Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives



Abstract
:1. Introduction
2. Antimicrobial Compounds from Marine Microorganisms
2.1. Antimicrobial Compounds from Bacteria
2.2. Antimicrobial Compounds from Fungi
3. Improving the Biodiscovery Pipeline for Deep-Sea Antibiotics
3.1. Sampling Techniques
3.2. Isolation and Cultivation Techniques
3.3. Metagenomics
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Corinaldesi, C. New perspectives in benthic deep-sea microbial ecology. Front. Mar. Sci. 2015, 2, 1–12. [Google Scholar] [CrossRef]
- Thistle, D. The deep-sea floor: An overview. In Ecosystems of the Deep Oceans; Tyler, A., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2003; Volume 28, pp. 6–8. [Google Scholar]
- Danovaro, R.; Carugati, L.; Berzano, M.; Cahill, A.E.; Carvalho, S.; Chenuil, A. Implementing and Innovating Marine Monitoring Approaches for Assessing Marine Environmental Status. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Schrenk, M.O.; Kelley, D.S.; Delaney, J.R.; Baross, J.A. Incidence and diversity of microorganisms within the walls of an active deep-sea sulfide chimney. Appl. Environ. Microbiol. 2003, 69, 3580–3592. [Google Scholar] [CrossRef] [PubMed]
- Skropeta, D. Deep-sea natural products. Nat. Prod. Rep. 2008, 25, 1131. [Google Scholar] [CrossRef] [PubMed]
- Thaler, A.D.; Van Dover, C.L.; Vilgalys, R. Ascomycete phylotypes recovered from a Gulf of Mexico methane seep are identical to an uncultured deep-sea fungal clade from the Pacific. Fungal Ecol. 2012, 5, 270–273. [Google Scholar] [CrossRef]
- Jebbar, M.; Franzetti, B.; Girard, E.; Oger, P. Microbial diversity and adaptation to high hydrostatic pressure in deep-sea hydrothermal vents prokaryotes. Extremophiles 2015, 19, 721–740. [Google Scholar] [CrossRef] [PubMed]
- Skropeta, D.; Wei, L. Recent advances in deep-sea natural products. Nat. Prod. Rep. 2014, 31, 999–1025. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The Antibiotic Resistance Crisis. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Goodfellow, M.; Fiedler, H.P. A guide to successful bioprospecting: Informed by actinobacterial systematics. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2010, 98, 119–142. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mageed, W.M.; Milne, B.F.; Wagner, M.; Schumacher, M.; Sandor, P.; Pathom-aree, W.; Goodfellow, M.; Bull, A.T.; Horikoshi, K.; Ebel, R.; et al. Dermacozines, a new phenazine family from deep-sea dermacocci isolated from a Mariana Trench sediment. Org. Biomol. Chem. 2010, 8, 2352–2362. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Yang, T.; Ren, X.; Liu, J.; Song, Y.; Sun, A.; Ma, J.; Wang, B.; Zhang, Y.; Huang, C.; et al. Cytotoxic angucycline class glycosides from the deep sea actinomycete Streptomyces lusitanus SCSIO LR32. J. Nat. Prod. 2012, 75, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tian, X.; Niu, S.; Zhang, W.; Chen, Y.; Zhang, H.; Yang, X.; Zhang, W.; Li, W.; Ju, J.; et al. Pseudonocardians A-C, new diazaanthraquinone derivatives from a deap-sea actinomycete Pseudonocardia sp. SCSIO 01299. Mar. Drugs. 2011, 9, 1428–1439. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Iwata, F.; Yamada, S.; Kawahara, H.; Katayama, M. Usabamycins A-C: New anthramycin-type analogues from a marine-derived actinomycete. Bioorganic Med. Chem. Lett. 2011, 21, 7099–7101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, Z.; Li, S.; Yang, T.; Zhang, Q.; Ma, L.; Tian, X.; Zhang, H.; Huang, C.; Zhang, S.; et al. Spiroindimicins A-D: New bisindole alkaloids from a deep-sea-derived actinomycete. Org. Lett. 2012, 14, 3364–3367. [Google Scholar] [CrossRef] [PubMed]
