Novel Bioactive Peptides from Meretrix meretrix Protect Caenorhabditis elegans against Free Radical-Induced Oxidative Stress through the Stress Response Factor DAF-16/FOXO

 ,
,
Abstract
:1. Introduction
2. Results
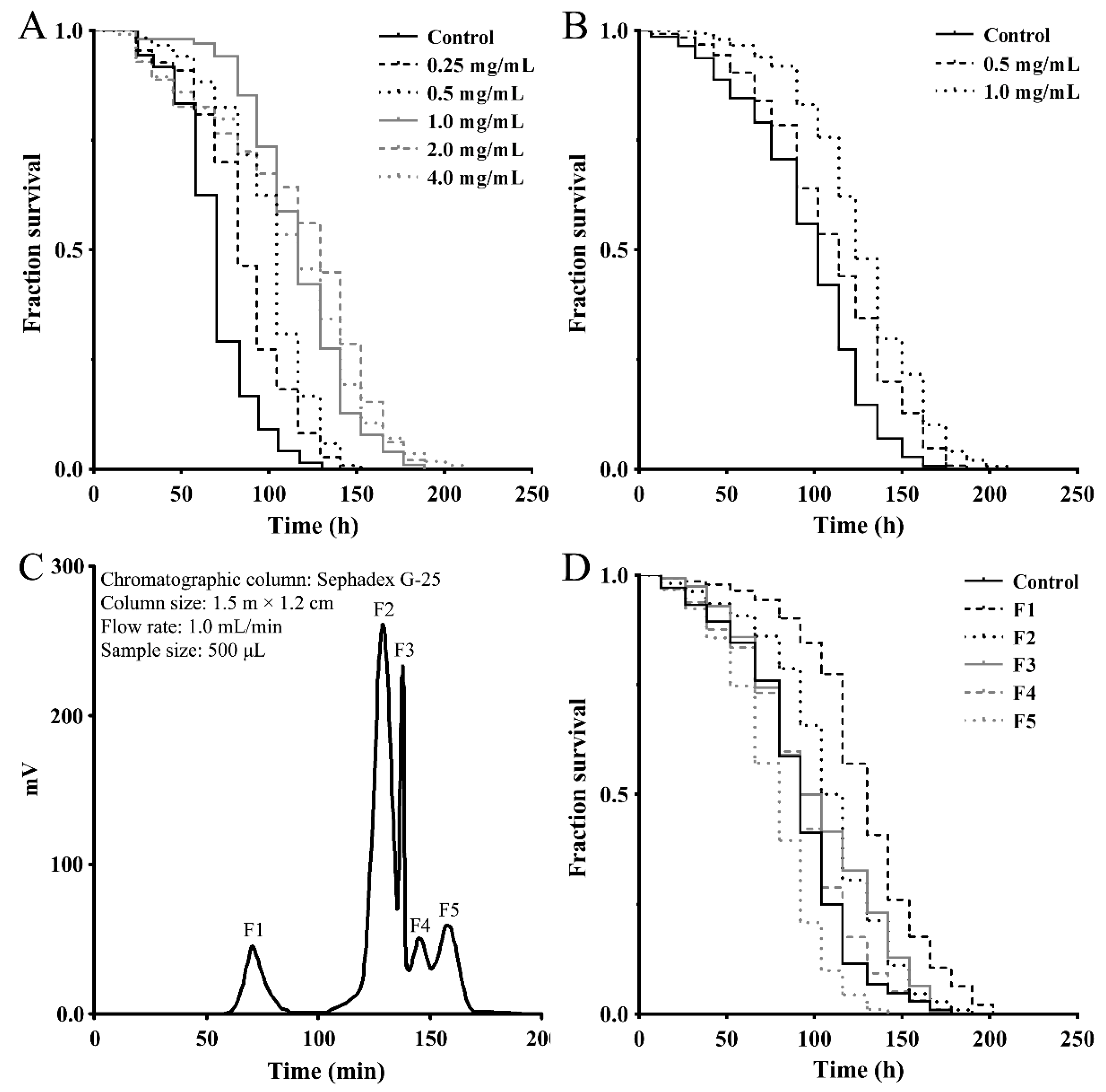
2.1. Separation of Antioxidant Peptide Fractions from M. meretrix Peptide-Rich Extract
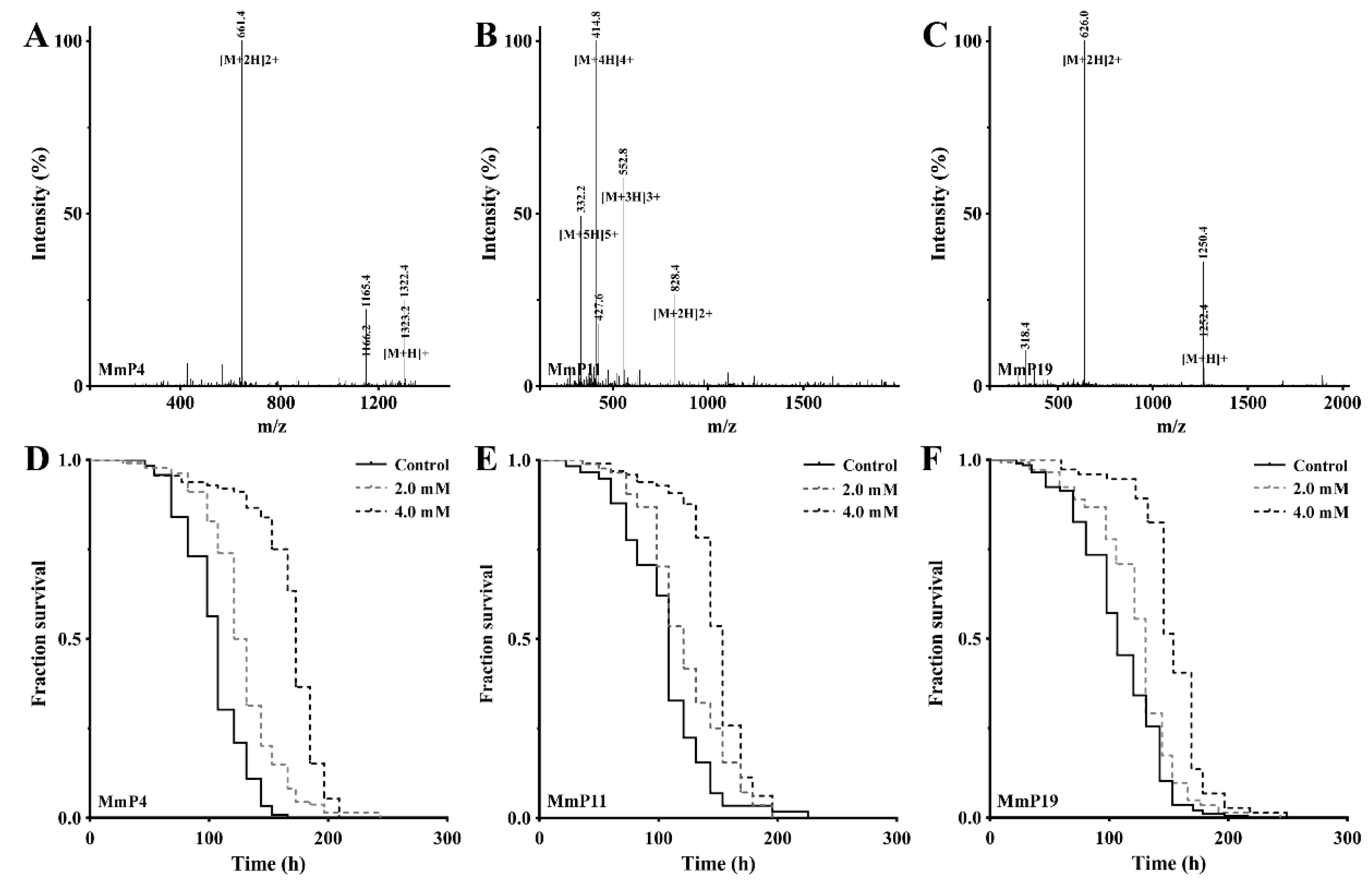
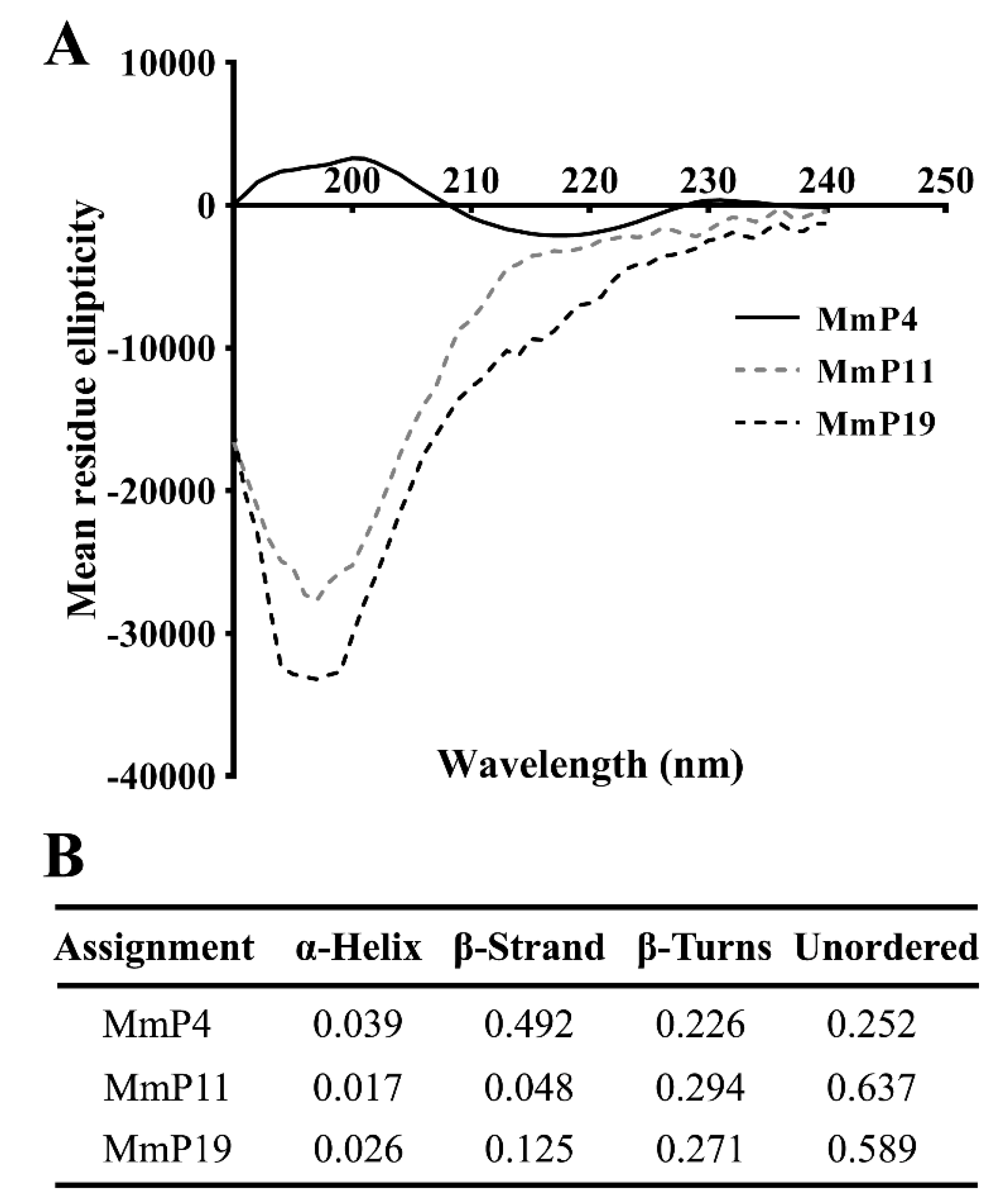
2.2. Identification, Synthesis and Characterization of Antioxidant M. meretrix Peptides
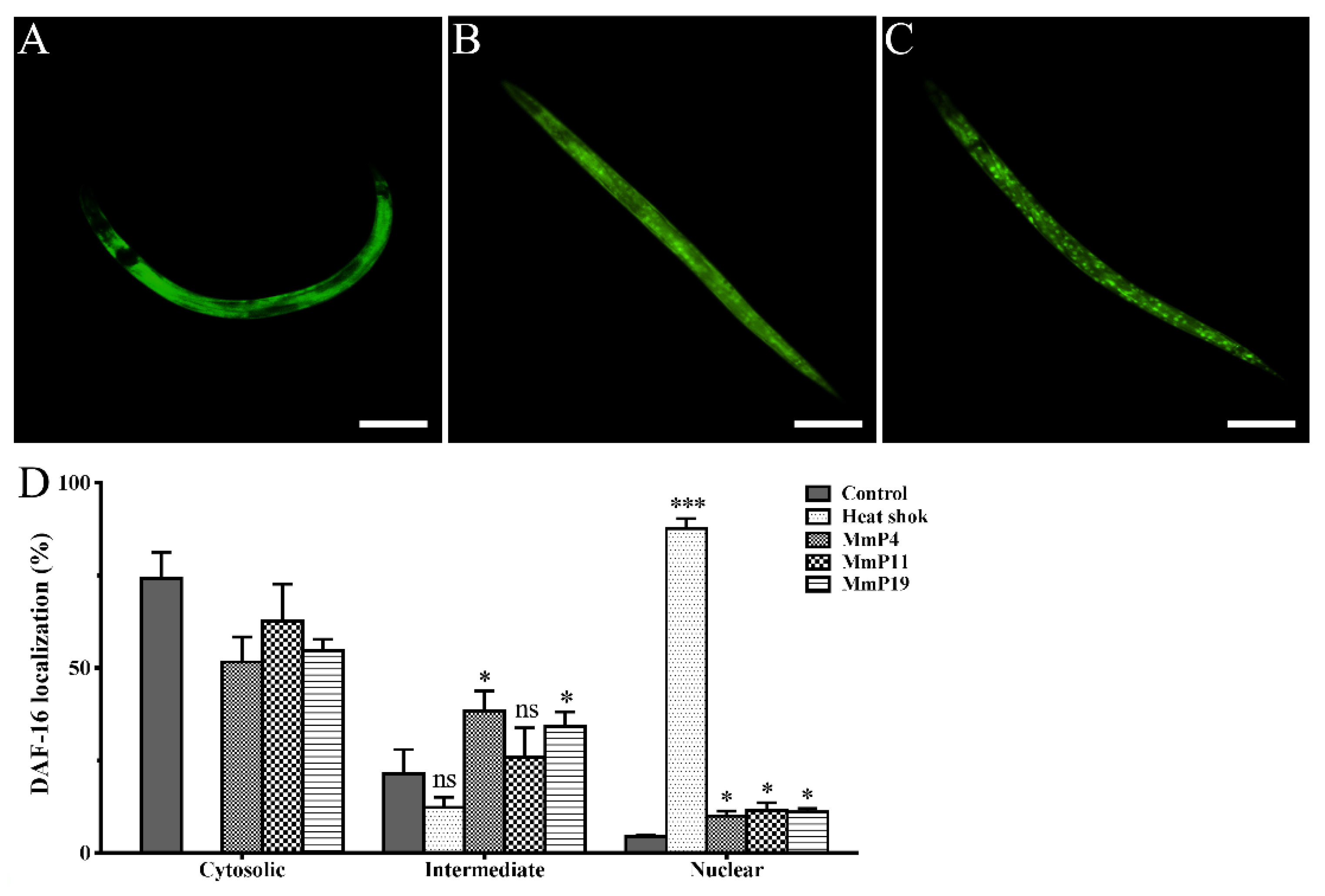
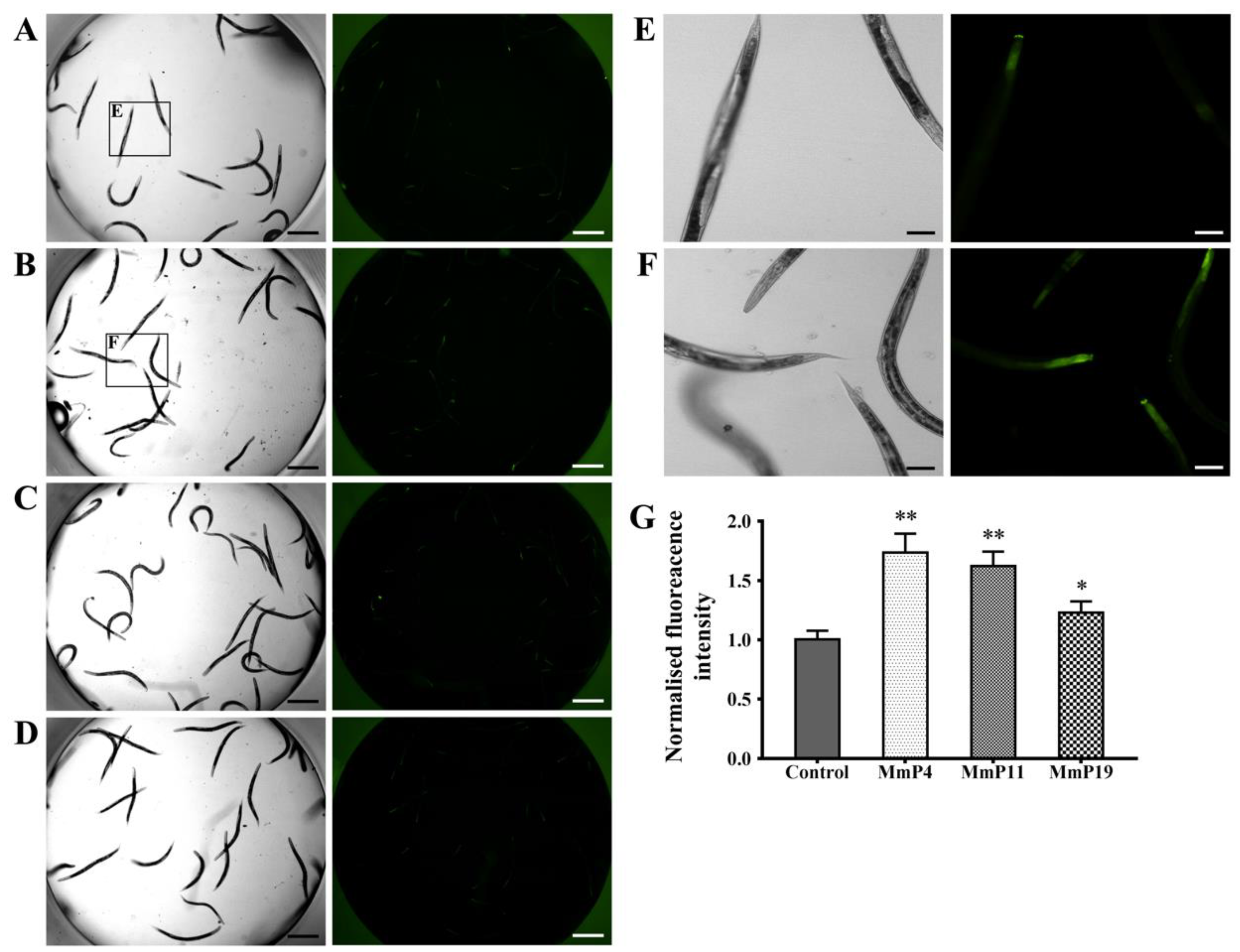
2.3. Effects of Antioxidant M. meretrix Peptides on Subcellular Localization of the Transcription Factor DAF-16
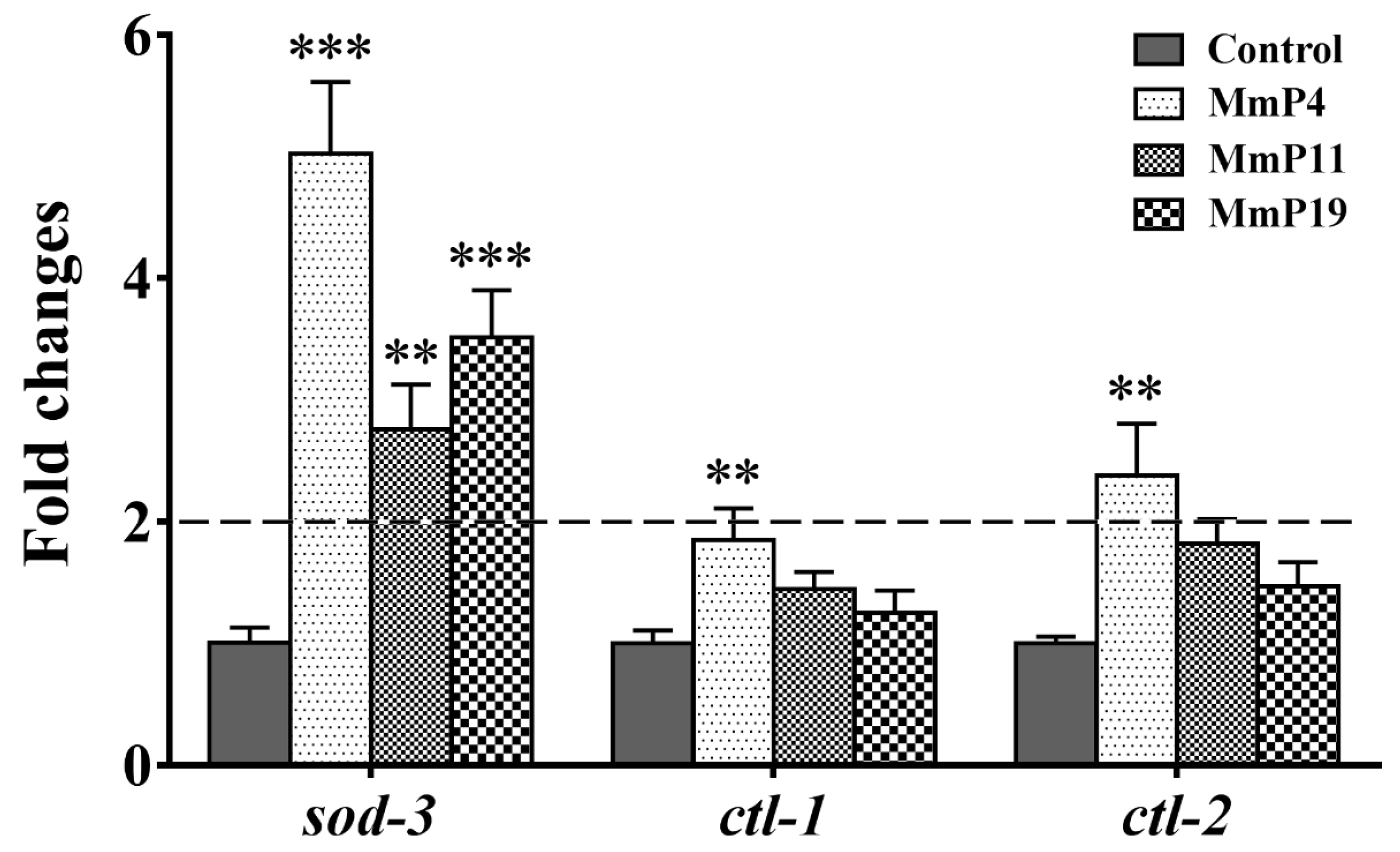
2.4. Effects of Antioxidant M. meretrix Peptides on the Expression of Oxidative Stress Related Genes
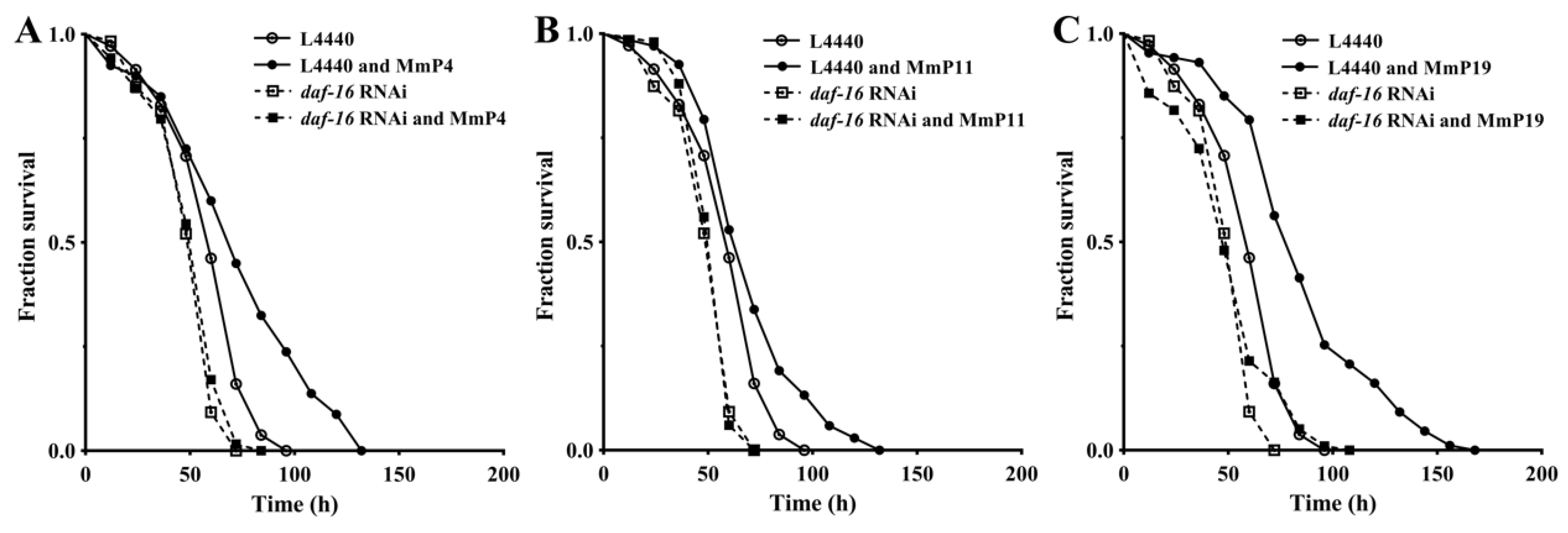
2.5. Dependency of Antioxidant Activity of M. meretrix Peptides on Transcription Factor DAF-16
3. Discussion
4. Materials and Methods
4.1. Chemicals and Materials
4.2. Nematode and Bacterial Strains
4.3. Preparation of Natural Peptides
4.4. Paraquat Resistance Assay
4.5. Identification and Synthesis of Peptides
4.6. Circular Dichroism Spectroscopy
4.7. Nuclear Localization Assay of Transcription Factor DAF-16
4.8. Determination of SOD-3 Expression Levels
4.9. Real-time PCR Analysis
4.10. RNA Interference Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mima, A. Inflammation and oxidative stress in diabetic nephropathy: New insights on its inhibition as new therapeutic targets. J. Diabetes Res. 2013, 6, 248563. [Google Scholar] [CrossRef] [PubMed]
- Angelova, P.R.; Abramov, A.Y. Functional role of mitochondrial reactive oxygen species in physiology. Free Radic. Biol. Med. 2016, 100, 81–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viktorinova, A.; Svitekova, K.; Stecova, A.; Krizko, M. Relationship between selected oxidative stress markers and lipid risk factors for cardiovascular disease in middle-aged adults and its possible clinical relevance. Clin. Biochem. 