Marine Spongin: Naturally Prefabricated 3D Scaffold-Based Biomaterial

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
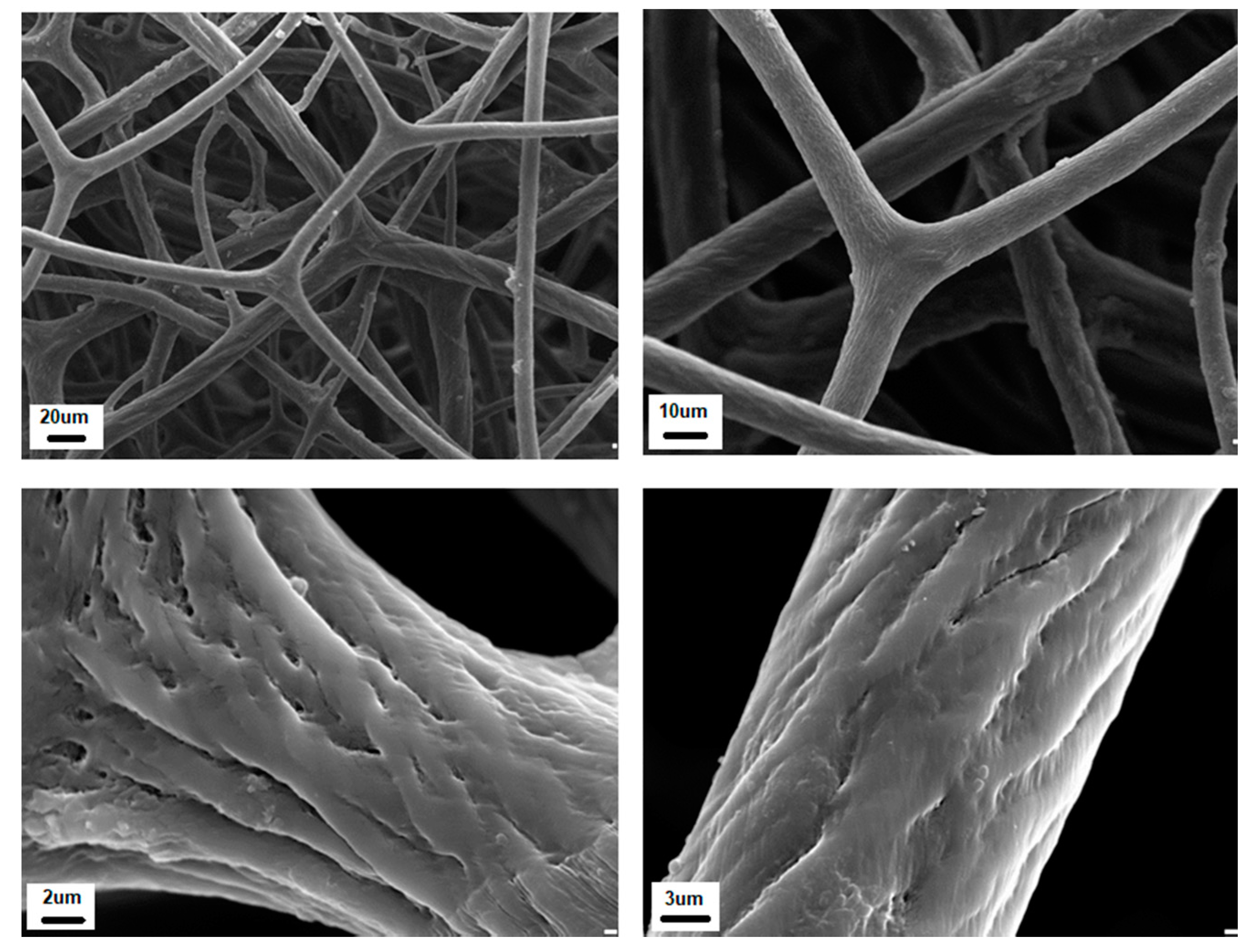{kind=link}
{kind=link}
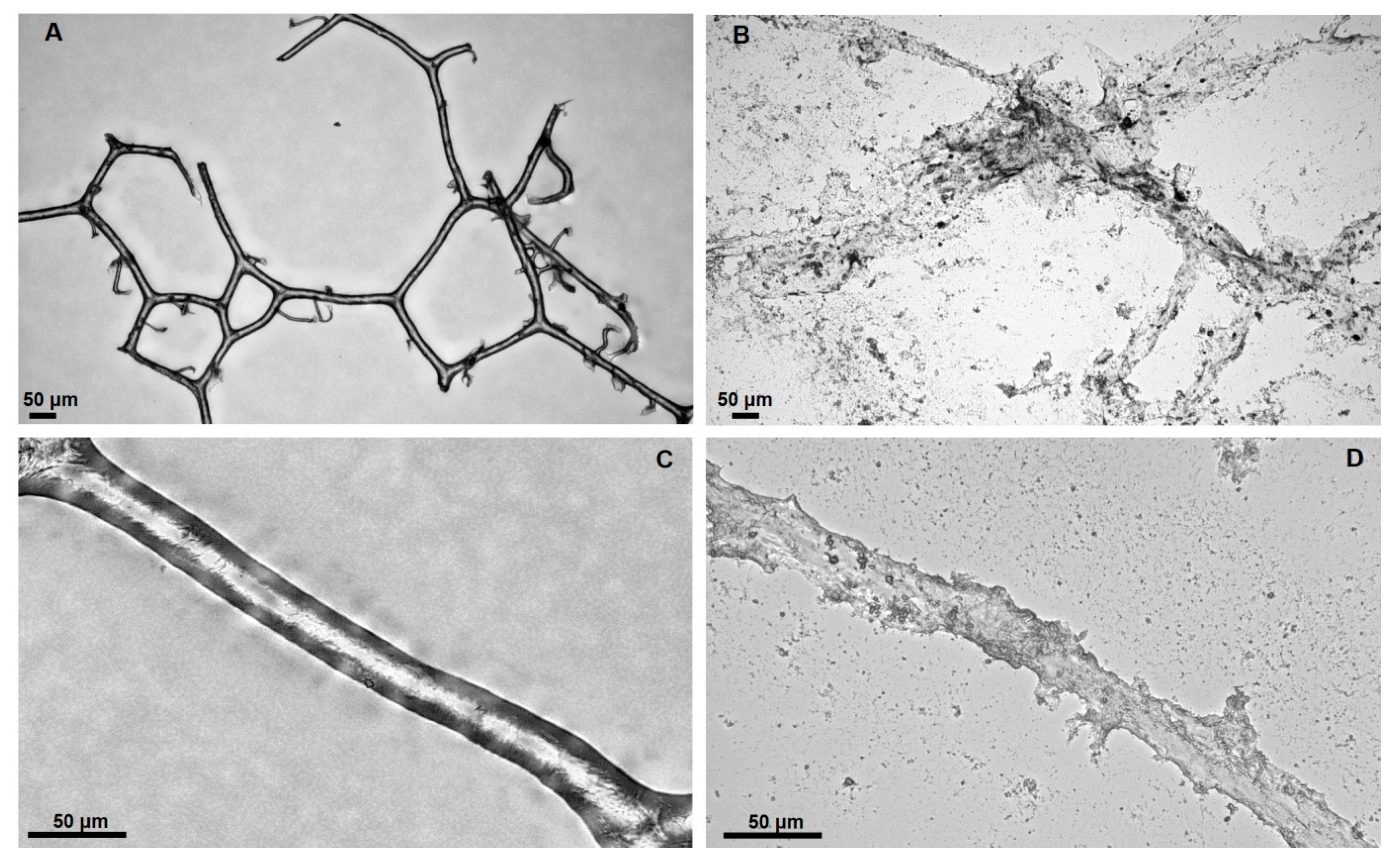{kind=link}
{kind=link}
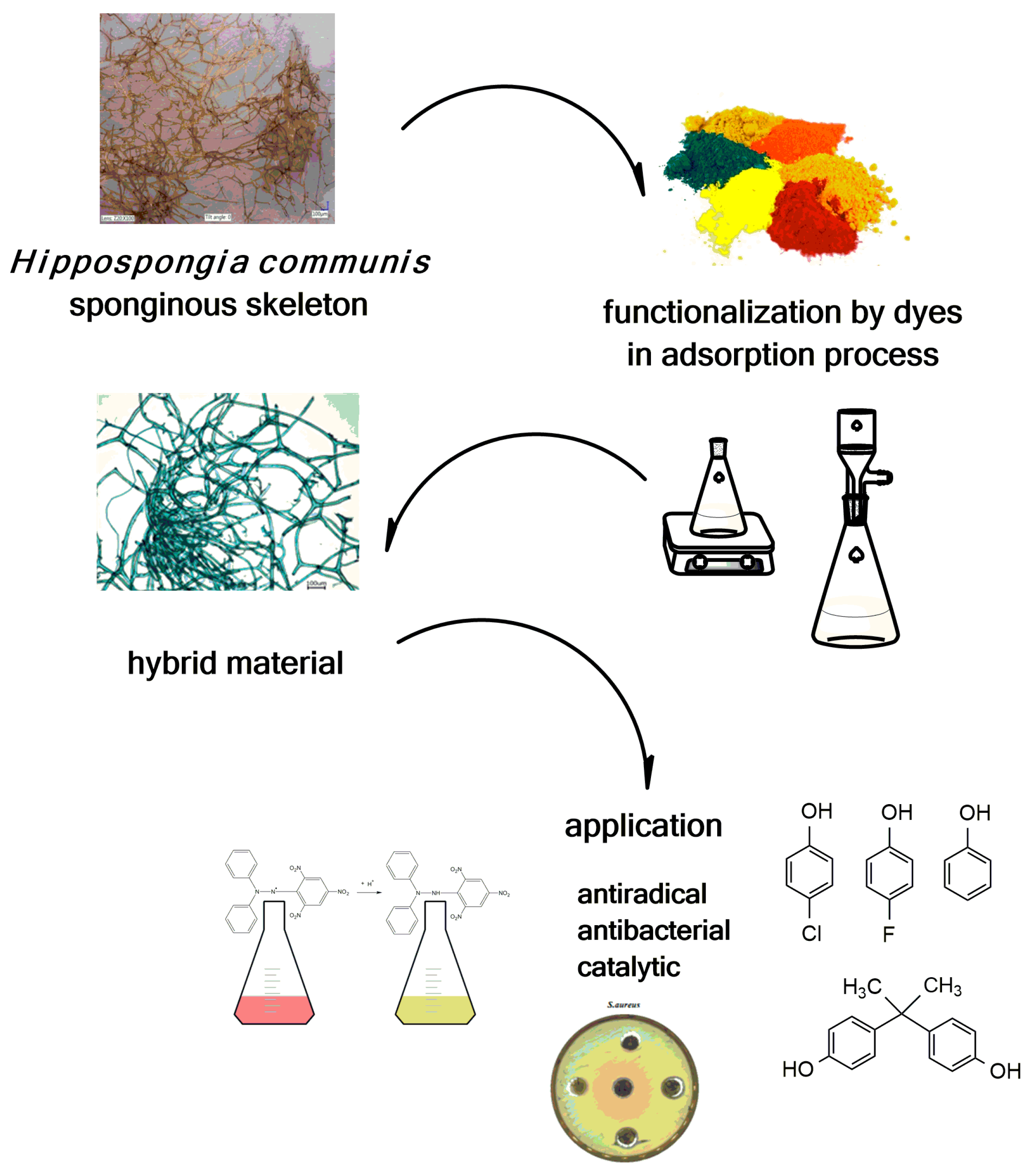{kind=link}
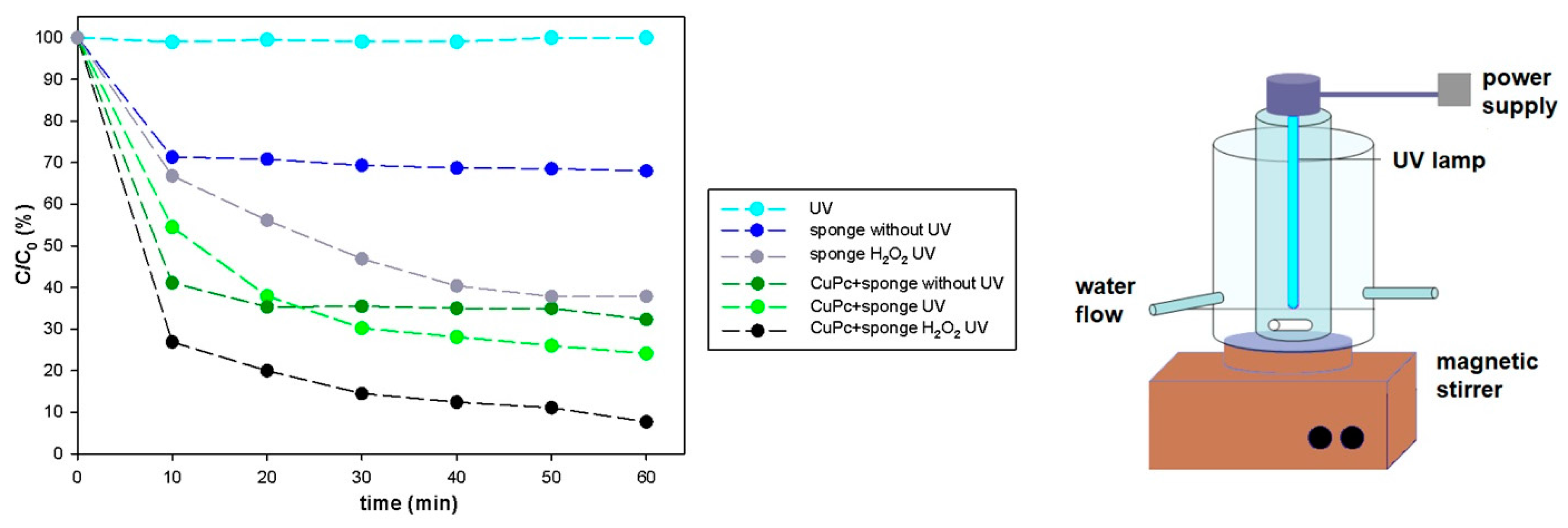{kind=link}
{kind=link}
Abstract
:1. Introduction
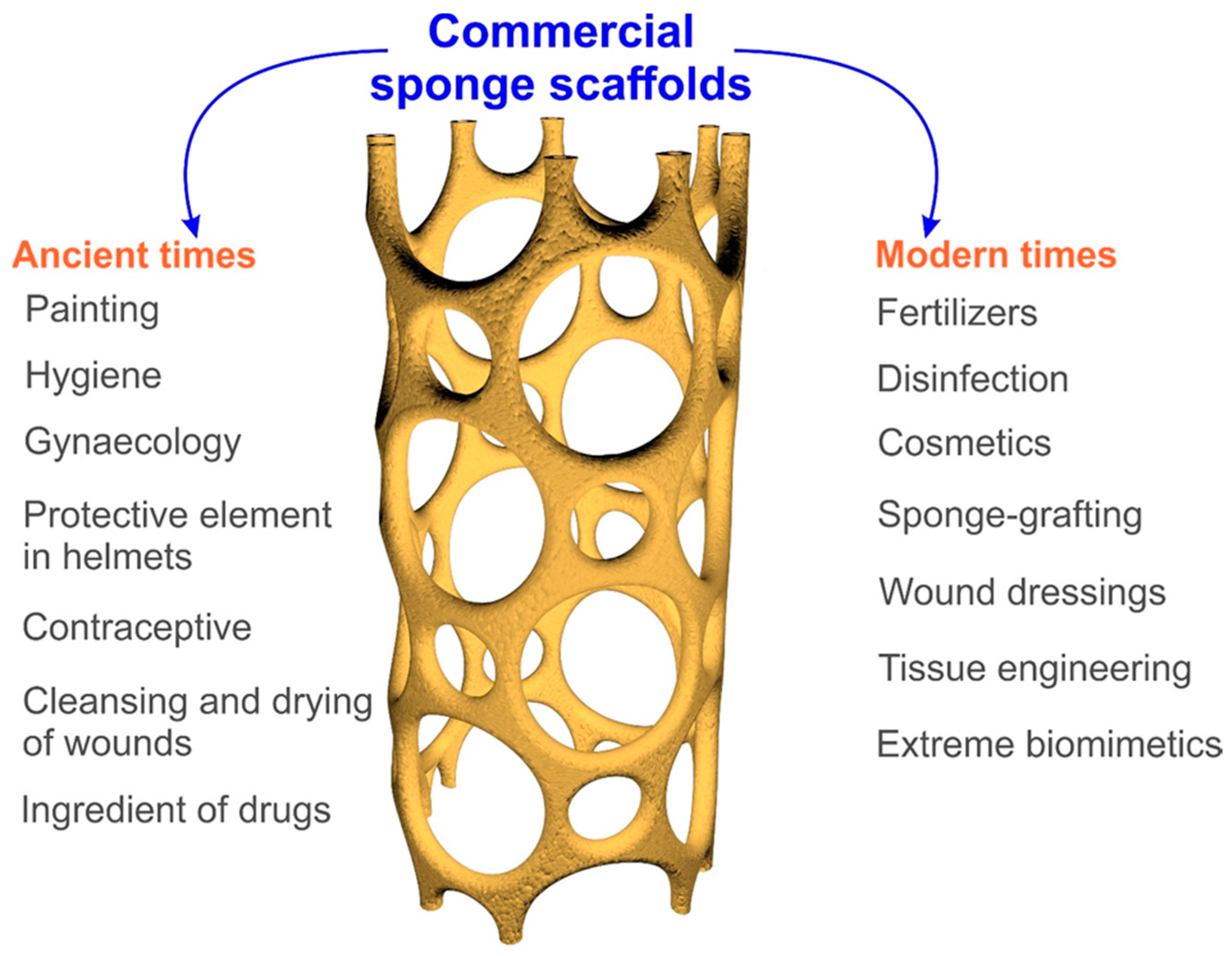
2. From Commercial Sponges to Modern 3D Spongin Scaffolds: Brief History
3. The Economics of the Commercial Sponge Industry
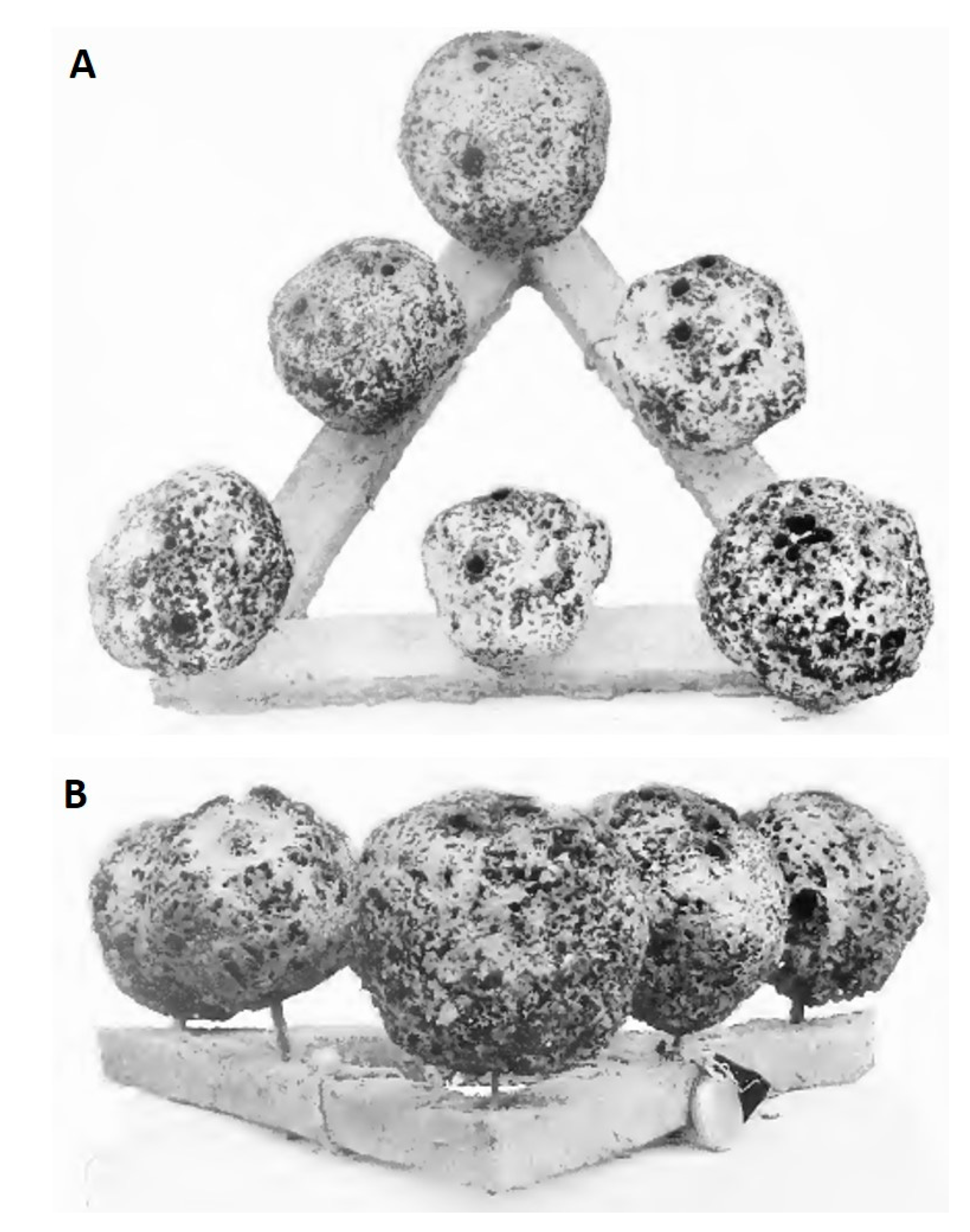