- Bérdy, J. Bioactive Microbial Metabolites. J. Antibiot. (Tokyo) 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.P.; Tang, S.K.; Dong, J.D.; Zhang, Y.Q.; Xu, L.H.; Zhang, S.; Li, W.J. Marinactinospora thermotolerans gen. nov., sp. nov., a marine actinomycete isolated from a sediment in the northern South China Sea. Int. J. Syst. Evol. Microbiol. 2009, 59, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Huang, H.; Chen, Y.; Tan, J.; Song, Y.; Zou, J.; Tian, X.; Hua, Y.; Ju, J. Marthiapeptide A, an anti-infective and cytotoxic polythiazole cyclopeptide from a 60 L scale fermentation of the deep sea-derived Marinactinospora thermotolerans SCSIO 00652. J. Nat. Prod. 2012, 75, 2251–2255. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, Q.; Liu, X.; Chen, Y.; Zhang, Y.; Sun, A.; Zhang, W.; Zhang, J.; Ju, J. Cyclic hexapeptides from the deep South China sea-derived Streptomyces scopuliridis SCSIO ZJ46 active against pathogenic gram-positive bacteria. J. Nat. Prod. 2014, 77, 1937–1941. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Huang, H.; Li, J.; Song, Y.; Jiang, R.; Liu, J.; Zhang, S.; Hua, Y.; Ju, J. New anti-infective cycloheptadepsipeptide congeners and absolute stereochemistry from the deep sea-derived Streptomyces drozdowiczii SCSIO 10141. Tetrahedron 2014, 70. [Google Scholar] [CrossRef]
- Ballard, C.E.; Yu, H.; Wang, B. Recent Developments in Depsipeptide Research. Curr. Med. Chem. 2002, 9, 471–498. [Google Scholar] [CrossRef] [PubMed]
- Jimeno, J.; Faircloth, G.; Sousa-Faro, J.F.; Scheuer, P.; Rinehart, K. New Marine Derived Anticancer Therapeutics—A Journey from the Sea to Clinical Trials. Mar. Drugs 2004, 2, 14–29. [Google Scholar] [CrossRef]
- Lee, Y.; Phat, C.; Hong, S.C. Structural diversity of marine cyclic peptides and their molecular mechanisms for anticancer, antibacterial, antifungal, and other clinical applications. Peptides 2017, 95, 94–105. [Google Scholar] [CrossRef] [PubMed]
- March, D.R.; Abbenante, G.; Bergman, D.A.; Brinkworth, R.I.; Wickramasinghe, W.; Begun, J.; Martin, J.L.; Fairlie, D.P. Substrate-based cyclic peptidomimetics of Phe-Ile-Val that inhibit HIV-1 protease using a novel enzyme-binding mode. J. Am. Chem. Soc. 1996, 118, 3375–3379. [Google Scholar] [CrossRef]
- Ariawan, A.D.; Webb, J.E.; Howe, E.N.; Gale, P.A.; Thordarson, P.; Hunter, L. Cyclic peptide unguisin A is an anion receptor with high affinity for phosphate and pyrophosphate. Org. Biomol. Chem. 2017, 15, 2962–2967. [Google Scholar] [CrossRef] [PubMed]
- Rezai, T.; Yu, B.; Millhauser, G.L.; Jacobson, M.P.; Lokey, R.S. Testing the conformational hypothesis of passive membrane permeability using synthetic cyclic peptide diastereomers. J. Am. Chem. Soc. 2006, 128, 2510–2511. [Google Scholar] [CrossRef] [PubMed]
- Batiste, S.M.; Johnston, J.N. Evidence for Ion-Templation During Macrocyclooligomerization of Depsipeptides. J. Am. Chem. Soc. 2018, 140, 4560–4568. [Google Scholar] [CrossRef] [PubMed]
- Ahlbach, C.L.; Lexa, K.W.; Bockus, A.T.; Chen, V.; Crews, P.; Jacobson, M.P.; Lokey, R.S. Beyond cyclosporine A: Conformation-dependent passive membrane permeabilities of cyclic peptide natural products. Future Med. Chem. 2015, 7, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Zorzi, A.; Deyle, K.; Heinis, C. Cyclic peptide therapeutics: Past, present and future. Curr. Opin. Chem. Biol. 2017, 38, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Jelokhani-Niaraki, M.; Hodges, R.S.; Meissner, J.E.; Hassenstein, U.E.; Wheaton, L. Interaction of gramicidin S and its aromatic amino-acid analog with phospholipid membranes. Biophys. J. 2008, 95, 3306–3321. [Google Scholar] [CrossRef] [PubMed]