2016, 9, 868–872. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, F.; Pupo, G.; Giraldo, E.; Badìa, M.C.; Monllor, P.; Lloret, A.; Schininà, M.E.; Giorgi, A.; Cini, C.; Tramutola, A.; et al. Oxidative signature of cerebrospinal fluid from mild cognitive impairment and Alzheimer disease patients. Free Radic. Biol. Med. 2016, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Godic, A.; Poljšak, B.; Adamic, M.; Dahmane, R. The role of antioxidants in skin cancer prevention and treatment. Oxid. Med. Cell. Longev. 2014, 860479. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, D.M.; Asaithamby, A.; Bailey, S.M.; Costes, S.V.; Doetsch, P.W.; Dynan, W.S.; Kronenberg, A.; Rithidech, K.N.; Saha, J.; Snijders, A.M.; Werner, E.; et al. Understanding cancer development processes after HZE-particle exposure: Roles of ROS, DNA damage repair and inflammation. Radiat. Res. 2015, 183, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Görlach, A.; Dimova, E.Y.; Petry, A.; Martínez-Ruiz, A.; Hernansanz-Agustín, P.; Rolo, A.P.; Palmeira, C.M.; Kietzmann, T. Reactive oxygen species, nutrition, hypoxia and diseases: Problems solved? Redox Biol. 2015, 6, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Van Raamsdonk, J.M.; Hekimi, S. Reactive oxygen species and aging in Caenorhabditis elegans: Causal or casual relationship? Antioxid. Redox Signal. 2010, 13, 1911–1953. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Tunnacliffe, A. Cryptobiosis, aging and cancer: Yin-yang balancing of signaling networks. Rejuv. Res. 2006, 9, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Himaya, S.W.; Ryu, B.; Ngo, D.H.; Kim, S.K. Peptide isolated from Japanese flounder skin gelatin protects against cellular oxidative damage. J. Agric. Food Chem. 2012, 60, 9112–9119. [Google Scholar] [CrossRef] [PubMed]
