Functional Comparison for Lipid Metabolism and Intestinal and Fecal Microflora Enzyme Activities between Low Molecular Weight Chitosan and Chitosan Oligosaccharide in High-Fat-Diet-Fed Rats



Abstract
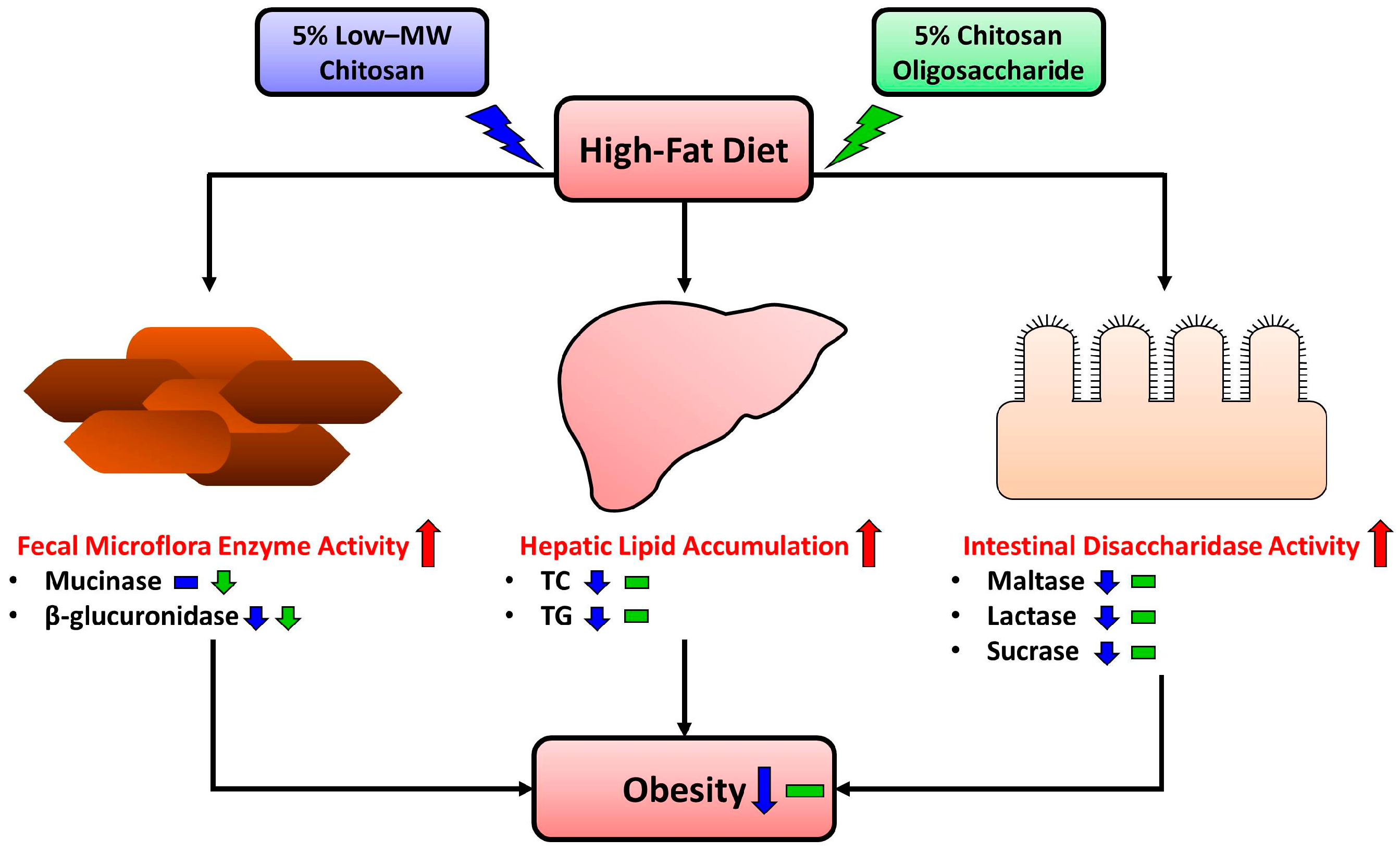
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Diets
2.3. Determination of Triglyceride (TG) and Total Cholesterol (TC)
2.4. Determination of Plasma Tumor Necrosis Factor-α (TNF-α)
2.5. Determination of Hepatic Fatty Acid Synthetase (FAS) Activity
2.6. Determination of Small Intestinal Disaccharidase Activity
2.7. Determination of Fecal Mucinase and β-Glucuronidase Activities
2.8. Statistical Evaluation
3. Results and Discussion
3.1. Effects of Both LC and CO on Body Weight, Organ and Tissue Weights, Food Intake, and Feed Efficiency in HF-Diet-Fed Rats
3.2. Effects of Both LC and CO on Hepatic and Fecal Lipid Responses and Lipid-Related Metabolic Changes in HF-Diet-Fed Rats
3.3. Effects of Both LC and CO on Intestinal Disaccharidase Activities and Fecal Bacterial Enzyme Activities in HF-Diet-Fed Rats
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Kearney, J. Food consumption trends and drivers. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2010, 365, 2793–2807. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity and Overweight. 2016. Available online: http://www.who.int/mediacentre/factsheets/fs311/en/ (accessed on 17 February 2017).