- Jelokhani-Niaraki, M.; Prenner, E.J.; Kay, C.M.; McElhaney, R.N.; Hodges, R.S. Conformation and interaction of the cyclic cationic antimicrobial peptides in lipid bilayers. J. Pept. Res. 2002, 60, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.D.; Jensen, P.R.; Fenical, W. Lobophorins A and B, new antiinflammatory macrolides produced by a tropical marine bacterium. Bioorg. Med. Chem. Lett. 1999, 9, 2003–2006. [Google Scholar] [CrossRef]
- Niu, S.; Li, S.; Chen, Y.; Tian, X.; Zhang, H.; Zhang, G.; Zhang, W.; Yang, X.; Zhang, S.; Ju, J.; et al. Lobophorins e and F, new spirotetronate antibiotics from a South China Sea-derived Streptomyces sp. SCSIO 01127. J. Antibiot. (Tokyo) 2011, 64, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Q.; Zhang, S.Y.; Wang, N.; Li, Z.L.; Hua, H.M.; Hu, J.C.; Wang, S.J. New spirotetronate antibiotics, lobophorins H and I, from a South China Sea-derived streptomyces sp. 12A35. Mar. Drugs 2013, 11, 3891–3901. [Google Scholar] [CrossRef] [PubMed]
- Lacoske, M.H.; Theodorakis, E.A. Spirotetronate polyketides as leads in drug discovery. J. Nat. Prod. 2015, 78, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Martinez-Bueno, M.; Molina-Henares, A.J.; Teran, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR Family of Transcriptional Repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bister, B.; Bischoff, D.; Ströbele, M.; Riedlinger, J.; Reicke, A.; Wolter, F.; Bull, A.T.; Zähner, H.; Fiedler, H.P.; Süssmuth, R.D. Abyssomicin C—A polycyclic antibiotic from a marine Verrucosispora strain as an inhibitor of the p-aminobenzoic acid/tetrahydrofolate biosynthesis pathway. Angew. Chem. Int. Ed. 2004, 43, 2574–2576. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Huang, H.; Chen, Y.; Ding, J.; Zhang, Y.; Sun, A.; Zhang, W.; Ju, J. Cytotoxic and antibacterial marfuraquinocins from the deep south china sea-derived streptomyces niveus scsio 3406. J. Nat. Prod. 2013, 76, 2263–2268. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, C.; Schneider, K.; Bruntner, C.; Irran, E.; Nicholson, G.; Bull, A.T.; Jones, A.L.; Brown, R.; Stach, J.E.; Goodfellow, M.; et al. Caboxamycin, a new antibiotic of the benzoxazole family produced by the deep-sea strain Streptomyces sp. NTK 937. J. Antibiot. (Tokyo) 2009, 62, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahé, S.; Rédou, V.; Calvez, T.L.; Vandenkoornhuyse, P.; Burgaud, G. Fungi in Deep-Sea Environments and Metagenomics. In The Ecological Genomics of Fungi; Martin, F., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 325–354. [Google Scholar]
- Bhatnagar, I.; Kim, S.K. Immense Essence of Excellence: Marine Microbial Bioactive Compounds. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs 2010, 8, 2673–2701. [Google Scholar] [CrossRef] [PubMed]
- Okutani, K. Gliotoxin produced by a strain of Aspergillus isolated from marine mud. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 995–1000. [Google Scholar] [CrossRef]
