Isolation and Tissue Distribution of an Insulin-Like Androgenic Gland Hormone (IAG) of the Male Red Deep-Sea Crab, Chaceon quinquedens

Abstract
:1. Introduction
2. Results
2.1. cDNA Sequence Analysis
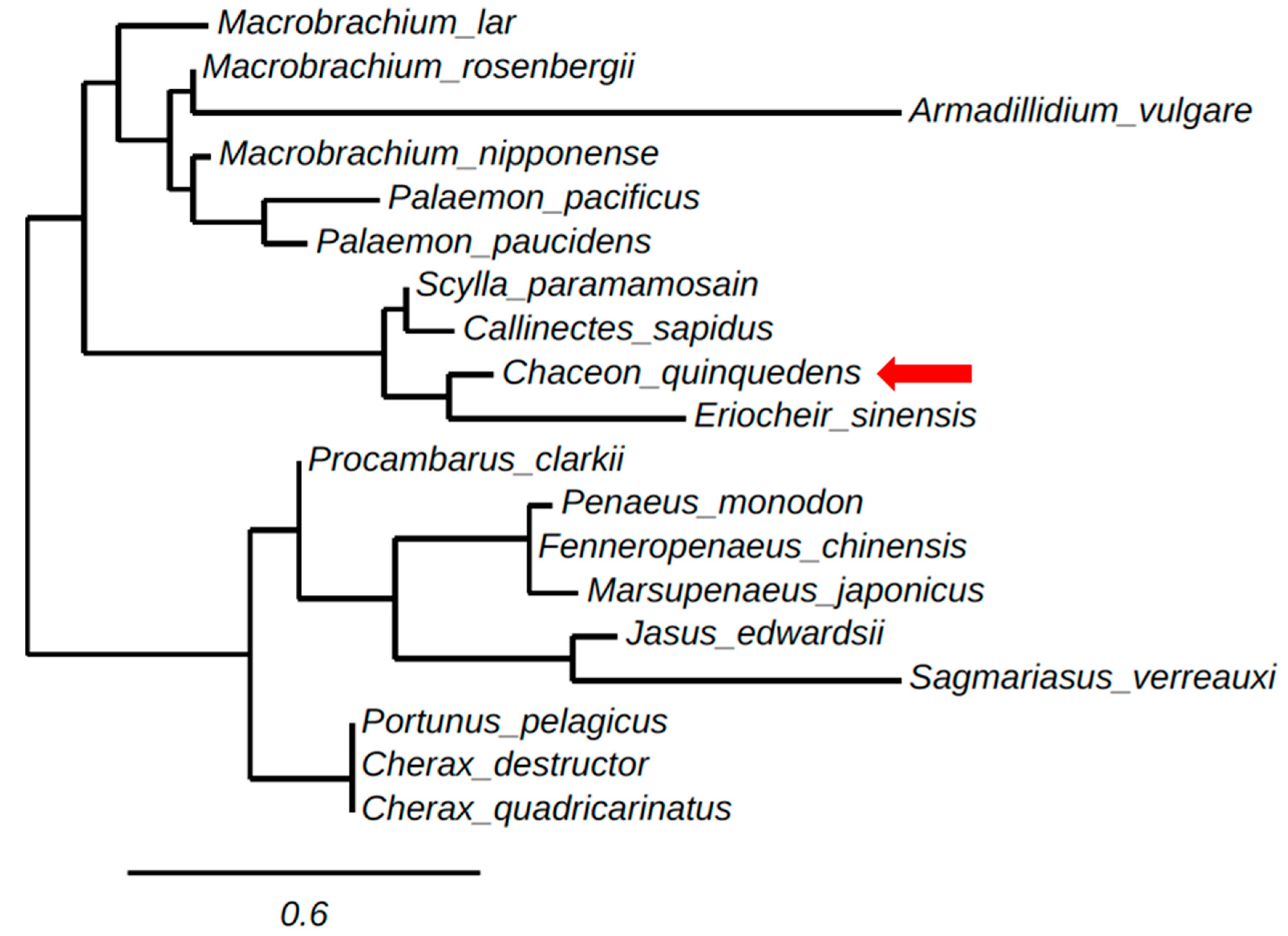
2.2. Phylogenetic Tree Analysis
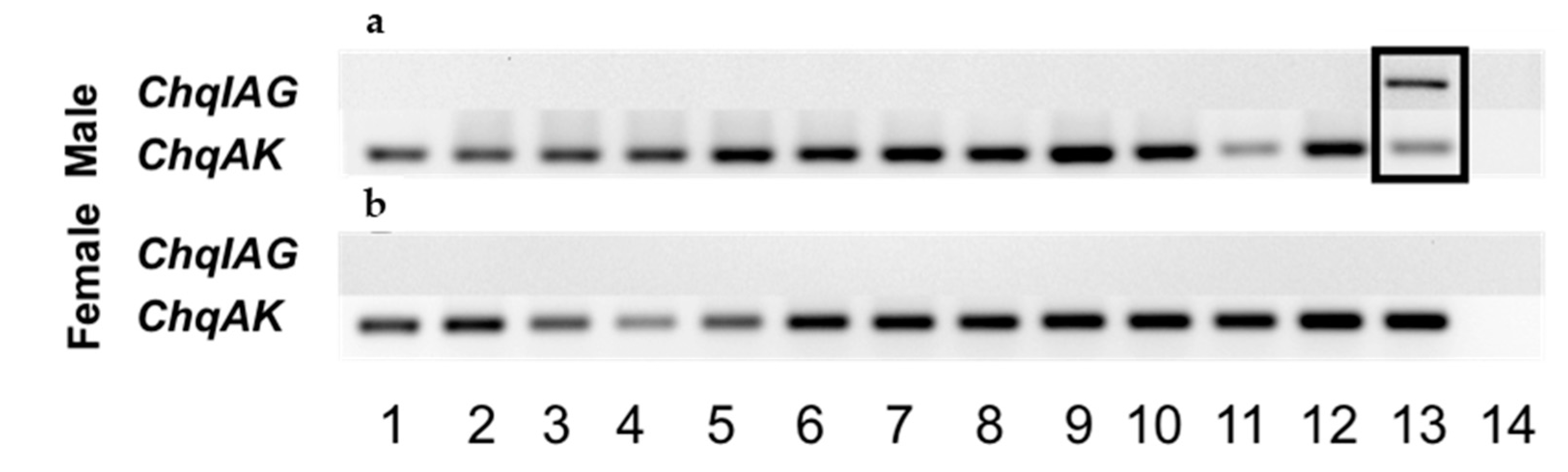
2.3. Tissue Distribution of ChqIAG
2.4. Translational Regulatory Sites in ChqIAG cDNA Sequence
3. Discussion
4. Materials and Methods
4.1. Animal Collection and Maintenance
4.2. Isolation of the Full-Length ChqIAG cDNA Using PCR with Degenerate Primers
4.3. 5′ and 3′ RACE of ChqIAG
4.4. Sequence Analyses
4.5. Tissue Distribution of ChqIAG Expression
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cronin, L.E. Anatomy and histology of the male reproductive system of Callinectes sapidus Rathbun. J. Morphol. 1947, 81, 209–239. [Google Scholar] [CrossRef] [PubMed]
- Charniaux-Cotton, H. Decouverte chez un crustace amphipode (Orchestia gammarella) d’une glande endocrine responsable de la differentiation de caracteres sexuels primaires et secondaires males. C. R. Acad. Sci. Paris 1954, 239, 780–782. [Google Scholar] [PubMed]
- Charniaux-Cotton, H. Androgenic gland of crustaceans. Gen. Comp. Endocrinol. 1962, 1, 241–247. [Google Scholar] [CrossRef]
- Suzuki, S.; Yamasaki, K. Sex reversal by implantations of ethanol-treated androgenic glands of female isopods, Armadillidium vulgare (Malacostraca, crustacea). Gen. Comp. Endocrinol. 1998, 111, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Manor, R.; Aflalo, E.D.; Segall, C.; Weil, S.; Azulay, D.; Ventura, T.; Sagi, A. Androgenic gland implantation promotes growth and inhibits vitellogenesis in Cherax quadricarinatus females held in individual compartments. Invertebr. Reprod. Dev. 2004, 45, 151–159. [Google Scholar] [CrossRef]
- Martin, G.; Sorokine, O.; Moniatte, M.; Bulet, P.; Hetru, C.; Van Dorsselaer, A. The structure of a glycosylated protein hormone responsible for sex determination in the isopod, Armadillidium vulgare. Eur. J. Biochem. 1999, 262, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Okuno, A.; Hasegawa, Y.; Ohira, T.; Katakura, Y.; Nagasawa, H. Characterization and cDNA cloning of androgenic gland hormone of the terrestrial isopod Armadillidium vulgare. Biochem. Biophys. Res. Commun. 1999, 264, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Khalaila, I.; Rosen, O.; Sagi, A. Expression of an androgenic gland-specific insulin-like peptide during the course of prawn sexual and morphotypic differentiation. ISRN Endocrinol. 