- World Health Organization. Diabetes. 2016. Available online: http://www.who.int/mediacentre/factsheets/fs312/en/ (accessed on 17 February 2017).
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Field, A.E.; Coakley, E.H.; Must, A.; Spadano, J.L.; Laird, N.; Dietz, W.H.; Rimm, E.; Colditz, G.A. Impact of overweight on the risk of developing common chronic diseases during a 10-year period. Arch. Intern. Med. 2001, 161, 1581–1586. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S. Functional foods against metabolic syndrome (obesity, diabetes, hypertension and dyslipidemia) and cardiovascular disease. Trends Food Sci. Technol. 2014, 35, 114–128. [Google Scholar] [CrossRef]
- Deuchi, K.; Kanauchi, O.; Imasato, Y.; Kobayashi, E. Effect of the viscosity or deacetylation degree of chitosan on fecal fat excreted from rats fed on a high fat diet. Biosci. Biotechnol. Biochem. 1995, 59, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.L.; Yao, H.T.; Cheng, R.S.; Chiang, M.T. Chitosan reduces plasma adipocytokines and lipid accumulation in liver and adipose tissues and ameliorates insulin resistance in diabetic rats. J. Med. Food 2012, 15, 453–460. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Li, L.; Xia, W. Dietary chitosan improves hypercholesterolemia in rats fed high-fat diets. Nutr. Res. 2008, 28, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Cherniak, E.P. Potential applications for alternative medicine to treat obesity in an aging population. Altern. Med. Rev. 2008, 13, 34–42. [Google Scholar]
- Mhurchu, C.N.; Poppitt, S.D.; McGill, A.T.; Leahy, F.E.; Bennett, D.A.; Lin, R.B.; Ormrod, D.; Ward, L.; Strik, C.; Rodgers, A. The effect of the dietary supplement, Chitosan, on body weight: A randomised controlled trial in 250 overweight and obese adults. Int. J. Obes. Relat. Metab. Disord. 2004, 28, 1149–1456. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, V.R.; Satia, M.C.; Deschamps, A.; Maquet, V.; Shah, R.B.; Zinzuwadia, P.H.; Trivedi, J.V. Single-blind, placebo controlled randomised clinical study of chitosan for body weight reduction. Nutr. J. 2016, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Schiller, R.N.; Barrager, E.; Schauss, A.G.; Nichols, E.J. A randomized, double-blind, placebo-controlled study examining the effects of a rapidly soluble chitosan dietary supplement on weight loss and body composition in overweight and mildly obese individuals. J. Am. Nutraceutical Assoc. 2001, 4, 42–49. [Google Scholar]
- Ho, S.C.; Tai, E.S.; Eng, P.H.; Tan, C.E.; Fok, A.C. In the absence of dietary surveillance chitosan does not reduce plasma lipids or obesity in hypercholesterolaemic obese Asian subjects. Singap. Med. J. 2001, 42, 6–10. [Google Scholar]
- Huang, L.; Chen, J.; Cao, P.; Pan, H.; Ding, C.; Xiao, T.; Zhang, P.; Guo, J.; Su, Z. Anti-obese effect of glucosamine and chitosan oligosaccharide in high-fat diet-induced obese rats. Mar. Drugs 2015, 13, 2732–2756. [Google Scholar] [CrossRef] [PubMed]
