics to selectively impact beneficial microbes and promote intestinal health. Curr. Opin. Biotechnol. 2015, 32, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Kellow, N.J.; Coughlan, M.T.; Reid, C.M. Metabolic benefits of dietary prebiotics in human subjects: A systematic review of randomised controlled trials. Br. J. Nutr. 2014, 111, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Adamu, B.O.; Lawley, T.D. Bacteriotherapy for the treatment of intestinal dysbiosis caused by clostridium difficile infection. Curr. Opin. Microbiol. 2013, 16, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Hall, S.; Coulson, S. Metabolic interactions in the gastrointestinal tract (git): Host, commensal, probiotics, and bacteriophage influences. Microorganisms 2015, 3, 913–932. [Google Scholar] [CrossRef] [PubMed]
- Lawson, B.R.; Belkowski, S.M.; Whitesides, J.F.; Davis, P.; Lawson, J.W. Immunomodulation of murine collagen-induced arthritis by n, n-dimethylglycine and a preparation of perna canaliculus. BMC Complement. Altern. Med. 2007, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.R.; Apte, S.; MacAvoy, E.S.; Gardner, J.P. A molecular phylogeny of the marine mussel genus perna (bivalvia: Mytilidae) based on nuclear (its1&2) and mitochondrial (coi) DNA sequences. Mol. Phylogenet. Evol. 2007, 44, 685–698. [Google Scholar] [PubMed]
- Zawadzki, M.; Janosch, C.; Szechinski, J. Perna canaliculus lipid complex pcso-524 demonstrated pain relief for osteoarthritis patients benchmarked against fish oil, a randomized trial, without placebo control. Mar. Drugs 2013, 11, 1920–1935. [Google Scholar] [CrossRef] [PubMed]
- Coulson, S.; Vecchio, P.; Gramotnev, H.; Vitetta, L. Green-lipped mussel (perna canaliculus) extract efficacy in knee osteoarthritis and improvement in gastrointestinal dysfunction: A pilot study. Inflammopharmacology 2012, 20, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Mickleborough, T.D.; Vaughn, C.L.; Shei, R.J.; Davis, E.M.; Wilhite, D.P. Marine lipid fraction pcso-524 (lyprinol/omega xl) of the new zealand green lipped mussel attenuates hyperpnea-induced bronchoconstriction in asthma. Respir. Med. 2013, 107, 1152–1163. [Google Scholar] [CrossRef] [PubMed]
- Tenikoff, D.; Murphy, K.J.; Le, M.; Howe, P.R.; Howarth, G.S. Lyprinol (stabilised lipid extract of new zealand green-lipped mussel): A potential preventative treatment modality for inflammatory bowel disease. J. Gastroenterol. 2005, 40, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Scotti, P.D.; Dearing, S.C.; Greenwood, D.R.; Newcomb, R.D. Pernin: A novel, self-aggregating haemolymph protein from the new zealand green-lipped mussel, perna canaliculus (bivalvia: Mytilidae). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 128, 767–779. [Google Scholar] [CrossRef]
