Accumulation of Microcystin (LR, RR and YR) in Three Freshwater Bivalves in Microcystis aeruginosa Bloom Using Dual Isotope Tracer

 ,
, 
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
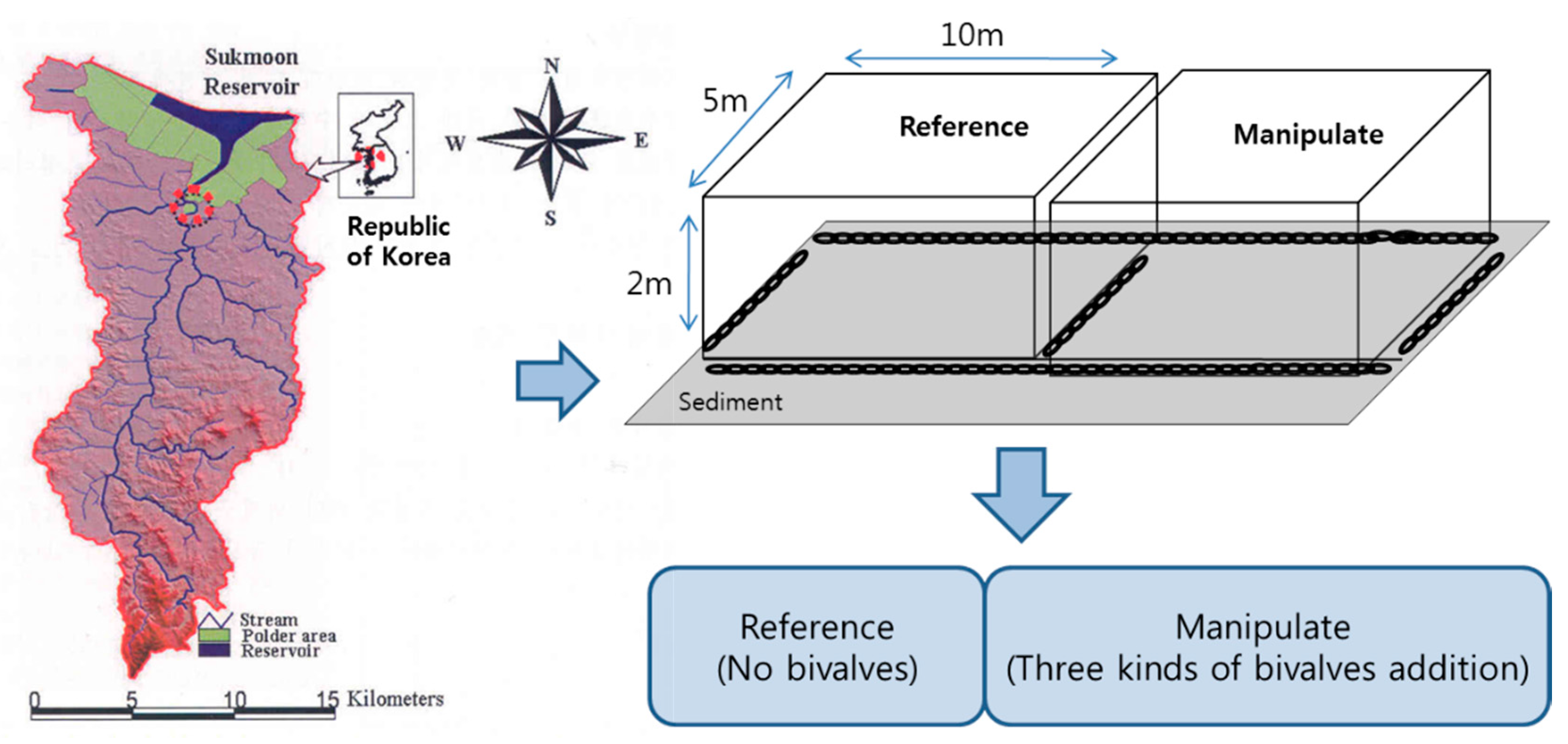
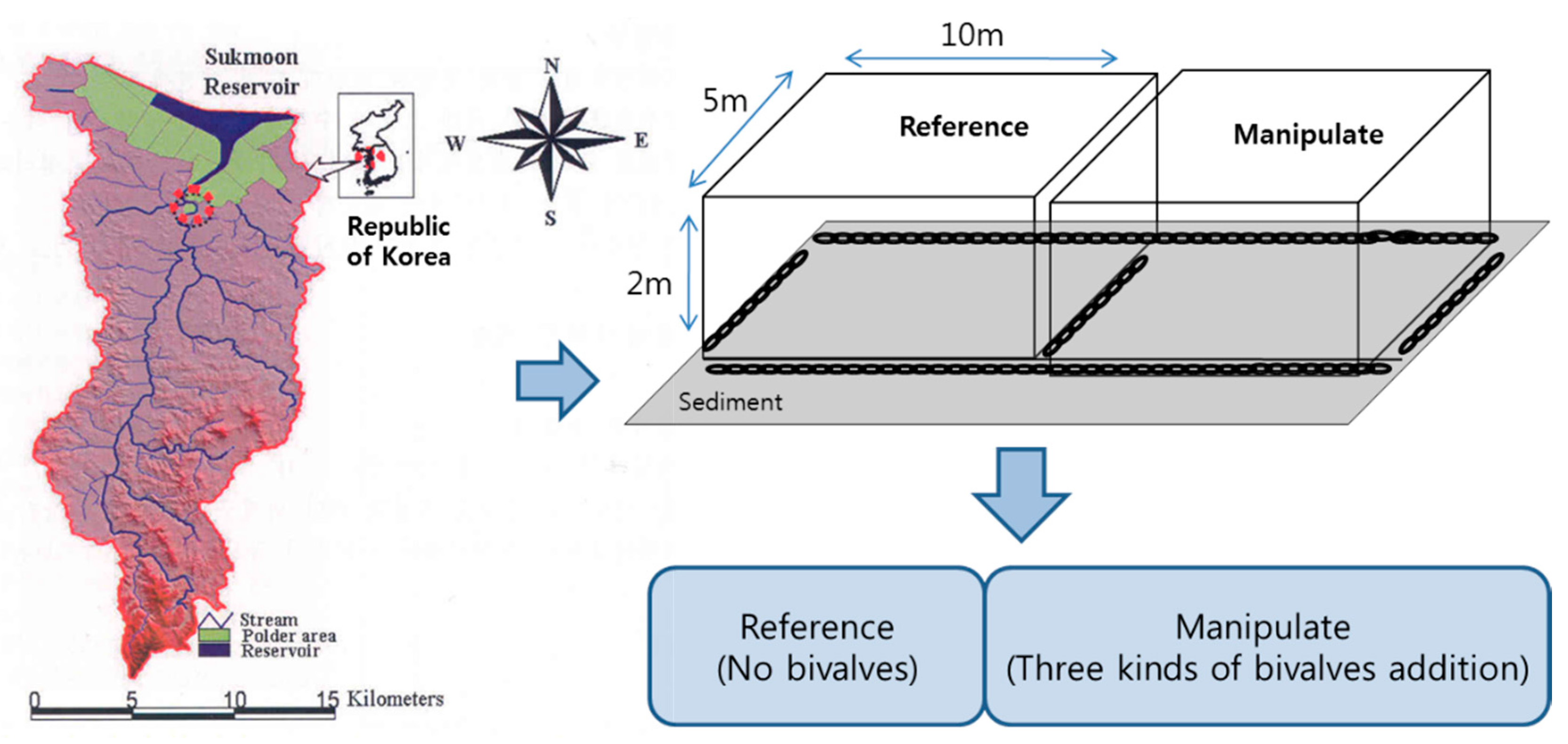
2.1. Characteristics of Ponds
- (1)
- Reference Pond (RP; in situ water containing predominantly M. aeruginosa).