- Sumiyoshi, M.; Kimura, Y. Low molecular weight chitosan inhibits obesity induced by feeding a high-fat diet long-term in mice. J. Pharm. Pharmacol. 2006, 58, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Shiau, S.Y.; Chang, G.W. Effects of dietary fiber on fecal mucinase and beta-glucuronidase activity in rats. J. Nutr. 1983, 113, 138–144. [Google Scholar] [PubMed]
- Institute of Laboratory Animal Resources. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Takehisa, F.; Suzuki, Y. Effect of guar gum and cholestyramine on plasma lipoprotein cholesterol in rats. J. Jpn. Soc. Nutr. Food Sci. 1990, 43, 269–274. [Google Scholar] [CrossRef]
- Nepokroeff, C.M.; Lakshmanan, M.R.; Porter, J.W. Fatty-acid synthase from rat liver. Methods Enzymol. 1975, 35, 37–44. [Google Scholar] [PubMed]
- Yao, H.T.; Huang, S.Y.; Chiang, M.T. A comparative study on hypoglycemic and hypocholesterolemic effects of high and low molecular weight chitosan in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2008, 46, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Dahlqvist, A. Assay of intestinal disaccharidases. Anal. Biochem. 1968, 22, 99–107. [Google Scholar] [CrossRef]
- Yao, H.T.; Chiang, M.T. Chitosan shifts the fermentation site toward the distal colon and increases the fecal short-chain fatty acids concentrations in rats. Int. J. Vitam. Nutr. Res. 2006, 76, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Ito, M. Antidiabetic action of low molecular weight chitosan in genetically obese diabetic KK-Ay mice. Biol. Pharm. Bull. 2002, 25, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.H.; Yang, H.P.; Chun, H.S. Chitooligosaccharide ameliorates diet-induced obesity in mice and affects adipose gene expression involved in adipogenesis and inflammation. Nutr. Res. 2012, 32, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Jo, S.H.; Ha, K.S.; Kim, S.C.; Kim, Y.C.; Apostolidis, E.; Kwon, Y. Effect of long-term supplementation of low molecular weight chitosan oligosaccharide (GO2KA1) on fasting blood glucose and HbA1c in db/db mice model and elucidation of mechanism of action. BMC Complement. Altern. Med. 2014, 14, 272. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.H.; Ha, K.S.; Moon, K.S.; Kim, J.G.; Oh, C.G.; Kim, Y.C.; Apostolidis, E.; Kwon, Y.I. Molecular weight dependent glucose lowering effect of low molecular weight Chitosan Oligosaccharide (GO2KA1) on postprandial blood glucose level in SD rats model. Int. J. Mol. Sci. 2013, 14, 14214–14224. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Chan, I.L.; Yang, T.H.; Liu, S.H.; Chiang, M.T. Supplementation of chitosan alleviates high-fat diet-enhanced lipogenesis in rats via adenosine monophosphate (AMP)-activated protein kinase activation and inhibition of lipogenesis-associated genes. J. Agric. Food Chem. 2015, 63, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Chang, T.C.; Liu, S.H.; Chiang, M.T. The regulatory effects of fish oil and chitosan on hepatic lipogenic signals in high-fat diet-induced obese rats. J. Food Drug Anal. 2017. [Google Scholar] [CrossRef]
- Guo, J.; Jou, W.; Gavrilova, O.; Hall, K.D. Persistent diet-induced obesity in male C57BL/6 mice resulting from temporary obesigenic diets. PLoS ONE 2009, 4, e5370. [Google Scholar] [CrossRef] [PubMed]