- Jung, W.-K.; Kim, S.-K. Isolation and characterisation of an anticoagulant oligopeptide from blue mussel, mytilus edulis. Food Chem. 2009, 117, 687–692. [Google Scholar] [CrossRef]
- Abdulazim, A.; Hädrich, M.; Montani, M.; Semmo, N. Acute hepatitis induced by lyprinol, the lipid extract of the green-lipped mussel (perna canaliculus), in a patient with polyarthrosis. Case Rep. Hepatol. 2012, 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, S.; Pittman, K.B.; Patterson, W.K.; Dickson, J.; Yeend, S.; Townsend, A.; Broadbridge, V.; Price, T.J. A phase I study to determine the safety, tolerability and maximum tolerated dose of green-lipped mussel (perna canaliculus) lipid extract, in patients with advanced prostate and breast cancer. Ann. Oncol. 2010, 21, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.; Wu, H. In vivo evidence for prostaglandin inhibitory activity in new zealand green-lipped mussel extract. N. Z. Med. J. 1984, 97, 355–357. [Google Scholar] [PubMed]
- Coulson, S.; Palacios, T.; Vitetta, L. Perna canaliculus (green-lipped mussel): Bioactive components and therapeutic evaluation for chronic health conditions. Prog. Drug Res. 2015, 70, 91–132. [Google Scholar] [PubMed]
- Whitehouse, M.W.; Butters, D.E. Combination anti-inflammatory therapy: Synergism in rats of nsaids/corticosteroids with some herbal/animal products. Inflammopharmacology 2003, 11, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, M.W. Anti-tnf-alpha therapy for chronic inflammation: Reconsidering pentoxifylline as an alternative to therapeutic protein drugs. Inflammopharmacology 2004, 12, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Rainsford, K.D.; Whitehouse, M.W. Gastroprotective and anti-inflammatory properties of green lipped mussel (perna canaliculus) preparation. Arzneimittel-Forschung 1980, 30, 2128–2132. [Google Scholar] [PubMed]
- Mani, S.; Lawson, J.W. In vitro modulation of inflammatory cytokine and igg levels by extracts of perna canaliculus. BMC Complement. Altern. Med. 2006, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Grienke, U.; Silke, J.; Tasdemir, D. Bioactive compounds from marine mussels and their effects on human health. Food Chem. 2014, 142, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kandasamy, S.; Zhang, J.; Kirby, C.W.; Karakach, T.; Hafting, J.; Critchley, A.T.; Evans, F.; Prithiviraj, B. Prebiotic effects of diet supplemented with the cultivated red seaweed chondrus crispus or with fructo-oligo-saccharide on host immunity, colonic microbiota and gut microbial metabolites. BMC Complement. Altern. Med. 2015, 15, 279. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, T.; Kondo, H.; Nii, H.; Kimura, K.; Egami, Y.; Oka, Y.; Yoshida, M.; Kida, E.; Ye, Y.; Akahoshi, S.; et al. Furan fatty acid as an anti-inflammatory component from the green-lipped mussel perna canaliculus. Proc. Nat. Acad. Sci. USA 2011, 108, 17533–17537. [Google Scholar] [CrossRef] [PubMed]
- Charlet, M.; Chernysh, S.; Philippe, H.; Hetru, C.; Hoffmann, J.A.; Bulet, P. Innate immunity. Isolation of several cysteine-rich antimicrobial peptides from the blood of a mollusc, mytilus edulis. J. Biol. Chem. 1996, 271, 21808–21813. [Google Scholar] [CrossRef] [PubMed]
- Fredrick, W.S.; Ravichandran, S. Hemolymph proteins in marine crustaceans. Asian Pac. J. Trop. Biomed. 2012, 2, 496–502. [Google Scholar] [CrossRef]
- Je, J.Y.; Park, P.J.; Byun, H.G.; Jung, W.K.; Kim, S.K. Angiotensin I converting enzyme (ace) inhibitory peptide derived from the sauce of fermented blue mussel, mytilus edulis. Biores. Technol. 2005, 96, 1624–1629. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.J.; Mooney, B.D.; Mann, N.J.; Nichols, P.D.; Sinclair, A.J. Lipid, fa, and sterol composition of new zealand green lipped mussel (perna canaliculus) and tasmanian blue mussel (mytilus edulis). Lipids 2002, 37, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Naughton, J.M.; O’Dea, K.; Sinclair, A.J. Animal foods in traditional australian aboriginal diets: Polyunsaturated and low in fat. Lipids 1986, 21, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.G.; Gibson, S.L. Green-lipped mussel extract in arthritis. Lancet 1981, 1, 439. [Google Scholar] [CrossRef]
- Audeval, B.; Bouchacourt, P. Double-blind trial against placebo extract of perna canaliculus (green-lipped mussel) in osteoarthritis of the knee. Gazette Med. 1986, 93, 111–116. [Google Scholar]
- Gibson, S.L.M.; Gibson, R.G. The treatment of arthritis with a lipid extract of perna canaliculus: A randomized trial. Complement. Ther. Med. 1998, 6, 122–126. [Google Scholar] [CrossRef]
- Lau, C.S.; Chiu, P.K.Y.; Chu, E.M.Y.; Cheng, I.Y.W.; Tang, W.M.; Man, R.Y.K.; Halpern, G.M. Treatment of knee osteoarthritis with lyprinol®, lipid extract of the green-lipped mussel—A double-blind placebo-controlled study. Prog. Nutr. 2004, 6, 17–31. [Google Scholar]