- Alashi, A.M.; Blanchard, C.L.; Mailer, R.J.; Agboola, S.O.; Mawson, A.J.; He, R.; Girgih, A.; Aluko, R.E. Antioxidant properties of Australian canola meal protein hydrolysates. Food Chem. 2014, 146, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Ngoungoure, V.L.; Schluesener, J.; Moundipa, P.F.; Schluesener, H. Natural polyphenols binding to amyloid: A broad class of compounds to treat different human amyloid diseases. Mol. Nutr. Food Res. 2015, 59, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Feng, L.; Bao, W.R.; Ma, D.L.; Leung, C.H.; Nie, S.P.; Han, Q.B. Structure characterization and immunomodulating effects of polysaccharides isolated from Dendrobium officinale. J. Agric. Food Chem. 2016, 64, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A. review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Li, H.; Ding, F.; Xiao, L.; Shi, R.; Wang, H.; Han, W.; Huang, Z. Food-derived antioxidant polysaccharides and their pharmacological potential in neurodegenerative diseases. Nutrients 2017, 9, 778. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; He, R.; Malomo, S.A.; Offengenden, M.; Wu, J.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Fan, X.; Bai, L.; Zhu, L.; Yang, L.; Zhang, X. Marine algae-derived bioactive peptides for human nutrition and health. J. Agric. Food Chem. 2014, 62, 9211–9222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Huang, Y.; Qin, C.; Liang, M.; Mao, X.; Li, S.; Zou, Y.; Jia, W.; Li, H.; Ma, C.W.; et al. Bioactive peptides from Angelica sinensis protein hydrolyzate delay senescence in Caenorhabditis elegans through antioxidant activities. Oxid. Med. Cell. Longev. 2016, 2016, 8956981. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Cheng, Q.; Peng, Q.; Yu, X.; Yin, X.; Liang, M.; Ma, C.W.; Huang, Z.; Jia, W. Antioxidant peptides derived from the hydrolyzate of purple sea urchin (Strongylocentrotus nudus) gonad alleviate oxidative stress in Caenorhabditis elegans. J. Funct. Foods 2018, 48, 594–604. [Google Scholar] [CrossRef]