2011, 2011, 476283. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ye, H.; Huang, H.; Yang, Y.; Gong, J. An insulin-like androgenic gland hormone gene in the mud crab, Scylla paramamosain, extensively expressed and involved in the processes of growth and female reproduction. Gen. Comp. Endocrinol. 2014, 204, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.S.; Manor, R.; Sagi, A. Cloning of an insulin-like androgenic gland factor (IAG) from the blue crab, Callinectes sapidus: Implications for eyestalk regulation of IAG expression. Gen. Comp. Endocrinol. 2011, 173, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Fitzgibbon, Q.; Battaglene, S.; Sagi, A.; Elizur, A. Identification and characterization of androgenic gland specific insulin-like peptide-encoding transcripts in two spiny lobster species: Sagmariasus verreauxi and Jasus edwardsii. Gen. Comp. Endocrinol. 2015, 214, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, F.; Sun, Z.; Xiang, J. Two spliced variants of insulin-like androgenic gland hormone gene in the Chinese shrimp, Fenneropenaeus chinensis. Gen. Comp. Endocrinol. 2012, 177, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Bai, H.; Xiong, Y.; Fu, H.; Jiang, S.; Jiang, F.; Jin, S.; Sun, S.; Qiao, H.; Zhang, W. Molecular characterization of insulin-like androgenic gland hormone-binding protein gene from the oriental river prawn Macrobrachium nipponense and investigation of its transcriptional relationship with the insulin-like androgenic gland hormone gene. Gen. Comp. Endocrinol. 2015, 216, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.S. An insulin-like growth factor found in hepatopancreas implicates carbohydrate metabolism of the blue crab Callinectes sapidus. Gen. Comp. Endocrinol. 2014, 199, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.S.; Zmora, N. Functional studies of crustacean hyperglycemic hormones (CHHs) of the blue crab, Callinectes sapidus—The expression and release of CHH in eyestalk and pericardial organ in response to envir. FEBS J. 2008, 275, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Zmora, N.; Chung, J.S. A novel hormone is required for the development of reproductive phenotypes in adult female crabs. Endocrinology 2014, 155, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Stevens, B.G.; Guida, V. Depth and temperature distribution, morphometrics, and sex ratios of red deepsea crab (Chaceon quinquedens) at 4 sampling sites in the mid-atlantic bight. Fish. Bull. 2016, 114, 343–359. [Google Scholar] [CrossRef]