- Blunt, J.; Copp, B.; Keyzers, R.; Munro, M.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Zhang, Y.; Xu, X.Y.; Qi, S.H. Diverse Deep-Sea Fungi from the South China Sea and Their Antimicrobial Activity. Curr. Microbiol. 2013, 67, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.M.; Sousa, M.E.; Nascimento, M.S.J. Xanthone derivatives: New insights in biological activities. Curr. Med. Chem. 2005, 12, 2517–2538. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.M.M.; Castanheiro, R.A.P. Synthesis of Prenylated Xanthones: An Overview. Curr. Org. Chem. 2009, 13, 1215–1240. [Google Scholar] [CrossRef]
- Fredimoses, M.; Zhou, X.; Lin, X.; Tian, X.; Ai, W.; Wang, J.; Liao, S.; Liu, J.; Yang, B.; Yang, X.; et al. New prenylxanthones from the deep-sea derived fungus Emericella sp. SCSIO 05240. Mar. Drugs 2014, 12, 3190–3202. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Wang, J.; Zhang, X.; Nong, X.; Xu, X.; Qi, S. Cytotoxic polyketides from the deep-sea-derived fungus Engyodontium album DFFSCS021. Mar. Drugs 2014, 12, 5902–5915. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.J.; Lin, S.H.; Lin, M.R.; Ku, H.; Szkaradek, N.; Marona, H.; Hsu, A.; Shiuan, D. Xanthone derivatives could be potential antibiotics: Virtual screening for the inhibitors of enzyme i of bacterial phosphoenolpyruvate-dependent phosphotransferase system. J. Antibiot. (Tokyo) 2013, 66, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Liu, D.; Hu, X.; Proksch, P.; Shao, Z.; Lin, W. Spiromastixones A-O, antibacterial chlorodepsidones from a deep-sea-derived spiromastix sp. fungus. J. Nat. Prod. 2014, 77, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liao, Y.; Chen, R.; Hou, Y.; Ke, W.; Zhang, B.; Gao, M.; Shao, Z.; Chen, J.; Li, F. Chlorinated Azaphilone Pigments with Antimicrobial and Cytotoxic Activities Isolated from the Deep Sea Derived Fungus Chaetomium sp. NA-S01-R1. Mar. Drugs 2018, 16, 61. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat Prod Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Zhang, Z.; Zhu, T.; Gu, Q.; Li, D. Penicyclones A-E, Antibacterial Polyketides from the Deep-Sea-Derived Fungus Penicillium sp. F23-2. J. Nat. Prod. 2015, 78, 2699–2703. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, D.; Zhu, T.; Cai, S.; Wang, F.; Xiao, X.; Gu, G. New alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]
- Egan, S.T.; McCarthy, D.M.; Patching, J.W.; Fleming, G.T.A. An investigation of the physiology and potential role of components of the deep ocean bacterial community (of the NE Atlantic) by enrichments carried out under minimal environmental change. Deep. Res. Part I Oceanogr. Res. Pap. 2012, 61, 11–20. [Google Scholar] [CrossRef]
- McCarthy, D.M.; Pearce, D.; Patching, J.W.; Fleming, G.T. Contrasting responses to nutrient enrichment of prokaryotic communities collected from deep sea sites in the southern ocean. Biology 2013, 2, 1165–1188. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Kato, C. Taxonomic studies of extremely barophilic bacteria isolated from the Mariana Trench and description of Moritella yayanosii sp. nov., a new barophilic bacterial isolate. Extremophiles 1999, 3, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Yanagibayashi, M.; Nogi, Y.; Li, L.; Kato, C. Changes in the microbial community in Japan Trench sediment from a depth of 6292 m during cultivation without decompression. FEMS Microbiol. Lett. 1999, 170, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohnber, H.-J.; Amann, H.; Abegg, F.; Mudrack, H.; Bohrmann, G.; Maggiulli, M. Pressurized Coring of Near-Surface Gas-Hydrate Sediments on Hydrate Ridge: The Multiple Autoclave Corer, and First Results from Pressure-Core X-ray CT Scans. In Proceedings of the EGS—AGU—EUG Joint Assembly, Nice, France, 6–11 April 2003. [Google Scholar]