- Wang, C.; Liu, M.; Cheng, L.; Wei, J.; Wu, N.; Zheng, L.; Lin, X. A novel polypeptide from Meretrix meretrix Linnaeus inhibits the growth of human lung adenocarcinoma. Exp. Biol. Med. 2012, 237, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, J.; Wu, N.; Liu, M.; Wang, C.; Zhang, Y.; Wang, F.; Liu, H.; Lin, X. Mere15, a novel polypeptide from Meretrix meretrix, inhibits adhesion, migration and invasion of human lung cancer A549 cells via down-regulating MMPs. Pharm. Biol. 2013, 51, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhao, X.; Zhao, J.; Xiao, L.; Liu, H.; Wang, C.; Cheng, L.; Wu, N.; Lin, X. Induction of apoptosis, G0/G1 phase arrest and microtubule disassembly in K562 leukemia cells by Mere15, a novel polypeptide from Meretrix meretrix Linnaeus. Mar. Drugs 2012, 10, 2596–2607. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, H.; Qian, J.; He, Y.; Zheng, J.; Lu, Z.; Xu, Z.; Shi, J. Structural and immunological activity characterization of a polysaccharide isolated from Meretrix meretrix Linnaeus. Mar. Drugs 2015, 14, 6. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhao, S.; Yu, F.; Yang, Z.; Ding, G. Protective effects and mechanism of Meretrix meretrix oligopeptides against nonalcoholic fatty liver disease. Mar. Drugs 2017, 15, 31. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wang, J.; Yu, F.; Tang, Y.; Ding, G.; Yang, Z.; Sun, Y. Protective effect of Meretrix meretrix oligopeptides on high-fat-diet-induced non-alcoholic fatty liver disease in mice. Mar. Drugs 2018, 16, 39. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, F.; Guo, W.; Zhang, J.; Xiao, L.; Li, H.; Jia, W.; Huang, Z. Caenorhabditis elegans in Chinese medicinal studies: Making the case for aging and neurodegeneration. Rejuv. Res. 2014, 17, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Hong, R.; He, S.; Liu, G.; Huang, Z.; Zheng, Y. Polyproline II structure is critical for the enzyme protective function of soybean Em (LEA1) conserved domains. Biotechnol. Lett. 2011, 33, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Wallace, B.A. Protein secondary structure analyses from circular dichroism spectroscopy: Methods and reference databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, T.; Biggs, W.H.; Tieu, D.; Boyer, A.D.; Varki, N.M.; Cavenee, W.K.; Arden, K.C. Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc. Natl. Acad. Sci. USA 2004, 101, 2975–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.W.; Mukhopadhyay, A.; Dixit, B.L.; Raha, T.; Green, M.R.; Tissenbaum, H.A. Identification of direct DAF-16 targets controlling longevity, metabolism and diapause by chromatin immunoprecipitation. Nat. Genet. 2006, 38, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Li, W.H.; Shi, Y.C.; Chang, C.H.; Huang, C.W.; Hsiu-Chuan Liao, V. Selenite protects Caenorhabditis elegans from oxidative stress via DAF-16 and TRXR-1. Mol. Nutr. Food Res. 2014, 58, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Evason, K.; Collins, J.J.; Huang, C.; Hughes, S.; Kornfeld, K. Valproic acid extends Caenorhabditis elegans lifespan. Aging Cell 2008, 7, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, R.; Li, H.; Xiang, Y.; Xiao, L.; Hu, M.; Ma, F.; Ma, C.W.; Huang, Z. Antioxidant and neuroprotective effects of Dictyophora indusiata polysaccharide in Caenorhabditis elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.T.; McCarroll, S.A.; Bargmann, C.I.; Fraser, A.; Kamath, R.S.; Ahringer, J.; Li, H.; Kenyon, C. Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 2003, 424, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Libina, N.; Berman, J.R.; Kenyon, C. Tissue-specific activities of C. elegans DAF-16 in the regulation of lifespan. Cell 2003, 115, 489–502. [Google Scholar] [CrossRef]
- Chandrasekaran, A.; Idelchik, M.D.; Melendez, J.A. Redox control of senescence and age-related disease. Redox Biol. 2017, 11, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Pan, N.; Xiong, S.; Zou, S.; Li, H.; Xiao, L.; Cao, Z.; Tunnacliffe, A.; Huang, Z. Inhibition of polyglutamine-mediated proteotoxicity by Astragalus membranaceus polysaccharide through DAF-16/FOXO transcription factor in Caenorhabditis elegans. Biochem. J. 2012, 441, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.L.; Murphy, C.T.; Kenyon, C. Regulation of aging and age-related disease by DAF-16 and heat-shock factor. Science 2003, 300, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.M.; Vural, S.; King, B.R.; Guda, C. Mining for class-specificmotifs in protein sequence classification. BMC Bioinform. 