- Zhang, Y.; Qiao, K.; Wang, S.; Peng, H.; Shan, Z.; Wang, K. Molecular identification of a new androgenic gland-specific insulin-like gene from the mud crab, Scylla paramamosain. Aquaculture 2014, 433, 325–334. [Google Scholar] [CrossRef]
- Tanguay, R.L.; Gallie, D.R. Translational efficiency is regulated by the length of the 3′ untranslated region. Mol. Cell. Biol. 1996, 16, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C.; Posakony, J.W. The Bearded box, a novel 3′ UTR sequence motif, mediates negative post-transcriptional regulation of Bearded and Enhancer of split Complex gene expression. Development 1997, 124, 4847–4856. [Google Scholar] [PubMed]
- Hellen, T.U.C.; Sarrow, P. Internal ribosome entry sites in eukaryotic mRNA molecules. J. Virol. 2001, 85, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.S.; Ahn, I.S.; Yu, O.H.; Kim, D.S. Crustacean hyperglycemic hormones of two cold water crab species, Chionoecetes opilio and C. japonicus: Isolation of cDNA sequences and localization of CHH neuropeptide in eyestalk ganglia. Gen. Comp. Endocrinol. 2015, 214, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.B.; Powell, G.C. Size at maturity in the male alaskan tanner crab, Chionoecetes bairdi, as determined by chela allometry, reproductive tract weights, and size of precopulatory males. J. Fish. Res. Board Can. 1972, 29, 423–427. [Google Scholar] [CrossRef]
- Rosen, O.; Weil, S.; Manor, R.; Roth, Z.; Khalaila, I.; Sagi, A. A crayfish insulin-like-binding protein: Another piece in the androgenic gland insulin-like hormone puzzle is revealed. J. Biol. Chem. 2013, 288, 22289–22298. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Chien, C.H.; Jen, K.H.; Huang, H.D. RegRNA: A regulatory RNA motifs and elements finder. Nucleic Acids Res. 2006, 34, W429–W434. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′ → 3′) |
|---|---|
| dF1 | GAYTTYGAYTGYGGNSAYYT |
| dR2 | ACTGCGGCSACMTSGSCGACA |
| ChqIAG-3F1 | ACCAAGAACCTTATCCTCTGTATGCCTGACC |
| ChqIAG-3F2 | GCCCCCGCCCACCTCACGCAGTCCCCATGA |
| ChqIAG-3F3 | AGCGGTCTGTCAACGTGCACGACGAGTGCTGC |
| ChqIAG-5R1 | GCAGCACTCGTCGTGCACGTTGACAGACCGC |
| ChqIAG-5R2 | TGTCCATTGTGTCTTGACAAGCTGGAAGGCGA |
| ChqIAG-5R3 | ATCATCAGGGGTGTCAGCGCCCTGTGACTCGG |
| ChqIAG-st | ATGTTCCTGCCCGTGATTATACTACTGAT |
| ChqIAG-end | TACTGATTCTGAGCTGCTATAATTTCCCT |
| ChqAK-QF | CTGGGCCAGGTATACCGCCGCCTTGTCAGC |
| ChqAK-QR | GGGGAGCTTGATGTGGACGGAGGCACGCAC |
| TOP | IRES | uORF | GAIT Element | SECIS | ADH_DRE | KB | CPE | GY | Brd | BRE | |
| Species: | |||||||||||