- Biddinger, S.B.; Almind, K.; Miyazaki, M.; Kokkotou, E.; Ntambi, J.M.; Kahn, C.R. Effects of diet and genetic background on sterol regulatory element-binding protein-1c, stearoyl-CoA desaturase 1, and the development of the metabolic syndrome. Diabetes 2005, 54, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Meugnier, E.; Bossu, C.; Oliel, M.; Jeanne, S.; Michaut, A.; Sothier, M.; Brozek, J.; Rome, S.; Laville, M.; Vidal, H. Changes in gene expression in skeletal muscle in response to fat overfeeding in lean men. Obesity 2007, 15, 2583–2594. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Vazquez, I.; Fernández-Veledo, S.; Krämer, D.K.; Vila-Bedmar, R.; Garcia-Guerra, L.; Lorenzo, M. Insulin resistance associated to obesity: The link TNF-α. Arch. Physiol. Biochem. 2008, 114, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xun, K.; Chen, L.; Wang, Y. TNF-α, a potent lipid metabolism regulator. Cell. Biochem. Funct. 2009, 27, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R.; Soued, M.; Staprans, I.; Gavin, L.A.; Donahue, M.E.; Huang, B.J.; Moser, A.H.; Gulli, R.; Grunfeld, C. Effect of tumor necrosis factor (TNF) on lipid metabolism in the diabetic rat. Evidence that inhibition of adipose tissue lipoprotein lipase activity is not required for TNF-induced hyperlipidemia. J. Clin. Investig. 1989, 83, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.P.; Yao, H.T.; Chiang, M.T. Effects of high and low molecular weight chitosan on plasma cholesterol, glucose and adipocytokines in diabetic rats induced by streptozotocin and nicotinamide. J. Food Drug Anal. 2012, 20, 661–667. [Google Scholar]
- Kumar, S.G.; Rahman, M.A.; Lee, S.H.; Hwang, H.S.; Kim, H.A.; Yun, J.W. Plasma proteome analysis for anti-obesity and anti-diabetic potentials of chitosan oligosaccharides in ob/ob mice. Proteomics 2009, 9, 2149–2162. [Google Scholar] [CrossRef] [PubMed]
- Flores, C.A.; Bezerra, J.; Bustamante, S.A.; Goda, T.; MacDonald, M.P.; Kaplan, M.L.; Koldovský, O. Age-related changes in sucrase and lactase activity in the small intestine of 3- and 10-week-old obese mice (C57BL/6Jobob). J. Am. Coll. Nutr. 1990, 9, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Seeley, R.J.; Rushing, P.A.; D’Alessio, D.; Tso, P. A controlled high-fat diet induces an obese syndrome in rats. J. Nutr. 2003, 133, 1081–1087. [Google Scholar] [PubMed]
- Adachi, T.; Mori, C.; Sakurai, K.; Shihara, N.; Tsuda, K.; Yasuda, K. Morphological changes and increased sucrase and isomaltase activity in small intestines of insulin-deficient and type 2 diabetic rats. Endocr. J. 2003, 50, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Cho, S.H.; Kim, H.J.; Lee, H.J. Effects of soluble dietary fibers on lipid metabolism and activities of intestinal disaccharidases in rats. J. Nutr. Sci. Vitaminol. 1998, 44, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Doi, K. Effect of konjac fibre (glucomannan) on glucose and lipids. Eur. J. Clin. Nutr. 1995, 49 (Suppl. 3), S190–S197. [Google Scholar] [PubMed]