- (2)
- Manipulated Pond with bivalves (MP; in situ water containing predominantly M. aeruginosa + bivalves (U. douglasiae, S. woodiana and S. arcaeformis)).
2.2. In Situ Artificial Pond Experiment
2.3. Analysis of Water Quality Parameters
2.4. Enumeration and Biomass Determination of Phytoplankton
2.5. Analysis of Stable Isotope Ratios
2.6. Determination of MCs Concentration and Production Rate
2.7. Statistical Analysis
3. Results
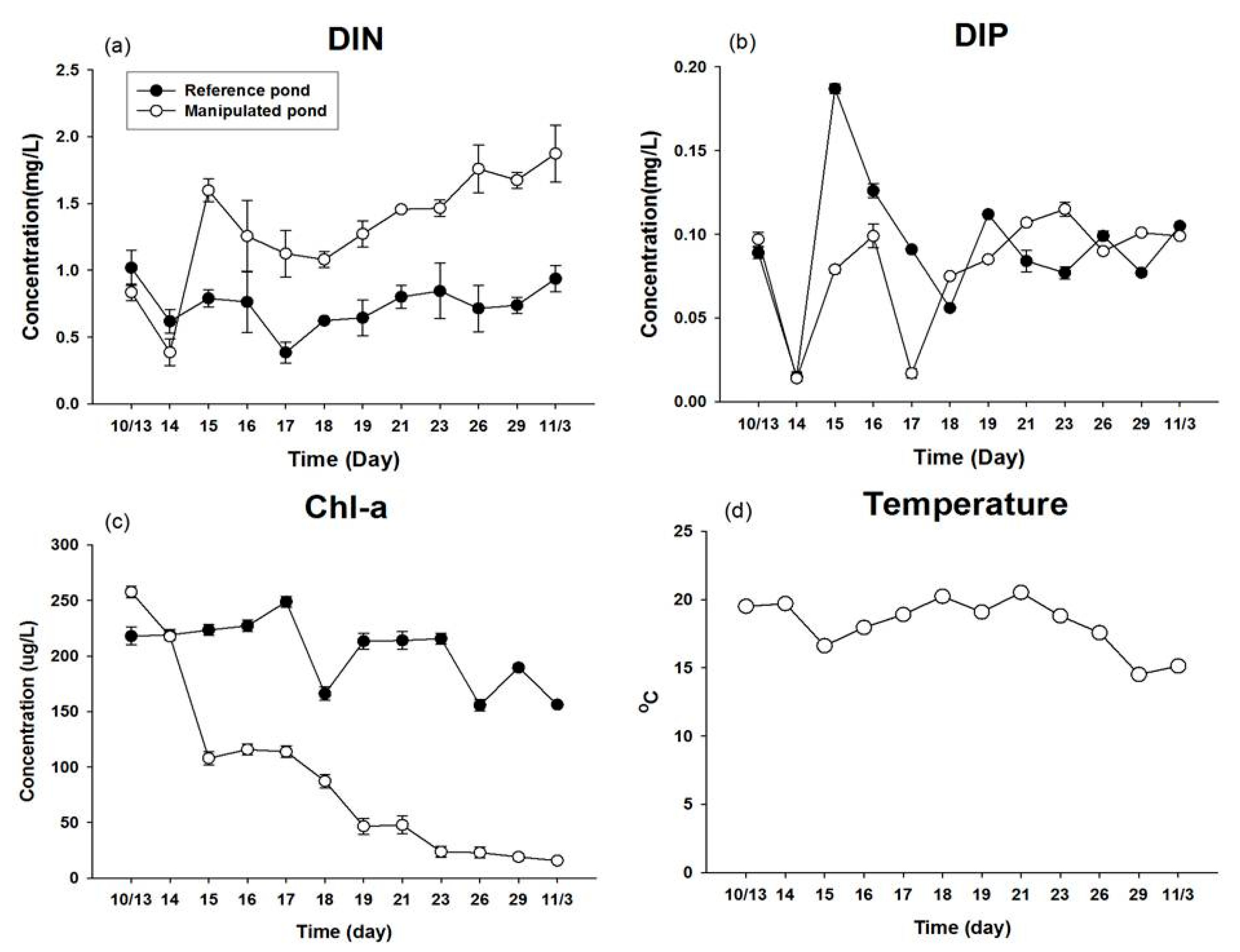
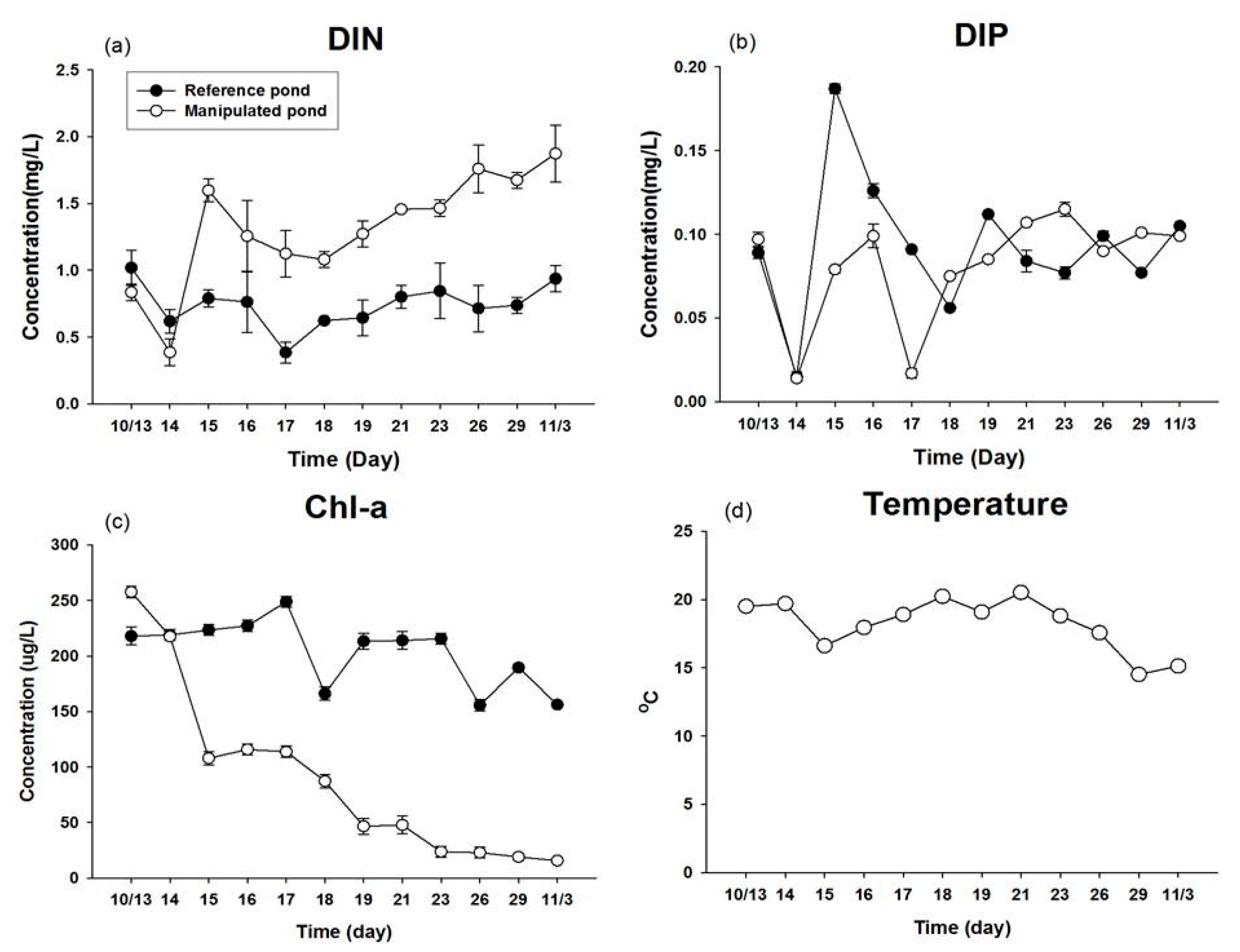
3.1. Water Quality Condition in an Artificial Enclosed Pond
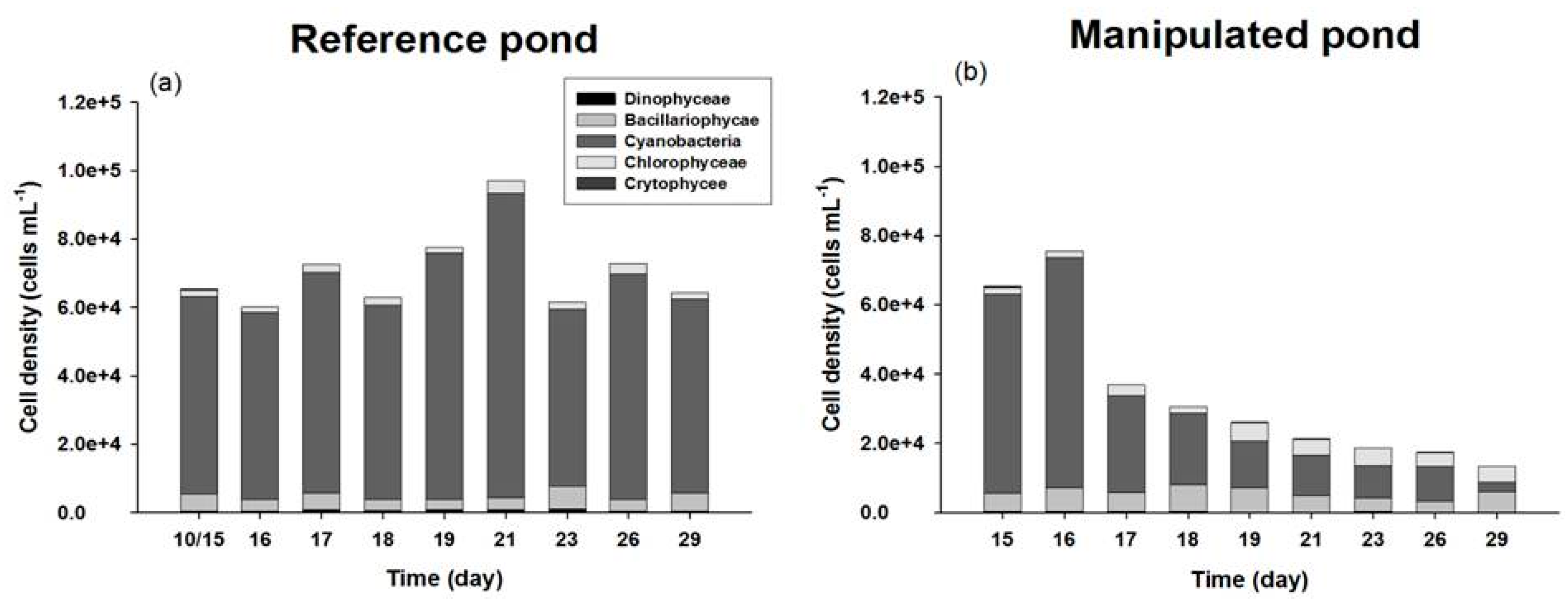
3.2. The Composition and Biomass Variation of Phytoplankton
3.3. Measurement of 13C and 15N Atom % of Filter Feeders
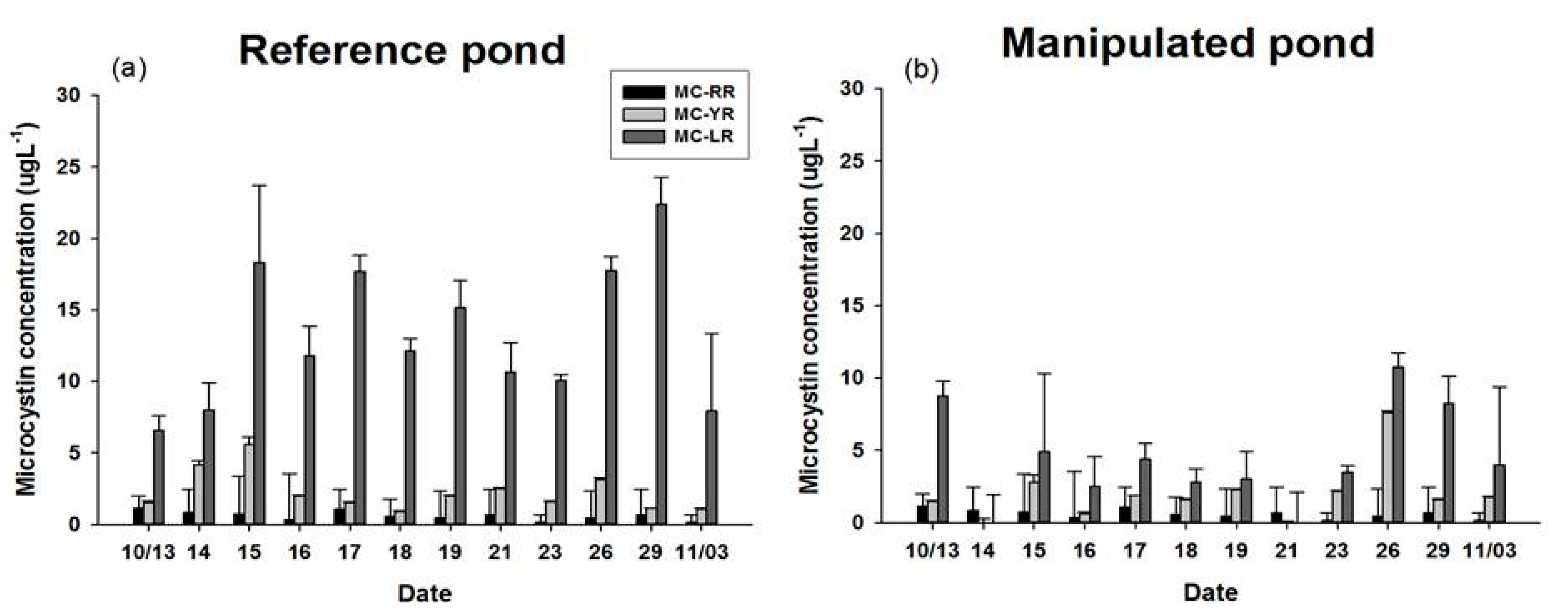
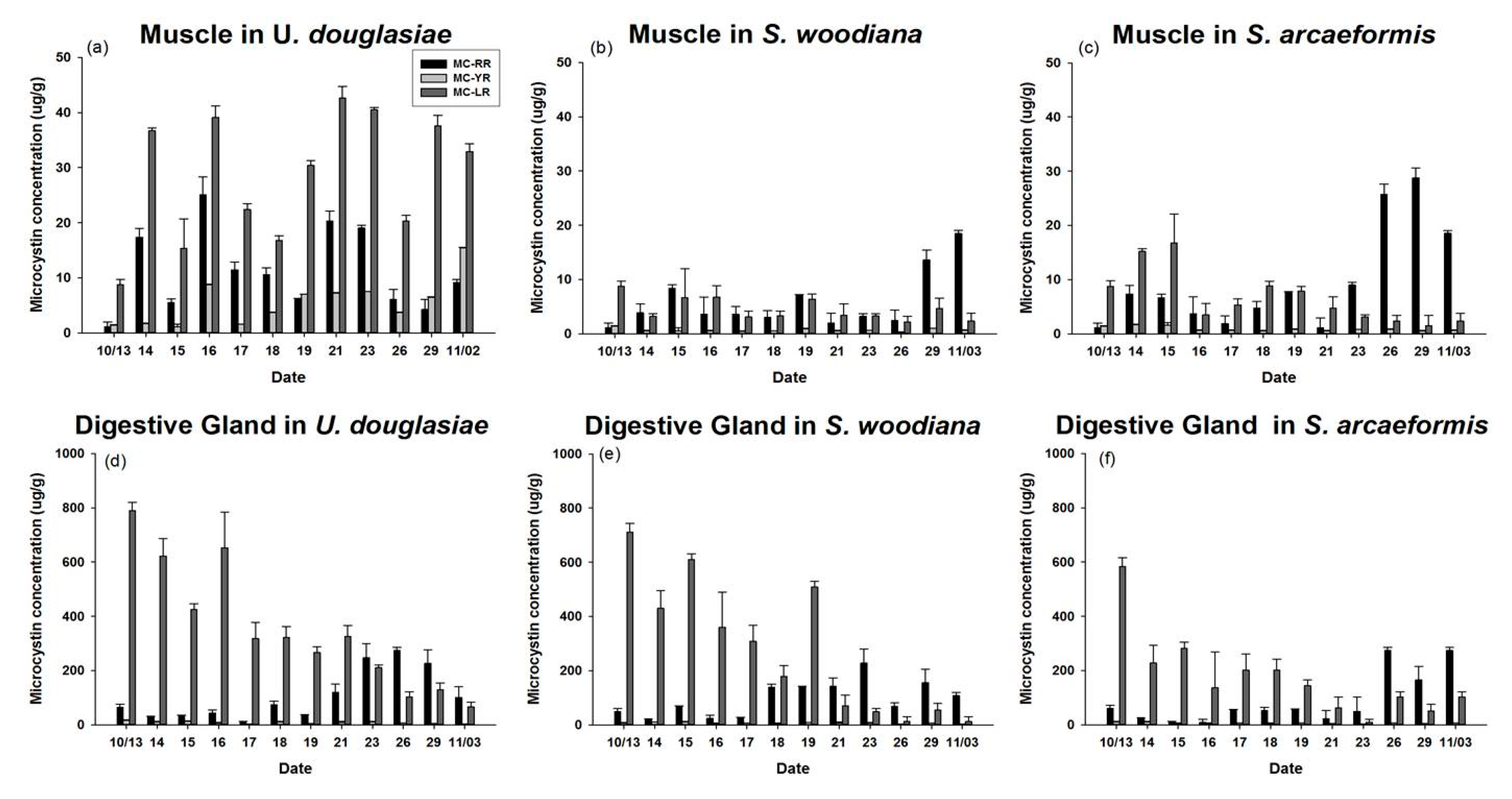
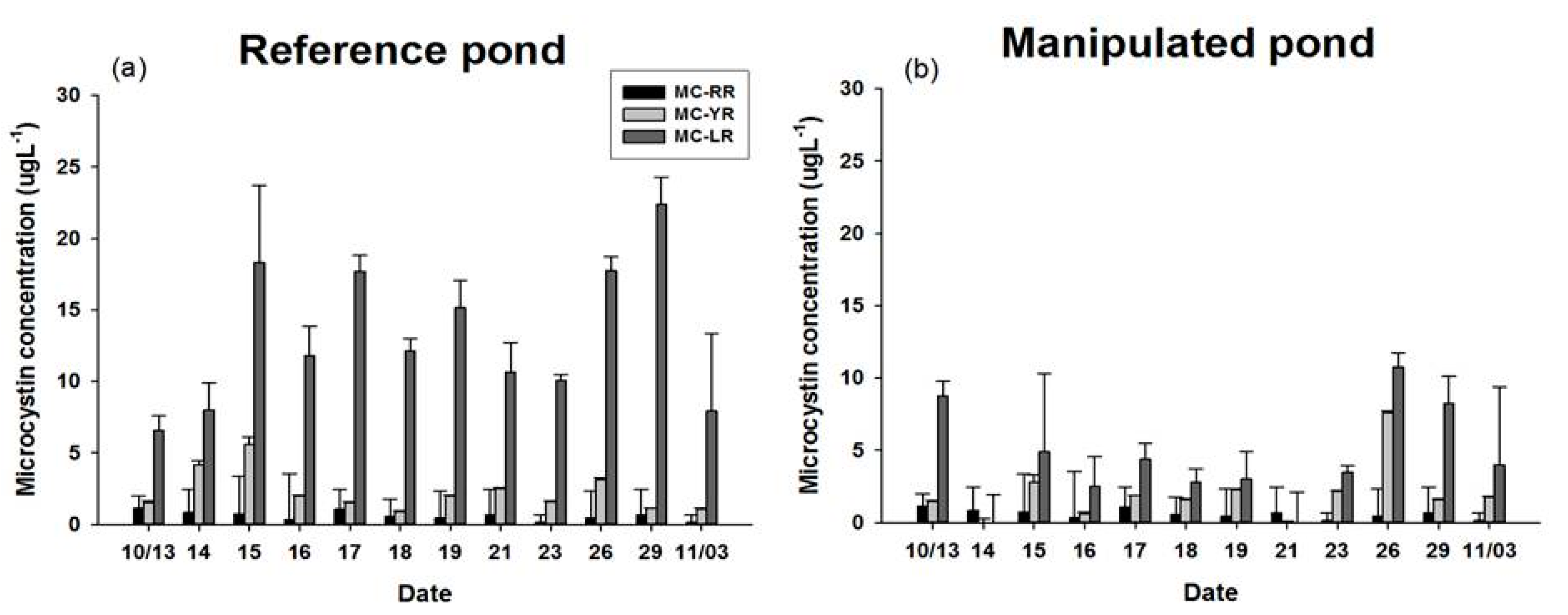
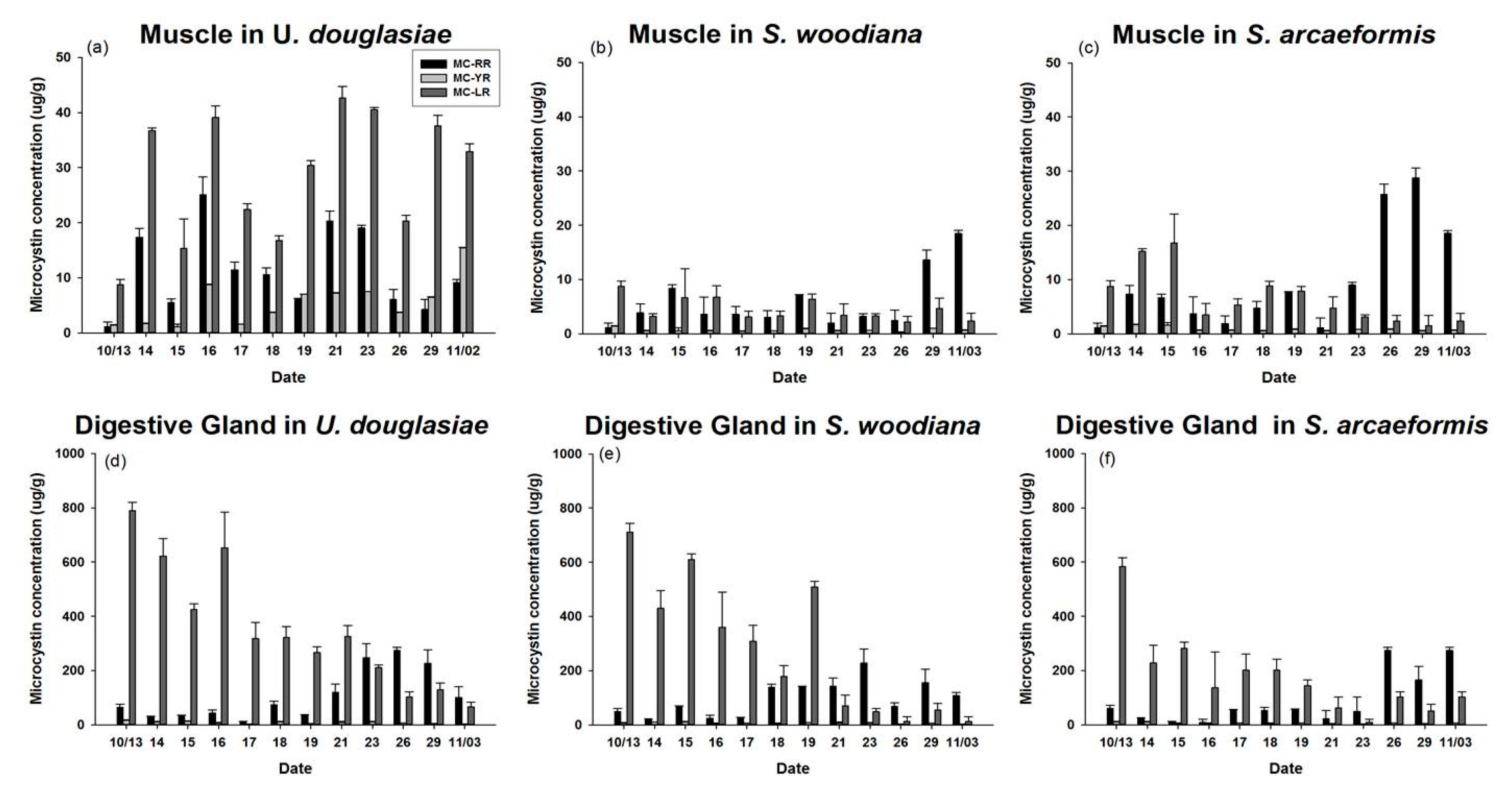
3.4. MC Concentration of POM and Three Kinds of Bivalves
4. Discussion
4.1. Dynamics of Feeding Activity and MCs Concentration in Filter Feeders
4.2. Stable Isotope Fractionation in Differential Organs
4.3. Management and Implications for Water Quality
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bekli̇oğlu, M. A review on the control of eutrophication in deep and shallow lakes. Turk. J. Zool. 1999, 23, 327–336. [Google Scholar]
- Chorus, E.I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E&FN Spon: London, UK, 1999; Chapter 8; pp. 235–273. [Google Scholar]
- De Figueiredo, D.R.; Azeiteiro, U.M.; Esteves, S.M.; Gonçalves, F.J.; Pereira, M.J. Microcystin-producing blooms—A serious global public health issue. Ecotoxicol. Environ. Saf. 2004, 59, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Seikai, T.; Tominaga, O. Estimation of growth and food consumption in juvenile Japanese flounder Paralichthys olivaceus using carbon stable isotope ratio δ13C under laboratory conditions. J. Exp. Mar. Biol. Ecol. 2005, 326, 187–198. [Google Scholar] [CrossRef]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Boon, P.I.; Bunn, S.E.; Green, J.D.; Shiel, R.J. Consumption of cyanobacteria by freshwater zooplankton: Implications for the success of ‘top-down’ control of cyanobacterial blooms in Australia. Mar. Freshw. Res. 1994, 45, 875–887. [Google Scholar] [CrossRef]