4. Marine Farming of Bath Sponges and Spongin as Renewable Biomaterial
5. Structure, Chemistry, and Properties of Spongin
6. Material Properties of 3D Spongin Scaffolds
7. 3D Spongin Scaffolds and Tissue Engineering
8. 3D Spongin Scaffolds as a Support for Dye Immobilization
9. Spongin and Extreme Biomimetics
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Feuda, R.; Dohrmann, M.; Pett, W.; Philippe, H.; Rota-Stabelli, O.; Lartillot, N.; Worheide, G.; Pisani, D. Improved modeling of compositional heterogeneity supports sponges as sister to all other animals. Curr. Biol. 2017, 27, 3864–3870. [Google Scholar] [CrossRef] [PubMed]
- World Porifera Database. Available online: http://www.marinespecies.org/porifera/ (accessed on 28 January 2018).
- Morrow, C.; Cárdenas, P. Proposal for a revised classification of the Demospongiae (Porifera). Front. Zool. 2015, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.E. Tabular View of the Primary Divisions of the Animal Kingdom, Intended to Serve As an Outline of an Elementary Course of Recent Zoology (Caino-Zoology); Walton and Maberly: London, UK, 1861; ISBN 1236783921. [Google Scholar]
- Minchin, E. Sponges. In A Treatise on Zoology. Part II. The Porifera and Coelenterata; Lankaster, E., Ed.; Adam & Charles Black: London, UK, 1900; pp. 1–178. [Google Scholar]