- Manoj, G.; Thampi, B.S.; Leelamma, S.; Menon, P.V. Effect of dietary fiber on the activity of intestinal and fecal β-glucuronidase activity during 1,2-dimethylhydrazine induced colon carcinogenesis. Plant Foods Hum. Nutr. 2001, 56, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.T.; Chen, H.L. Effects of konjac glucomannan on putative risk factors for colon carcinogenesis in rats fed a high-fat diet. J. Agric. Food Chem. 2011, 59, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.S.; Hoskins, L.C. Mucin degradation in human colon ecosystems. Fecal population densities of mucin-degrading bacteria estimated by a “most probable number” method. Gastroenterology 1981, 81, 759–765. [Google Scholar] [PubMed]
- Okazaki, Y.; Fukuda, E.; Chiji, H.; Kato, N. Edible lily bulb modulates colonic barrier functions, microflora and fermentation in rats fed a high-fat diet. J. Nutr. Health Food Sci. 2014, 2, 1–7. [Google Scholar] [CrossRef]
- Reddy, B.S.; Hanson, D.; Mangat, S.; Mathews, L.; Sbaschnig, M.; Sharma, C.; Simi, B. Effect of high-fat, high-beef diet and of mode of cooking of beef in the diet on fecal bacterial enzymes and fecal bile acids and neutral sterols. J. Nutr. 1980, 110, 1880–1887. [Google Scholar] [PubMed]


{kind=link}
| Ingredient | NC 1 | HF 2 | HF + LC 3 | HF + CO 4 |
|---|---|---|---|---|
| Lard | - | 10 | 10 | 10 |
| Cholesterol | - | 0.5 | 0.5 | 0.5 |
| Cholic acid | - | 0.1 | 0.1 | 0.1 |
| Chitosan 5 | - | - | 5 | - |
| Chitosan 6 oligosaccharide | - | - | - | 5 |
| Chow diet | 100 | 89.4 | 84.4 | 84.4 |
| Diet | NC | HF | HF + LC | HF + CO |
|---|---|---|---|---|
| Initial body weight (g) | 158.8 ± 18.4 | 164.3 ± 12.0 | 160.4 ± 11.8 | 162.1 ± 12.6 |
| Final body weight (g) | 476.8 ± 21.2 * | 550.5 ± 51.7 | 489.7 ± 49.5 * | 495.0 ± 51.9 |
| Body weight gain (g) | 318.0 ± 23.0 * | 386.2 ± 43.9 | 329.4 ± 47.7 * | 332.9 ± 56.0 |
| Liver weight (g) | 12.6 ± 0.6 * | 25.5 ± 3.8 | 19.2 ± 3.7 *,# | 23.8 ± 3.4 |
| Relative liver weight | - | - | - | - |
| (g/100 g BW) | 2.6 ± 0.1* | 4.6 ± 0.4 | 3.9 ± 0.4 *,# | 4.9 ± 0.9 |
| Perirenal fat (g) | 8.9 ± 2.5* | 14.0 ± 4.7 | 9.9 ± 2.7 * | 9.9 ± 1.9 * |
| Relative perirenal fat weight | - | - | - | - |
| (g/100 g BW) | 1.85 ± 0.5 * | 2.6 ± 0.6 | 1.9 ± 0.5 * | 2.0 ± 0.3 * |
| Food intake (g/day) | 30.2 ± 3.1 | 28.6 ± 3.4 | 27.5 ± 4.7 | 28.3 ± 3.4 |
| Feed efficiency (%) | 7.3 ± 0.8 * | 9.1 ± 1.2 | 7.8 ± 1.0 * | 8.2 ± 1.5 |
| Diet | NC | HF | HF + LC | HF + CO |
|---|---|---|---|---|
| Total cholesterol (mg/dL) | 52.5 ± 8.3 * | 116.9 ± 50.5 | 73.6 ± 14.9 * | 77.2 ± 24.8 |
| HDL-C (mg/dL) | 30.6 ± 1.8 * | 21.5 ± 11.7 | 18.2 ± 8.8 | 13.3 ± 3.6 |
| LDL-C (mg/dL) | 16.3 ± 6.4 * | 36.6 ± 13.6 | 28.0 ± 8.5 | 32.1 ± 17.2 |
| VLDL-C (mg/dL) | 5.6 ± 2.8 * | 58.8 ± 43.8 | 27.4 ± 12.2 * | 31.8 ± 9.9 |
| LDL-C + VLDL-C | 21.9 ± 7.5 * | 95.4 ± 49.0 | 55.4 ± 17.4 * | 63.9 ± 26.5 |
| TC/HDL-C ratio | 1.7 ± 0.2 * | 6.7 ± 5.5 | 5.1 ± 3.2 | 6.3 ± 3.1 |
| HDL-C/(LDL-C + VLDL-C) ratio | 1.6 ± 0.7 * | 0.27 ± 0.2 | 0.38 ± 0.3 | 0.25 ± 0.1 |
| Triglyceride (mg/dL) | 46.2 ± 19.1 | 36.3 ± 7.9 | 29.4 ± 7.7 | 25.2 ± 6.6 * |