- Koski, M.; Engström, J.; Viitasalo, M. Reproduction and survival of the calanoid copepod Eurytemora affinis fed with toxic and non-toxic cyanobacteria. Mar. Ecol. Prog. Ser. 1999, 186, 187–197. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, Y.; Hong, S.; Hwang, S.J.; Kim, B.H.; An, K.G.; Park, Y.S.; Park, S.G.; Um, H.Y.; Shin, S.H. Effects of biocontrol with an atyid shrimp (Caridina deniculata) and a bagrid catfish (Pseudobagrus fulvidraco) on toxic cyanobacteria bloom (Microcystis aeruginosa) in a eutrophic agricultural reservoir. Paddy Water Environ. 2017, 15, 483–497. [Google Scholar] [CrossRef]
- Bontes, B.M.; Verschoor, A.M.; Pires, L.M.D.; van Donk, E.; Ibelings, B.W. Functional response of anodonta anatina feeding on a green alga and four strains of cyanobacteria, differing in shape, size and toxicity. Hydrobiologia 2007, 584, 191–204. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Carmichael, W.W.; Hussein, A.A. Estimation of microcystins in the freshwater fish oreochromis niloticus in an egyptian fish farm containing a microcystis bloom. Environ. Toxicol. 2003, 18, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Hosper, S.; Jagtman, E. Biomanipulation additional to nutrient control for restoration of shallow lakes in the netherlands. In Biomanipulation Tool for Water Management; Springer: Heidelberg, Germany, 1990; pp. 523–534. [Google Scholar]
- Meijer, M.-L. Biomanipulation in The Netherlands—15 Years of Experience. Ph.D. Thesis, Wageningen Agricultural University, Wageningen, The Netherlands, May 2000. [Google Scholar]
- Welker, M.; Walz, N. Can mussels control the plankton in rivers?—A planktological approach applying a lagrangian sampling strategy. Limnol. Oceanogr. 1998, 43, 753–762. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Liebig, J.R.; Carmichael, W.W.; Agy, M.A.; Johengen, T.H.; Fahnenstiel, G.L.; Nalepa, T.F. Zebra mussel (Dreissena polymorpha) selective filtration promoted toxic microcystis blooms in saginaw bay (lake huron) and lake erie. Can. J. Fish. Aquat. Sci. 2001, 58, 1208–1221. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Nalepa, T.F.; Jude, D.J.; Mills, E.L.; Holeck, K.T.; Liebig, J.R.; Grigorovich, I.A.; Ojaveer, H. Dispersal and emerging ecological impacts of ponto-caspian species in the laurentian great lakes. Can. J. Fish. Aquat. Sci. 2002, 59, 1209–1228. [Google Scholar] [CrossRef]
- Reeders, H.; Vaate, A.; Slim, F. The fitration rate of Dreissena polymorpha (bivalvia) in three dutch lakes with reference to biological water quality management. Freshw. Biol. 1989, 22, 133–141. [Google Scholar] [CrossRef]