- Erpenbeck, D.; Hall, K.; Alvarez, B.; Büttner, G.; Sacher, K.; Schätzle, S.; Schuster, A.; Vargas, S.; Hooper, J.; Wörheide, G. The phylogeny of halichondrid demosponges: Past and present revisited with DNA-barcoding data. Org. Divers. Evol. 2012, 12, 57–70. [Google Scholar] [CrossRef]
- Wörheide, G.; Dohrmann, M.; Erpenbeck, D.; Larroux, C.; Maldonado, M.; Voigt, O.; Borchiellini, C.; Lavrov, D.V. Deep phylogeny and evolution of sponges (phylum Porifera). Adv. Mar. Biol. 2012, 61, 1–78. [Google Scholar] [PubMed]
- Luo, C.; Reitner, J. First report of fossil ‘keratose’ demosponges in Phanerozoic carbonates: Preservation and 3-D reconstruction. Naturwissenschaften 2014, 101, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Bergquist, P. Order Dictyoceratida Minchin, 1900. In Systema Porifera: A Guide to the Classification of Sponges; Hooper, J., Van Soest, R., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; p. 1021. ISBN 9780306472602. [Google Scholar]
- Cresswell, E. Sponges: Their Nature, History, Modes of Fishing, Varieties, Cultivation, Etc; Sir I. Pitman & Sons: London, UK, 1922. [Google Scholar]
- Dailianis, T.; Tsigenopoulos, C.S.; Dounas, C.; Voultsiadou, E. Genetic diversity of the imperilled bath sponge Spongia officinalis Linnaeus, 1759 across the Mediterranean Sea: Patterns of population differentiation and implications for taxonomy and conservation. Mol. Ecol. 2011, 20, 3757–3772. [Google Scholar] [CrossRef] [PubMed]
- DeLaubenfels, M.; Storr, J. The taxonomy of American commercial sponges. Bull. Mar. Sci. 1958, 8, 99–117. [Google Scholar]
- Storr, J. Ecology of the Gulf of Mexico Commercial Sponges and Its Relation to the Fishery; U.S. Department of the Interior, Bureau of Commercial Fisherie: Waschington, DC, USA, 1964.