| TNF-α (pg/mL) | 18.5 ± 1.5 * | 24.7 ± 1.8 | 11.4 ± 4.9 *,# | 25.8 ± 17.0 |
| Diet | NC | HF | HF + LC | HF + CO |
|---|---|---|---|---|
| Total cholesterol | - | - | - | - |
| (mg/g liver) | 1.5 ± 0.41 * | 63.1 ± 10.8 | 34.7 ± 15.5 *,# | 67.7 ± 19.4 |
| (g/liver) | 0.02 ± 0.01 * | 1.60 ± 0.37 | 0.66 ± 0.30 *,# | 1.60 ± 0.57 |
| Triglyceride | - | - | - | - |
| (mg/g liver) | 9.8 ± 2.2 * | 47.2 ± 15.9 | 38.8 ± 11.1 | 38.6 ± 15.9 |
| (g/liver) | 0.12 ± 0.03 * | 1.20 ± 0.46 | 0.76 ± 0.31 * | 0.93 ± 0.48 |
| Fatty acid synthetase | 2.5 ± 2.0 | 3.3 ± 1.1 | 2.1 ± 0.9 * | 2.2 ± 1.1 * |
| (nmole NADPH/min/mg protein) |
| Diet | NC | HF | HF + LC | HF + CO |
|---|---|---|---|---|
| Feces wet weight (g/day) | 9.7 ± 1.7 | 8.5 ± 2.3 | 10.7 ± 1.8 * | 9.9 ± 1.7 |
| Feces dry weight (g/day) | 6.2 ± 0.7 | 5.7 ± 1.1 | 6.2 ± 0.6 | 5.9 ± 0.8 |
| Total cholesterol | - | - | - | - |
| (mg/g feces) | 6.6 ± 1.5 * | 12.8 ± 3.1 | 15.5 ± 1.7 *,# | 12.9 ± 2.5 |
| (mg/day) | 40.6 ± 11.3 * | 71.0 ± 23.4 | 96.5 ± 12.6 * | 77.3 ± 22.0 |
| Triglyceride | - | - | - | - |
| (mg/g feces) | 12.2 ± 1.2 | 12.8 ± 0.9 | 13.1 ± 0.9 | 13.6 ± 1.2 |
| (mg/day) | 75.6 ± 13.3 | 69.6 ± 11.9 | 81.8 ± 10.9 * | 80.8 ± 14.9 |
| Diet | NC | HF | HF + LC | HF + CO |
|---|---|---|---|---|
| Maltase (mg glucose/min/mg protein) | 774.7 ± 124.4 * | 986.8 ± 73.2 | 837.6 ± 128.6 *,# | 990.2 ± 30.9 |
| Lactase (mg glucose/min/mg protein) | 152.2 ± 50.7 * | 205.1 ± 31.6 | 163.5 ± 41.9 * | 184.7 ± 18.4 |
| Sucrase (mg glucose/min/mg protein) | 217.6 ± 66.4 * | 329.3 ± 62.2 | 228.2 ± 103.6 * | 266.2 ± 59.4 |
| Mucinase (nmole reducing sugar/min/mg protein) | 3.6 ± 0.9 * | 4.9 ± 1.0 | 4.0 ± 1.1 | 3.1 ± 0.6 * |
| β-glucuronidase (nmole phenolphthalein/min/mg protein) | 2.4 ± 0.4 * | 3.1 ± 0.5 | 2.2 ± 0.7 * | 2.5 ± 0.2 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-Y.; Feng, S.-A.; Liu, S.-H.; Chiang, M.-T. Functional Comparison for Lipid Metabolism and Intestinal and Fecal Microflora Enzyme Activities between Low Molecular Weight Chitosan and Chitosan Oligosaccharide in High-Fat-Diet-Fed Rats. Mar. Drugs 2017, 15, 234. https://doi.org/10.3390/md15070234
Chiu C-Y, Feng S-A, Liu S-H, Chiang M-T. Functional Comparison for Lipid Metabolism and Intestinal and Fecal Microflora Enzyme Activities between Low Molecular Weight Chitosan and Chitosan Oligosaccharide in High-Fat-Diet-Fed Rats. Marine Drugs. 2017; 15(7):234. https://doi.org/10.3390/md15070234
Chicago/Turabian StyleChiu, Chen-Yuan, Shih-An Feng, Shing-Hwa Liu, and Meng-Tsan Chiang. 2017. "Functional Comparison for Lipid Metabolism and Intestinal and Fecal Microflora Enzyme Activities between Low Molecular Weight Chitosan and Chitosan Oligosaccharide in High-Fat-Diet-Fed Rats" Marine Drugs 15, no. 7: 234. https://doi.org/10.3390/md15070234
APA StyleChiu, C.-Y., Feng, S.-A., Liu, S.-H., & Chiang, M.-T. (2017). Functional Comparison for Lipid Metabolism and Intestinal and Fecal Microflora Enzyme Activities between Low Molecular Weight Chitosan and Chitosan Oligosaccharide in High-Fat-Diet-Fed Rats. Marine Drugs, 15(7), 234. https://doi.org/10.3390/md15070234