- Pires, L.D.; Bontes, B.M.; Van Donk, E.; Ibelings, B.W. Grazing on colonial and filamentous, toxic and non-toxic cyanobacteria by the zebra mussel Dreissena polymorpha. J. Plankton Res. 2005, 27, 331–339. [Google Scholar] [CrossRef]
- Byllaardt, J.V.; Ackerman, J.V. Hydrodynamic habitat influences suspension feeding by unionid mussels in freshwater ecosystem. Freshw. Biol. 2014, 59, 1187–1196. [Google Scholar] [CrossRef]
- Atkinson, C.L.; First, M.R.; Covich, A.P.; Opsahl, S.P.; Golladay, S.W. Suspended material availability and filtration-biodeposition processes performed by a native and invasive bivalve species in streams. Hydrobiologia 2011, 667, 191–204. [Google Scholar] [CrossRef]
- Reichwaldt, E.S.; Ghadouani, A. Can mussels be used as sentinel organisms for characterization of pollution in urban water systems? Hydrol. Earth Syst. Sci. 2016, 20, 2679–2689. [Google Scholar] [CrossRef]
- Reeders, H.; De Vaate, A.B. Zebra mussels (Dreissena polymorpha): A new perspective for water quality management. In Biomanipulation Tool for Water Management; Springer: Heidelberg, Germany, 1990; pp. 437–450. [Google Scholar]
- Soto, D.; Mena, G. Filter feeding by the freshwater mussel, diplodon chilensis, as a biocontrol of salmon farming eutrophication. Aquaculture 1999, 171, 65–81. [Google Scholar] [CrossRef]
- Horgan, M.J.; Mills, E.L. Zebra mussel filter feeding and food-limited production of daphnia: Recent changes in lower trophic level dynamics of oneida lake, New York, USA. Hydrobiologia 1999, 411, 79–88. [Google Scholar] [CrossRef]
- Ricciardi, A.; Neves, R.J.; Rasmussen, J.B. Impending extinctions of north american freshwater mussels (unionoida) following the zebra mussel (Dreissena polymorpha) invasion. J. Anim. Ecol. 1998, 67, 613–619. [Google Scholar] [CrossRef]
- Watters, G.T. A synthesis and review of the expanding range of the asian freshwater mussel anodonta woodiana (Lea, 1834) (Bivalvia, Unionidae). Veliger 1997, 40, 152–156. [Google Scholar]
- Prins, T.C.; Smaal, A.C.; Dame, R.F. A review of the feedbacks between bivalve grazing and ecosystem processes. Aquat. Ecol. 1997, 31, 349–359. [Google Scholar] [CrossRef]
- Mills, S.C.; Reynolds, J.D. Host species preferences by bitterling, rhodeus sericeus, spawning in freshwater mussels and consequences for offspring survival. Anim. Behav. 2002, 63, 1029–1036. [Google Scholar] [CrossRef]