- Highton, T.C.; McArthur, A.W. Pilot study on the effect of new zealand green mussel on rheumatoid arthritis. N. Z. Med. J. 1975, 81, 261–262. [Google Scholar] [PubMed]
- Caughey, D.E.; Grigor, R.R.; Caughey, E.B.; Young, P.; Gow, P.J.; Stewart, A.W. Perna canaliculus in the treatment of rheumatoid arthritis. Eur. J. Rheumatol. Inflamm. 1983, 6, 197–200. [Google Scholar] [PubMed]
- Larkin, J.G.; Capell, H.A.; Sturrock, R.D. Seatone in rheumatoid arthritis: A six-month placebo-controlled study. Ann. Rheum. Dis. 1985, 44, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Puente, R.; Illnait, J.; Mas, R.; Mendoza, S.; Carbajal, D.; Fernández, J.; Mesa, M.; Prieto, Y. Effects of d-002, a mixture of beeswax alcohols, co-administered with green-lipped mussel extract, on osteoarthritis symptoms. Int. J. Pharm. Sci. Rev. Res. 2014, 27, 209–216. [Google Scholar]
- Cho, S.H.; Jung, Y.B.; Seong, S.C.; Park, H.B.; Byun, K.Y.; Lee, D.C.; Song, E.K.; Son, J.H. Clinical efficacy and safety of lyprinol, a patented extract from new zealand green-lipped mussel (perna canaliculus) in patients with osteoarthritis of the hip and knee: A multicenter 2-month clinical trial. Eur. Ann. Allergy Clin. Immunol. 2003, 35, 212–216. [Google Scholar] [PubMed]
- Rajkumar, H.; Mahmood, N.; Kumar, M.; Varikuti, S.R.; Challa, H.R.; Myakala, S.P. Effect of probiotic (vsl#3) and omega-3 on lipid profile, insulin sensitivity, inflammatory markers, and gut colonization in overweight adults: A randomized, controlled trial. Mediat. Inflamm. 2014, 2014, 348959. [Google Scholar]
- Yu, H.N.; Zhu, J.; Pan, W.S.; Shen, S.R.; Shan, W.G.; Das, U.N. Effects of fish oil with a high content of n-3 polyunsaturated fatty acids on mouse gut microbiota. Arch. Med. Res. 2014, 45, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Balfego, M.; Canivell, S.; Hanzu, F.A.; Sala-Vila, A.; Martinez-Medina, M.; Murillo, S.; Mur, T.; Ruano, E.G.; Linares, F.; Porras, N.; et al. Effects of sardine-enriched diet on metabolic control, inflammation and gut microbiota in drug-naive patients with type 2 diabetes: A pilot randomized trial. Lipids Health Dis. 2016, 15, 78. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; O’Doherty, R.M.; Murphy, E.F.; Wall, R.; O’Sullivan, O.; Nilaweera, K.; Fitzgerald, G.F.; Cotter, P.D.; Ross, R.P.; Stanton, C. Impact of dietary fatty acids on metabolic activity and host intestinal microbiota composition in c57bl/6j mice. Br. Nutr. 2014, 111, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Kaliannan, K.; Wang, B.; Li, X.Y.; Kim, K.J.; Kang, J.X. A host-microbiome interaction mediates the opposing effects of omega-6 and omega-3 fatty acids on metabolic endotoxemia. Sci. Rep. 2015, 5, 11276. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; DeCoffe, D.; Brown, K.; Rajendiran, E.; Estaki, M.; Dai, C.; Yip, A.; Gibson, D.L. Fish oil attenuates omega-6 polyunsaturated fatty acid-induced dysbiosis and infectious colitis but impairs lps dephosphorylation activity causing sepsis. PLoS ONE 2013, 8, e55468. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.P.; Ritari, J.; Boeren, S.; de Waard, P.; Plugge, C.M.; de Vos, W.M. Production of butyrate from lysine and the amadori product fructoselysine by a human gut commensal. Nat. Commun. 2015, 6, 10062. [Google Scholar] [CrossRef] [PubMed]
- Rios-Covian, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilan, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Basson, A.; Trotter, A.; Rodriguez-Palacios, A.; Cominelli, F. Mucosal interactions between genetics, diet, and microbiome in inflammatory bowel disease. Front. Immunol. 2016, 7, 290. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, M.M.; El Aidy, S.; Crispie, F.; O’Sullivan, O.; Cotter, P.; Stanton, C.; Kelly, P.; Cryan, J.F.; Dinan, T.G. N-3 polyunsaturated fatty acids (pufas) reverse the impact of early-life stress on the gut microbiota. PLoS ONE 2015, 10, e0139721. [Google Scholar]
- Noriega, B.S.; Sanchez-Gonzalez, M.A.; Salyakina, D.; Coffman, J. Understanding the impact of omega-3 rich diet on the gut microbiota. Case Rep. Med. 2016, 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Nakada, H.I.; Wolfe, J.B. Glucosamine degradation by Escherichia coli. II. The isomeric conversion of glucosamine 6-po4 to fructose 6-po4 and ammonia. Arch. Biochem. Biophys. 1956, 64, 489–497. [Google Scholar] [PubMed]
- Gómez de Segura, I.A.; Valderrábano, S.; Vázquez, I.; Vallejo-Cremades, M.T.; Gόmez-García, L.; Sánchez, M.; de Miguel, E. Protective effects of dietary enrichment with docosahexaenoic acid plus protein in 5-fluorouracil-induced intestinal injury in the rat. Eur. J. Gastroenterol. Hepatol. 2004, 16, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Rani, I.; Vaiphei, K.; Agnihotri, N. Supplementation of fish oil augments efficacy and attenuates toxicity of 5-fluorouracil in 1,2-dimethylhydrazine dihydrochloride/dextran sulfate sodium-induced colon carcinogenesis. Cancer Chemother. Pharmacol. 