| C. quinquedens | 5 | 1 | 1 | 1 | 1 | ||||||
| M. nipponense | 6 | ||||||||||
| P. pelagicus | |||||||||||
| M. rosenbergii | 9 | 1 | |||||||||
| E. sinensis | 7 | 1 | |||||||||
| P. pacificus | 3 | 3 | |||||||||
| C. destructor | |||||||||||
| S. verreauxi | 9 | 1 | |||||||||
| S. paramamosain | 6 | 1 | |||||||||
| M. japonicus | 4 | ||||||||||
| M. lar | 6 | ||||||||||
| C. quadricarinatus | 4 | 2 | |||||||||
| C. sapidus | 3 | ||||||||||
| F. chinensis | 5 | ||||||||||
| P. paucidens | 2 | 1 | |||||||||
| P. clarkii | 3 | ||||||||||
| TOP | IRES | uORF | GAIT element | SECIS | ADH_DRE | KB | CPE | GY | Brd | BRE | |
| C. quinquedens | 5 | 4 | 3 | 1 | |||||||
| M. nipponense | 24 | 2 | 4 | 1 | 1 | 3 | |||||
| P. pelagicus | 5 | 1 | 2 | 2 | 1 | ||||||
| M. rosenbergii | 31 | 3 | 1 | 1 | 1 | 3 | |||||
| E. sinensis | 7 | 3 | 2 | 2 | 1 | 1 | |||||
| P. pacificus | 23 | 3 | 1 | 1 | 8 | ||||||
| C. destructor | 3 | 1 | 1 | 2 | 1 | ||||||
| S. verreauxi | 9 | 2 | 4 | 6 | 2 | ||||||
| S. paramamosain | 2 | 3 | 2 | 6 | 1 | 1 | |||||
| M. japonicus | 2 | 1 | |||||||||
| M. lar | 24 | 2 | 3 | 4 | 1 | 1 | 1 | 1 | 1 | ||
| C. quadricarinatus | 2 | 1 | 2 | 1 | 1 | ||||||
| C. sapidus | 3 | 2 | 2 | 1 | 1 | 1 | |||||
| F. chinensis | 12 | 4 | 2 | 1 | |||||||
| P. paucidens | 8 | 2 | 1 | ||||||||
| P. clarkii | 1 | ||||||||||
| motif distribution | 233 | 27 | 36 | 32 | 10 | 1 | 1 | 4 | 8 | 19 | 1 |
| motif frequency | 0.63 | 0.07 | 0.1 | 0.09 | 0.03 | 0.003 | 0.003 | 0.01 | 0.02 | 0.05 | 0.003 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, A.; Green, S.; Chung, J.S. Isolation and Tissue Distribution of an Insulin-Like Androgenic Gland Hormone (IAG) of the Male Red Deep-Sea Crab, Chaceon quinquedens. Mar. Drugs 2017, 15, 241. https://doi.org/10.3390/md15080241
Lawrence A, Green S, Chung JS. Isolation and Tissue Distribution of an Insulin-Like Androgenic Gland Hormone (IAG) of the Male Red Deep-Sea Crab, Chaceon quinquedens. Marine Drugs. 2017; 15(8):241. https://doi.org/10.3390/md15080241
Chicago/Turabian StyleLawrence, Amanda, Shadaesha Green, and Jum Sook Chung. 2017. "Isolation and Tissue Distribution of an Insulin-Like Androgenic Gland Hormone (IAG) of the Male Red Deep-Sea Crab, Chaceon quinquedens" Marine Drugs 15, no. 8: 241. https://doi.org/10.3390/md15080241
APA StyleLawrence, A., Green, S., & Chung, J. S. (2017). Isolation and Tissue Distribution of an Insulin-Like Androgenic Gland Hormone (IAG) of the Male Red Deep-Sea Crab, Chaceon quinquedens. Marine Drugs, 15(8), 241. https://doi.org/10.3390/md15080241