- Reynolds, J.D.; Guillaume, H.P. Effects of phosphate on the reproductive symbiosis between bitterling and freshwater mussels: Implications for conservation. J. Appl. Ecol. 1998, 35, 575–581. [Google Scholar] [CrossRef]
- Fry, B. Stable isotope diagrams of freshwater food webs. Ecology 1991, 72, 2293–2297. [Google Scholar] [CrossRef]
- Witte, U.; Wenzhöfer, F.; Sommer, S.; Boetius, A.; Heinz, P.; Aberle, N.; Sand, M.; Cremer, A.; Abraham, W.-R.; Jørgensen, B. in situ experimental evidence of the fate of a phytodetritus pulse at the abyssal sea floor. Nature 2003, 424, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Parkyn, S.M.; Quinn, J.M.; Cox, T.J.; Broekhuizen, N. Pathways of n and c uptake and transfer in stream food webs: An isotope enrichment experiment. J. N. Am. Benthol. Soc. 2005, 24, 955–975. [Google Scholar] [CrossRef]
- Kim, B.-H.; Lee, J.-H.; Hwang, S.-J. Inter- and intra-specific differences in filtering activities between two unionids, Anodonta woodiana and Unio douglasiae, in ambient eutrophic lake waters. Ecol. Eng. 2011, 37, 1957–1967. [Google Scholar] [CrossRef]
- Strickland, J.D.; Parsons, T.R. A Practical Handbook of Seawater Analysis; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1968; p. 311. [Google Scholar]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses, 2nd ed.; Springer: New York, NY, USA, 1991. [Google Scholar]
- Kim, M.S.; Hwang, J.Y.; Kwon, O.S.; Lee, W.S. Analytical methodology of stable isotopes ratios: Sample pretreatment, analysis and application. Korean J. Envion. Ecol. 2013, 46, 471–487. [Google Scholar]
- Kim, M.S.; Lee, W.W.; Suresh Kumar, S.; Shin, S.H.; Robarge, W.; Kim, M.; Lee, S.R. Effects of HCl pretreatment, drying, and storage on the stable isotope ratios of soil and sediment samples. Rapid Commun. Mass Spectrom. 2016, 30, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.-I.; Matsuura, K.; Suzuki, M.; Oka, H.; Watanabe, M.F.; Oishi, S.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W. Analysis and purification of toxic peptides from cyanobacteria by reversed-phase high-performance liquid chromatography. J. Chromatogr. A 1988, 448, 275–283. [Google Scholar] [CrossRef]
- Vaughn, C.C.; Gido, K.B.; Spooner, D.E. Ecosystem processes performed by unionid mussels in stream mesocosms: Species roles and effects of abundance. Hydrobiologia 2004, 527, 35–47. [Google Scholar] [CrossRef]