- Parkes, R.J.; Sellek, G.; Webster, G.; Martin, D.; Anders, E.; Weightman, A.J.; Sass, H. Culturable prokaryotic diversity of deep, gas hydrate sediments: First use of a continuous high-pressure, anaerobic, enrichment and isolation system for subseafloor sediments (DeepIsoBUG). Environ. Microbiol. 2009, 11, 3140–3153. [Google Scholar] [CrossRef] [PubMed]
- Galloway, K.C.; Becker, K.P.; Phillips, B.; Kirby, J.; Licht, S.; Tchernov, D.; Wood, R.J.; Gruber, D.F. Soft Robotic Grippers for Biological Sampling on Deep Reefs. Soft Robot. 2016, 3, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zobell, C.E.; Morita, R.Y. Barophilic bacteria in some deep sea sediments. J. Bacteriol. 1957, 73, 563–568. [Google Scholar] [PubMed]
- Yayanos, A.A.; Dietz, A.S.; Van Boxtel, R. Isolation of a deep-sea barophilic bacterium and some of its growth characteristics. Science 1979, 205, 808–810. [Google Scholar] [CrossRef] [PubMed]
- Sogin, M.L.; Morrison, H.G.; Huber, J.A.; Welch, D.M.; Huse, S.M.; Neal, P.R.; Arrieta, J.M.; Herndl, G.J. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc. Natl. Acad. Sci. USA 2006, 103, 12115–12120. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Yarza, P.; Rapp, J.Z.; Glöckner, F.O. Expanding the world of marine bacterial and archaeal clades. Front. Microbiol. 2016, 6, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eloe, E.A.; Malfatti, F.; Gutierrez, J.; Hardy, K.; Schmidt, W.E.; Pogliano, K.; Azam, F.; Barlett, D.H. Isolation and characterization of a psychropiezophilic alphaproteobacterium. Appl. Environ. Microbiol. 2011, 77, 8145–8153. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Miyazaki, M.; Hirayama, H.; Nakagawa, S.; Querellou, J.; Godfroy, A. Isolation and physiological characterization of two novel, piezophilic, thermophilic chemolithoautotrophs from a deep-sea hydrothermal vent chimney. Environ. Microbiol. 2009, 11, 1983–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, K.; Nakamura, K.; Toki, T.; Tsunogai, U.; Miyazaki, M.; Miyazaki, J.; Hirayama, H.; Nakagawa, S.; Nunoura, T.; Horikoshi, K. Cell proliferation at 122 °C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation. Proc. Natl. Acad. Sci. 2008, 105, 10949–10954. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Ehara, M.; Saito, Y.; Yoshioka, H.; Miyazaki, M.; Saito, Y.; Miyashita, A.; Kawakami, S.; Yamaguchi, T.; Ohashi, A.; et al. A long-term cultivation of an anaerobic methane-oxidizing microbial community from deep-sea methane-seep sediment using a continuous-flow bioreactor. PLoS ONE 2014, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Landreau, M.; Duthoit, F.; Roussel, E.; Schönherr, S.; Georges, M.; Godfroy, A.; Le Blay, G. Cultivation of an immobilized (hyper)thermophilic marine microbial community in a bioreactor. FEMS Microbiol. Lett. 2016, 363, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Barone, R.; De Santi, C.; Palma Esposito, F.; Tedesco, P.; Galati, F.; Visone, M.; Di Scala, A.; de Pascale, D. Marine metagenomics, a valuable tool for enzymes and bioactive compounds discovery. Front. Mar. Sci. 2014, 1, 1–6. [Google Scholar] [CrossRef]