2014, 74, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Torres, D.M.; Tooley, K.L.; Butler, R.N.; Smith, C.L.; Geier, M.S.; Howarth, G.S. Lyprinol only partially improves indicators of small intestinal integrity in a rat model of 5-fluorouracil-induced mucositis. Cancer Biol. Ther. 2008, 7, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Sebe, M.; Tsutsumi, R.; Yamaguchi, S.; Horikawa, Y.T.; Harada, N.; Oyama, T.; Kakuta, N.; Tanaka, K.; Tsutsumi, Y.M.; Nakaya, Y.; et al. The synergystic effects of omega-3 fatty acids against 5-fluorouracil-induced mucosal impairment in mice. BMC Nutr. 2016, 2, 17. [Google Scholar] [CrossRef]
- Janczyk, W.; Socha, P.; Lebensztejn, D.; Wierzbicka, A.; Mazur, A.; Neuhoff-Murawska, J.; Matusik, P. Omega-3 fatty acids for treatment of non-alcoholic fatty liver disease: Design and rationale of randomized controlled trial. BMC Pediatr. 2013, 13, 85. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.-H.; Nichols, R.G.; Sebastian, A.; Albert, I.; Patterson, A.D.; Ross, A.C. Gut microbiota increased by omega-3 fatty acids is negatively correlated with hepatic lipid metabolism-associated genes in mice with high carbohydrate diet-induced steatosis. FASEB J. 2017, 31, 654.3. [Google Scholar]
- Juliano, C.; Manconi, P.; Cossu, M. Characterisation of commercial perna canaliculus samples and development of extemporaneous oral veterinary paste formulations containing perna. Nat. Prod. Res. 2016, 30, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
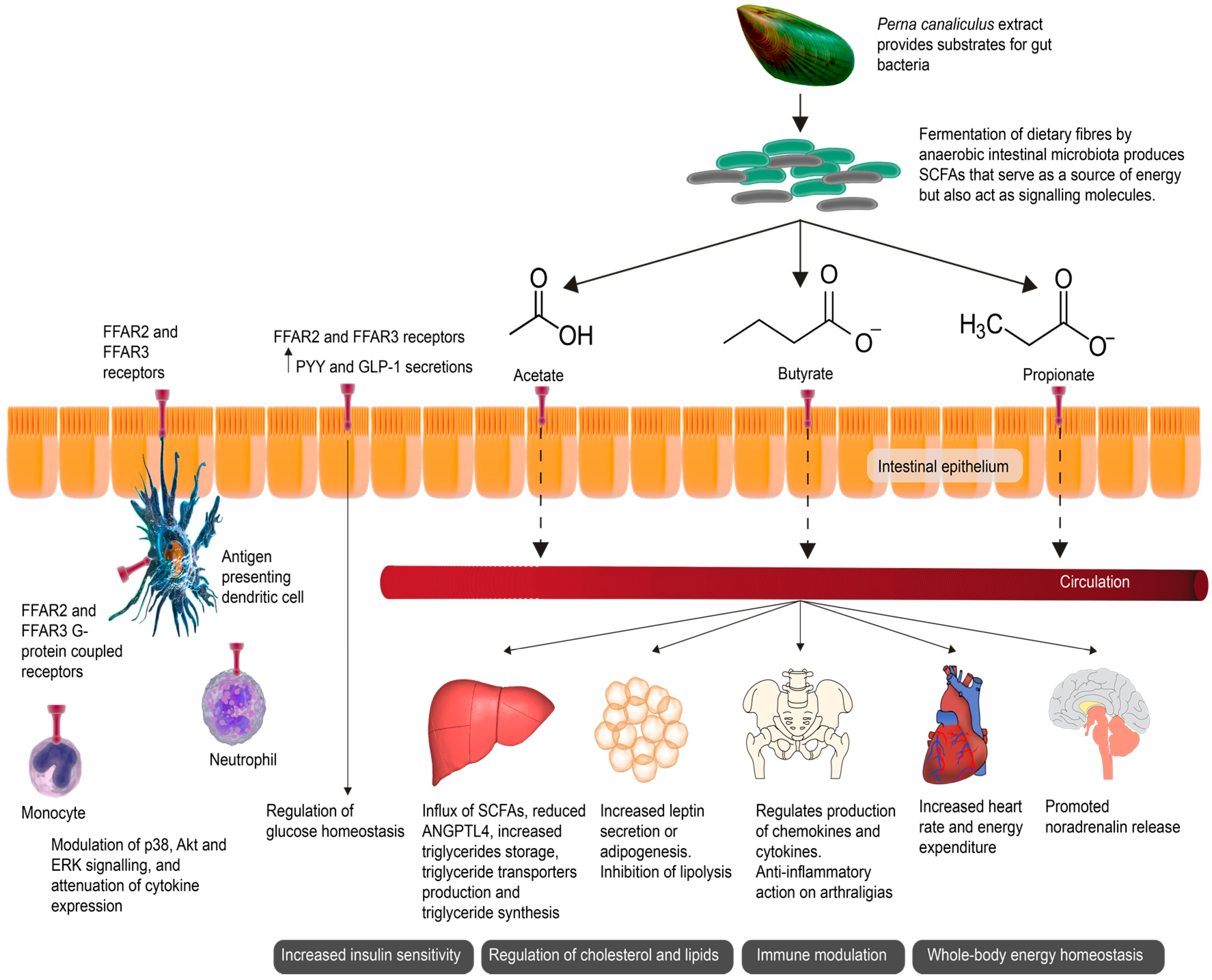

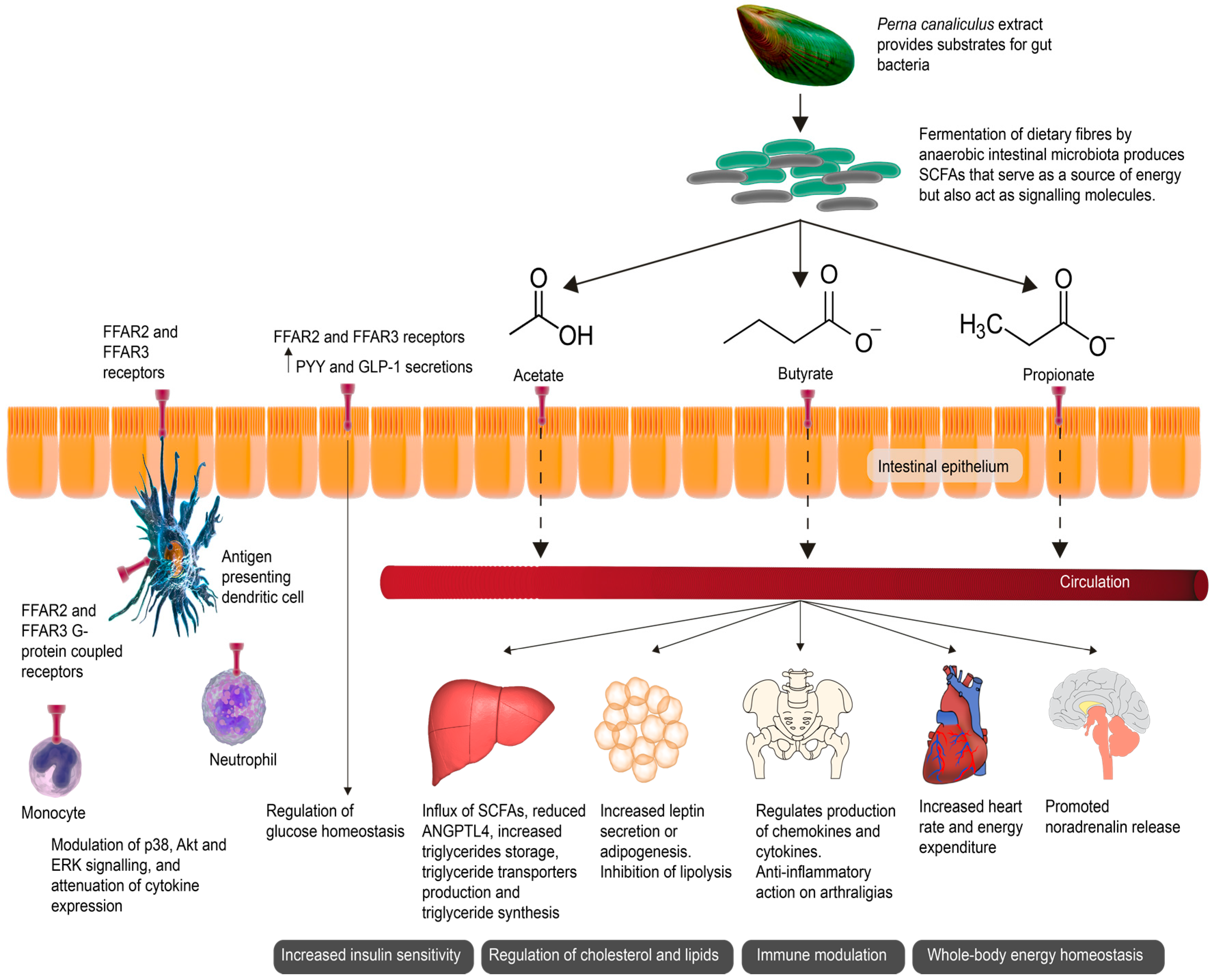{kind=link}
| Study Type (n) | Intervention/Control | Results |
|---|---|---|
| RCT (n = 38) [46] |
|
|
| RCT (n = 53) [47] |
|
|
| RCT (n = 30) [48] |
|
|
| RCT (n = 80) [49] |
|
|
| RCT (n = 6) [50] |
|
|
| RCT (n = 47) [51] |
|
|
| RCT (n = 35) [52] |
|
|
| RCT (n = 38) [17] |
|
|
| RCT (n = 50) [53] |
|
|
| RCT (n = 50) [24] |
|
|
| Open clinical trial (n = 60) [54] |
|
|
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saltzman, E.T.; Thomsen, M.; Hall, S.; Vitetta, L. Perna canaliculus and the Intestinal Microbiome. Mar. Drugs 2017, 15, 207. https://doi.org/10.3390/md15070207
Saltzman ET, Thomsen M, Hall S, Vitetta L. Perna canaliculus and the Intestinal Microbiome. Marine Drugs. 2017; 15(7):207. https://doi.org/10.3390/md15070207
Chicago/Turabian StyleSaltzman, Emma Tali, Michael Thomsen, Sean Hall, and Luis Vitetta. 2017. "Perna canaliculus and the Intestinal Microbiome" Marine Drugs 15, no. 7: 207. https://doi.org/10.3390/md15070207
APA StyleSaltzman, E. T., Thomsen, M., Hall, S., & Vitetta, L. (2017). Perna canaliculus and the Intestinal Microbiome. Marine Drugs, 15(7), 207. https://doi.org/10.3390/md15070207