- Yokoyama, A.; Park, H.D. Mechanism and prediction for contamination of freshwater bivalves (Unionidae) with the cyanobacterial toxin microcystin in hypereutrophic Lake Suwa, Japan. Environ. Toxicol. 2002, 17, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.E.; Meriluoto, J.A.; Lindholm, T. Accumulation of a peptide toxin from the cyanobacterium oscillatoria agardhii in the freshwater mussel anadonta cygnea. Hydrobiologia 1989, 183, 211–216. [Google Scholar] [CrossRef]
- Vasconcelos, V.M. Uptake and depuration of the heptapeptide toxin microcystin-lr in mytilus galloprovincialis. Aquat. Toxicol. 1995, 32, 227–237. [Google Scholar] [CrossRef]
- Yokoyama, A.; Park, H.D. Depuration kinetics and persistence of the cyanobacterial toxin microcystin-lr in the freshwater bivalve unio douglasiae. Environ. Toxicol. 2003, 18, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.F.; Park, H.D.; Kondo, F.; Harada, K.; Hayashi, H.; Okino, T. Identification and estimation of microcystins in freshwater mussels. Nat. Toxins 1997, 5, 31–35. [Google Scholar] [CrossRef]
- Chen, J.; Xie, P. Seasonal dynamics of the hepatotoxic microcystins in various organs of four freshwater bivalves from the large eutrophic lake taihu of subtropical China and the risk to human consumption. Environ. Toxicol. 2005, 20, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, V.; Oliveira, S.; Teles, F.O. Impact of a toxic and a non-toxic strain of microcystis aeruginosa on the crayfish procambarus clarkii. Toxicon 2001, 39, 1461–1470. [Google Scholar] [CrossRef]
- Wiegand, C.; Pflugmacher, S.; Oberemm, A.; Meems, N.; Beattie, K.A.; Steinberg, C.E.; Codd, G.A. Uptake and effects of microcystin-lr on detoxication enzymes of early life stages of the zebra fish (Danio rerio). Environ. Toxicol. 1999, 14, 89–95. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Wiegand, C.; Oberemm, A.; Beattie, K.A.; Krause, E.; Codd, G.A.; Steinberg, C.E. Identification of an enzymatically formed glutathione conjugate of the cyanobacterial hepatotoxin microcystin-lr: The first step of detoxication. Biochim. Biophys. Acta 1998, 1425, 527–533. [Google Scholar] [CrossRef]
- Kondo, F.; Ikai, Y.; Oka, H.; Okumura, M.; Ishikawa, N.; Harada, K.; Matsuura, K.; Murata, H.; Suzuki, M. Formation, characterization, and toxicity of the glutathione and cysteine conjugates of toxic heptapeptide microcystins. Chem. Res. Toxicol. 1992, 5, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Lorrain, A.; Paulet, Y.-M.; Chauvaud, L.; Savoye, N.; Donval, A.; Saout, C. Differential δ13C and δ15N signatures among scallop tissues: Implications for ecology and physiology. J. Exp. Mar. Biol. Ecol. 2002, 275, 47–61. [Google Scholar] [CrossRef]