- Tangherlini, M.; Dell’Anno, A.; Zeigler Allen, L.; Riccioni, G.; Corinaldesi, C. Assessing viral taxonomic composition in benthic marine ecosystems: Reliability and efficiency of different bioinformatic tools for viral metagenomic analyses. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Raghavan, R.V.; Jeon, C.O.; Lee, H.J.; Priya, P.V.; Dharani, G.; Kirubagaran, R. Complex bacterial communities in the deep-sea sediments of the Bay of Bengal and volcanic Barren Island in the Andaman Sea. Mar. Genom. 2017, 31, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.J.; Kimura, N.; Yokose, H.; Otsuka, M. Heterologous production of bisucaberin using a biosynthetic gene cluster cloned from a deep sea metagenome. Mol. BioSyst. 2012, 8, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tang, X.X.; Zheng, M.; Yi, Z.W.; Xiao, X.; Qiu, Y.K.; Wu, Z. A novel indole alkaloid from deep-sea sediment metagenomic clone-derived Escherichia coli fermentation broth. J. Asian Nat. Prod. Res. 2011, 13, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Tang, X.X.; Chen, L.; Yi, Z.W.; Fang, M.J.; Wu, Z.; Qui, Y.K. Two new cytotoxic indole alkaloids from a deep-sea sediment derived metagenomic clone. Mar. Drugs 2014, 12, 2156–2163. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.C.; Zhao, D.L.; Ran, L.Y.; Mi, Z.H.; Wu, Z.Y.; Pang, X.; Zhang, X.Y.; Su, H.N.; Shi, M.; Song, X.Y.; et al. Development of a genetic system for the deep-sea psychrophilic bacterium Pseudoalteromonas sp. SM9913. Microb. Cell Fact. 2014, 13. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.W.; Jian, H.H.; Wang, F.P. pSW2, a novel low-temperature-inducible gene expression vector based on a filamentous phage of the deep-sea bacterium Shewanella piezotolerans WP3. Appl. Environ. Microbiol. 2015, 81, 5519–5526. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Escribano, J.; Alt, S.; Bibb, M. Next Generation Sequencing of Actinobacteria for the Discovery of Novel Natural Products. Mar. Drugs 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Borchert, E.; Jackson, S.A.; O’Gara, F.; Dobson, A.D.W. Diversity of natural product biosynthetic genes in the microbiome of the deep sea sponges Inflatella pellicula, Poecillastra compressa, and Stelletta normani. Front. Microbiol. 2016, 7, 1027. [Google Scholar] [CrossRef] [PubMed]
- Skinnider, M.A.; Dejong, C.A.; Rees, P.N.; Johnston, C.W.; Li, H.; Webster, A.L.H.; Wyatt, M.A.; Magarvey, N.A. Genomes to natural products PRediction Informatics for Secondary Metabolomes (PRISM). Nucleic Acids Res. 2015, 43, 9645–9662. [Google Scholar] [CrossRef] [PubMed]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Muller, R.; Wohlleben, W.; et al. AntiSMASH 3.0-A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed]
- de Jong, A.; van Hijum, S.A.; Bijlsma, J.J.; Kok, J.; Kuipers, O.P. BAGEL: A web-based bacteriocin genome mining tool. Nucl. Acids. Res. 2006, 34, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- van Heel, A.J.; de Jong, A.; Montalban-Lopez, M.; Kok, J.; Kuipers, O.P. BAGEL3: Automated identification of genes encoding bacteriocins and (non-) bactericidal posttranslationally modified peptides. Nucl. Acids. Res. 2013, 41, W448–W453. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Crossman, L.; Almeida, E.; Margassery, L.; Kennedy, J.; Dobson, A. Diverse and Abundant Secondary Metabolism Biosynthetic Gene Clusters in the Genomes of Marine Sponge Derived Streptomyces spp. Isolates. Mar. Drugs 2018, 16, 67. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Choi, Y.H.; van Wezel, G.P. Metabolic profiling as a tool for prioritizing antimicrobial compounds. J. Ind. Microbiol. Biotechnol. 2016, 43, 299–312. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Depth | Geographic Location | Compound | Molecular Class | Antimicrobial Activity Against | Ref. | |