2013, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Lee, J.H.; Kim, D.H.; Kim, K.T.; Lee, G.W.; Choi, S.J.; Chang, P.S.; Paik, H.D. Antioxidative and nitric oxide scavenging activity of branched-chain amino acids. Food Sci. Biotechnol. 2015, 24, 1555–1558. [Google Scholar] [CrossRef]
- Dai, D.F.; Chiao, Y.A.; Marcinek, D.J.; Szeto, H.H.; Rabinovitch, P.S. Mitochondrial oxidative stress in aging and healthspan. Longev. Healthspan 2014, 3, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco-Ayala, T.; Andérica-Romero, A.C.; Pedraza-Chaverri, J. New insights into antioxidant strategies against paraquat toxicity. Free Radic. Res. 2014, 48, 623–640. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, J.; Yan, Y.; Chi, M.; Chen, W.; Sun, P.; Qin, S. The protective effect of C-phycocyanin on paraquat-induced acute lung injury in rats. Environ. Toxicol. Pharmacol. 2011, 32, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.B.; Lin, C.C.; Chiou, J.F.; Mau, S.Y.; Liu, T.Z.; Chen, C.H. Treatment of acute paraquat intoxication using recommended megadose of vitamin C: A reappraisal. Free Radic. Res. 2013, 47, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.I.; Pincus, Z.; Slack, F.J. Longevity and stress in Caenorhabditis elegans. Aging 2011, 3, 733–753. [Google Scholar] [CrossRef] [PubMed]
- Yanase, S.; Yasuda, K.; Ishii, N. Adaptive responses to oxidative damage in three mutants of Caenorhabditis elegans (age-1, mev-1 and daf-16) that affect life span. Mech. Ageing Dev. 2002, 123, 1579–1587. [Google Scholar] [CrossRef]
- Wang, B.; Gong, Y.; Li, Z.; Yu, D.; Chi, C.; Ma, J. Isolation and characterisation of five novel antioxidant peptides from ethanol-soluble proteins hydrolysate of spotless smoothhound (Mustelus griseus) muscle. J. Funct. Foods 2014, 6, 176–185. [Google Scholar] [CrossRef]