- Deudero, S.; Box, A.; Tejada, S.; Tintoré, J. Stable isotopes and metal contamination in caged marine mussel mytilus galloprovincialis. Mar. Pollut. Bull. 2009, 58, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Raikow, D.F.; Hamilton, S.K. Bivalve diets in a midwestern us stream: A stable isotope enrichment study. Limnol. Oceanogr. 2001, 46, 514–522. [Google Scholar] [CrossRef]
- Tieszen, L.L.; Boutton, T.W.; Tesdahl, K.G.; Slade, N.A. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia 1983, 57, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.M.; McQuaid, C.D. Effects of food quality on tissue-specific isotope ratios in the mussel perna perna. Hydrobiologia 2009, 635, 81–94. [Google Scholar] [CrossRef]
- Baker, S.M.; Levinton, J.S.; Kurdziel, J.P.; Shumway, S.E. Selective feeding and biodeposition by zebra mussels and their relation to changes in phytoplankton composition and seston load. J. Shellfish Res. 1998, 17, 1207–1214. [Google Scholar]
- Ibelings, B.W.; Vonk, M.; Los, H.F.; van der Molen, D.T.; Mooij, W.M. Fuzzy modeling of cyanobacterial surface waterblooms: Validation with noaa-avhrr satellite images. Ecol. Appl. 2003, 13, 1456–1472. [Google Scholar] [CrossRef]
- Reeders, H.; bij de Vatte, A.; Noordhuis, R. Potential of the zebra mussel (Dreissena polymorpha) for water quality management. In Zebra Mussels: Biology, Impacts, and Control; Nalepa, T.F., Schloesser, D.W., Eds.; Lewis Publishers/CRC Press: Boca Raton, FL, USA, 1993; pp. 439–451. [Google Scholar]
- Horgan, M.J.; Mills, E.L. Clearance rates and filtering activity of zebra mussel (Dreissena polymorpha): Implications for freshwater lakes. Can. J. Fish. Aquat. Sci. 1997, 54, 249–255. [Google Scholar] [CrossRef]
- Naddafi, R.; Pettersson, K.; Eklöv, P. The effect of seasonal variation in selective feeding by zebra mussels (Dreissena polymorpha) on phytoplankton community composition. Freshw. Biol. 2007, 52, 823–842. [Google Scholar] [CrossRef]
- Xie, L.; Xie, P.; Guo, L.; Li, L.; Miyabara, Y.; Park, H.D. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic Lake Chaohu, China. Environ. Toxicol. 2005, 20, 293–300. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters (Unit) | p Value |