|---|---|---|---|---|---|---|---|
| Bacteria | Marinactinospora thermotolerans (SCSIO 00652) | 3865 m | South China Sea, People’s Republic of China | Marthiapeptide A (1) | Cyclic peptide | M. luteus; S. aureus; B. subtilis; B. thuringiensis | Zhou et al., 2012 |
| Streptomyces scopuliridis (SCSIO ZJ46) | 3536 m | South China Sea, People’s Republic of China | Desotamide B (2) | Cyclic peptide | S. aureus; S. pneumoniae; MRSE shhs-E1 | Song et al., 2014 | |
| Streptomyces drozdowiczii (SCSIO 10141) | 1396 m | South China Sea, People’s Republic of China | Marfomycins A, B, E (3,4,5) | Cyclic peptide | M. luteus | Zhou et al., 2014 | |
| Streptomyces sp. (SCSIO 01127) | 1350 m | South China Sea, People’s Republic of China | Lobophorin F (6) | Spirotetronate poliketides | S. aureus; E. faecalis | Niu et al., 2011 | |
| Streptomyces sp. (12A35) | 2134 m | South China Sea, People’s Republic of China | Lobophorin H (7) | Spirotetronate poliketides | B. subtilis | Pan et al., 2013 | |
| Verrucosispora sp. (AB 18-032) | 289 m | Japanese Sea | Abyssomicin C (8) | Spirotetronate poliketides | MRSA; vancomycin-resistant S. aureus | Bister et al., 2004 | |
| Streptomyces niveus (SCSIO 3406) | 3536 m | South China Sea, People’s Republic of China | Marfuraquinocins A, C, D (9,10,11) | Sesquiterpene derivative | S.aureus; MRSE shhs-E1 | Song et al., 2013 | |
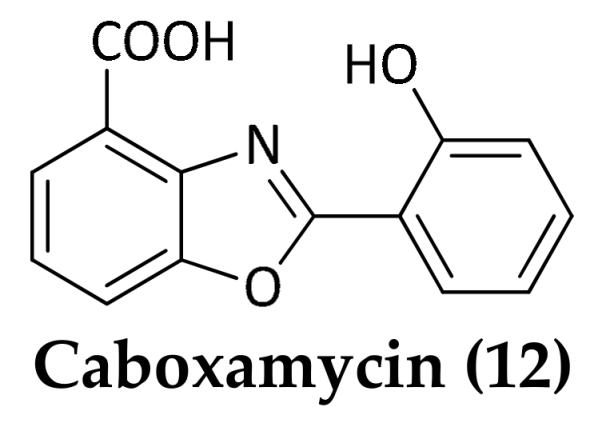
| Streptomyces sp. (NTK 937) | 3814 m | Saharan debris flow near the Canary Islands | Caboxamycin (12) | Alkaloid | B. subtilis; S. lentus; S. epidermidis | Hohmann et al., 2009 | |
| Fungi | Emericella sp. (SCSIO 05240) | 3258 m | South China Sea, People’s Republic of China | Emerixanthones A, B, C, D (13,14,15,16) | Xanthone-derivative | E. coli; K. pneumoniae; S. aureus; E. faecalis; A. baumannii; A. hydrophila | Fredimoses et al., 2014 |
| Engyodontium album (DFFSCS021) | 3739 m | South China Sea, People’s Republic of China | Engyodontiumone H (17) | Xanthone-derivative | E. coli; B. subtilis | Yao et al., 2014 | |
| Spiromastix sp. | 2869 m | South Atlantic Ocean | Spiromastixones A, B, C, D, E, F, G, H, K, L, M, N, O (18,19,20,21,22,23,24,25,26,27,28,29,30,31,32) | Depsidone analogs | S. aureus; B. thuringiensis; B. subtilis; MRSA; MRSE; E. faecalis; E. faecium | Niu et al., 2014 | |
| Penicillium sp. (F23-2) | 5080 m | Chinese Sea, People’s Republic of China | Penicyclones A, B, C, D, E, F (33,34,35,36,37) | Ambuic acid analogs | S.aureus | Guo et al., 2015 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tortorella, E.; Tedesco, P.; Palma Esposito, F.; January, G.G.; Fani, R.; Jaspars, M.; De Pascale, D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs 2018, 16, 355. https://doi.org/10.3390/md16100355
Tortorella E, Tedesco P, Palma Esposito F, January GG, Fani R, Jaspars M, De Pascale D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Marine Drugs. 2018; 16(10):355. https://doi.org/10.3390/md16100355
Chicago/Turabian StyleTortorella, Emiliana, Pietro Tedesco, Fortunato Palma Esposito, Grant Garren January, Renato Fani, Marcel Jaspars, and Donatella De Pascale. 2018. "Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives" Marine Drugs 16, no. 10: 355. https://doi.org/10.3390/md16100355
APA StyleTortorella, E., Tedesco, P., Palma Esposito, F., January, G. G., Fani, R., Jaspars, M., & De Pascale, D. (2018). Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Marine Drugs, 16(10), 355. https://doi.org/10.3390/md16100355