- Zhang, L.; Jie, G.; Zhang, J.; Zhao, B. Significant longevity-extending effects of EGCG on Caenorhabditis elegans under stress. Free Radic. Biol. Med. 2009, 46, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Kamath, R.S.; Martinez-Campos, M.; Zipperlen, P.; Fraser, A.G.; Ahringer, J. Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans. Genome Biol. 2000, 2, research0002-1. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide ID | Amino Acid Sequence | Observed m/z | Charge Number | Mascot Score | MW (Da) | Coding Gene | Purity a |
|---|---|---|---|---|---|---|---|
| MmP1 | LCLTPEMVSPTP | 644.32 | 2 | 56.27 | 1286.63 | gi: 299825528 | 99.69% |
| MmP2 | THQPCTSPSRLCYLCML | 656.96 | 3 | 50.21 | 1967.87 | gi: 299827727 | 99.05% |
| MmP3 | MSPSKLCCPCTLP | 690.30 | 2 | 51.58 | 1378.61 | gi: 238686243 | 99.60% |
| MmP4 | LSDRLEETGGASS | 661.31 | 2 | 60.55 | 1320.62 | gi:299825499 | 95.37% |
| MmP5 | AGFAGDDAPRAVFPS | 739.35 | 2 | 82.78 | 1476.7 | gi: 299825528 | 98.91% |
| MmP6 | GIECLGHYLCHL | 679.33 | 2 | 53.86 | 1356.63 | gi: 299827928 | 96.21% |
| MmP7 | YTKFVVESMMP | 682.33 | 2 | 51.38 | 1362.62 | gi: 299827824 | 95.39% |
| MmP8 | TLNICVMLVSVNC | 712.85 | 2 | 52.34 | 1423.69 | gi: 570035797 | 96.65% |
| MmP9 | KSYGCEKGTGCLLL | 736.36 | 2 | 60.21 | 1470.72 | gi: 299825528 | 97.13% |
| MmP10 | QPCTLPSRPCSLCT | 753.35 | 2 | 54.9 | 1504.68 | gi: 299825528 | 98.95% |
| MmP11 | KEGCREPETEKGHR | 828.41 | 2 | 61.14 | 1654.78 | gi: 299825499 | 96.51% |
| MmP12 | LSDRLEETGGASSIQHE | 610.29 | 3 | 54.51 | 1827.86 | gi: 299825499 | 95.45% |
| MmP13 | RPVCERNTRKPSMTC | 889.43 | 2 | 71.39 | 1776.85 | gi: 299825499 | 99.90% |
| MmP14 | GGEINCRIN | 488.24 | 2 | 55.59 | 974.46 | gi: 299827820 | 97.75% |
| MmP15 | ANIHGWCVKGFEHDL | 440.22 | 4 | 57.89 | 1756.8 | gi: 299827878 | 98.57% |
| MmP16 | GFAGDDAPRAVFPSIVG | 838.42 | 2 | 67.3 | 1674.84 | gi: 299825528 | 99.06% |
| MmP17 | EYEINCRINCR | 706.83 | 2 | 52.92 | 1411.63 | gi: 299825354 | 98.11% |