|---|---|
| Water temperature (°C) | 0.962 |
| Dissolved inorganic nitrogen (mg L−1) | 0.000 * |
| Dissolved inorganic phosphorus (mg L−1) | 0.364 |
| Chl-a (μg L−1) | 0.000 * |
| Phytoplankton cell density (cells mL−1) | 0.005 * |
| Class | Species | Cell Density (Cells mL−1) | Relative Proportion (%) |
|---|---|---|---|
| Cyanobacteria | 87.84 | ||
| Microcystis aeruginosa | 43,600 | ||
| Anabaena spiroides | 8560 | ||
| Synechocysits pevalekii | 2240 | ||
| Aphanocapsa elachista | 1548 | ||
| Chroococcus sp. | 1600 | 2.99 | |
| Chlorophyceae | |||
| Scenedesmus sp. | 1280 | ||
| Pediastrum boryanum | 0 | ||
| Pediastrum simplex | 0 | 0.73 | |
| Staurastrum sp. | 320 | 0.54 | |
| Monoraphidium contortum | 360 | ||
| Crytophyceae | |||
| Cryptomonas erosa | 480 | 7.87 | |
| Dinophyceae | |||
| Ceratium hirundinella | 360 | ||
| Euglena sp. | 0 | ||
| Bacillariophyceae | |||
| Synedra acus | 440 | ||
| Synedra sp. | 160 | ||
| Navicula sp. | 200 | ||
| Aulacoseira granulata | 2280 | ||
| Aulacoseira sp. | 240 | ||
| Nitzschia holsatica | 400 | ||
| Cyclotella comta | 1440 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.-S.; Lee, Y.-J.; Ha, S.-Y.; Kim, B.-H.; Hwang, S.-J.; Kwon, J.-T.; Choi, J.-W.; Shin, K.-H. Accumulation of Microcystin (LR, RR and YR) in Three Freshwater Bivalves in Microcystis aeruginosa Bloom Using Dual Isotope Tracer. Mar. Drugs 2017, 15, 226. https://doi.org/10.3390/md15070226
Kim M-S, Lee Y-J, Ha S-Y, Kim B-H, Hwang S-J, Kwon J-T, Choi J-W, Shin K-H. Accumulation of Microcystin (LR, RR and YR) in Three Freshwater Bivalves in Microcystis aeruginosa Bloom Using Dual Isotope Tracer. Marine Drugs. 2017; 15(7):226. https://doi.org/10.3390/md15070226
Chicago/Turabian StyleKim, Min-Seob, Yeon-Jung Lee, Sun-Yong Ha, Baik-Ho Kim, Soon-Jin Hwang, Jung-Taek Kwon, Jong-Woo Choi, and Kyung-Hoon Shin. 2017. "Accumulation of Microcystin (LR, RR and YR) in Three Freshwater Bivalves in Microcystis aeruginosa Bloom Using Dual Isotope Tracer" Marine Drugs 15, no. 7: 226. https://doi.org/10.3390/md15070226
APA StyleKim, M.-S., Lee, Y.-J., Ha, S.-Y., Kim, B.-H., Hwang, S.-J., Kwon, J.-T., Choi, J.-W., & Shin, K.-H. (2017). Accumulation of Microcystin (LR, RR and YR) in Three Freshwater Bivalves in Microcystis aeruginosa Bloom Using Dual Isotope Tracer. Marine Drugs, 15(7), 226. https://doi.org/10.3390/md15070226