| MmP18 | VSYRAETCHALL | 681.85 | 2 | 59.75 | 1361.68 | gi: 570035646 | 99.06% |
| MmP19 | IVTNWDDMEK | 625.79 | 2 | 77.36 | 1249.56 | gi:299827878 | 97.21% |
| MmP20 | LPMVITCP | 437.23 | 2 | 55.68 | 872.45 | gi:299827878 | 96.39% |
| MmP21 | LLTMAPVCA | 459.74 | 2 | 66.76 | 917.47 | gi: 299827727 | 97.56% |
| MmP22 | CRNSVSRTAAPCL | 689.33 | 2 | 63.83 | 1376.67 | gi: 238685652 | 97.46% |
| MmP23 | LGECLLCPG | 452.72 | 2 | 61.57 | 903.42 | gi: 299825408 | 99.44% |
| MmP24 | MATYMAGVLKVSNM | 774.38 | 2 | 52.99 | 1546.72 | gi: 299827727 | 98.38% |
| MmP25 | VECWGIECLGH | 639.26 | 2 | 56.98 | 1276.52 | gi: 299827928 | 98.87% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, W.; Peng, Q.; Su, L.; Yu, X.; Ma, C.W.; Liang, M.; Yin, X.; Zou, Y.; Huang, Z. Novel Bioactive Peptides from Meretrix meretrix Protect Caenorhabditis elegans against Free Radical-Induced Oxidative Stress through the Stress Response Factor DAF-16/FOXO. Mar. Drugs 2018, 16, 444. https://doi.org/10.3390/md16110444
Jia W, Peng Q, Su L, Yu X, Ma CW, Liang M, Yin X, Zou Y, Huang Z. Novel Bioactive Peptides from Meretrix meretrix Protect Caenorhabditis elegans against Free Radical-Induced Oxidative Stress through the Stress Response Factor DAF-16/FOXO. Marine Drugs. 2018; 16(11):444. https://doi.org/10.3390/md16110444
Chicago/Turabian StyleJia, Weizhang, Qiong Peng, Linnan Su, Xuesong Yu, Chung Wah Ma, Ming Liang, Xiquan Yin, Yongdong Zou, and Zebo Huang. 2018. "Novel Bioactive Peptides from Meretrix meretrix Protect Caenorhabditis elegans against Free Radical-Induced Oxidative Stress through the Stress Response Factor DAF-16/FOXO" Marine Drugs 16, no. 11: 444. https://doi.org/10.3390/md16110444
APA StyleJia, W., Peng, Q., Su, L., Yu, X., Ma, C. W., Liang, M., Yin, X., Zou, Y., & Huang, Z. (2018). Novel Bioactive Peptides from Meretrix meretrix Protect Caenorhabditis elegans against Free Radical-Induced Oxidative Stress through the Stress Response Factor DAF-16/FOXO. Marine Drugs, 16(11), 444. https://doi.org/10.3390/md16110444