- Kim, M.M.; Mendis, E.; Rajapakse, N.; Lee, S.H.; Kim, S.K. Effect of spongin derived from Hymeniacidon sinapium on bone mineralization. J. Biomed. Mater. Res. Part B Appl. Biomater. 2009, 90, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Pronzato, R.; Manconi, R. Mediterranean commercial sponges: Over 5000 years of natural history and cultural heritage. Mar. Ecol. 2008, 29, 146–166. [Google Scholar] [CrossRef]
- Voultsiadou, E. Sponges: An historical survey of their knowledge in Greek antiquity. J. Mar. Biol. Assoc. UK 2007, 87, 1757–1763. [Google Scholar] [CrossRef]
- Pronzato, R.; Cerrano, C.; Cubeddu, T.; Magnino, G.; Manconi, R.; Pastore, S.; Sara, A.; Sidri, M. The millennial history of commercial sponges: From harvesting to farming and integrated aquaculture. Biol. Mar. Mediterr. 2000, 7, 132–143. [Google Scholar]
- Lloyd, J.T. Sponges, its history in medicine with a brief account of its habits and structure. J. Am. Pharm. Assoc. 1928, 17, 149–154. [Google Scholar]
- Wiplinger, G. Der Gebrauch des Xylospongiums. Eine neue Theorie zu den hygienischen Verhältnissen in römischen Latrinen. In Proceedings of the International Frontinus-Symposiumon the Technical and Cultural History of Ancient Baths, Aachen, Germany, 18–22 March 2009; Kreiner, R., Letzner, W., Eds.; Peeters: Leuven, Belgium, 2012; pp. 295–306, ISBN 9789042926615. [Google Scholar]
- Louden, D.; Inderbitzin, S.; Peng, Z.; de Nys, R. Development of a new protocol for testing bath sponge quality. Aquaculture 2007, 271, 275–285. [Google Scholar] [CrossRef]
- Bromiley, G. The International Standard Bible Encyclopedia; Wm. B. Eerdmans Publishing Co.: Grand Rapids, MI, USA, 1979; ISBN 0802881610. [Google Scholar]
- White, C. An account of the successful use of the sponge, in the stoppage of an haemorrhage, occasioned by amputation below the knee; and of the rematkable effects of that application in preventing the absorption of matter. In Cases in Surgery with Remarks: Part 1; W. Johnston: Bath, UK, 1770; pp. 151–158. [Google Scholar]
- Schwarzett Der Gebrauch Des Pressschwammes (Spongia praeparata) in Wundblutungen. In Beobachtungen und Abhaandlungen aus dem Gebiete der Gesammten Praktischen Heilkunde; Band, D. (Ed.) Verlage bei Carl Gerold: Wien, Austria, 1815; pp. 308–311. [Google Scholar]
- Von Raimann, E. Uber den Nutzen des Bade-oder Waschschwammes Bey Heftigen Blutungen. Med. Jahrbücher 1839, 27, 306–307. [Google Scholar]
- Gamgee, S. On pressure in wound treatment. Lancet 1881, 118, 821. [Google Scholar] [CrossRef]
- Gardner, A. The natural and medicinal history of sponge. Med. Surg. Mon. 1866, 1, 1–9. [Google Scholar]
- Zschiesche, P. Die Anwendung des Pressschwammes in der Gynaekologie und Seine Gefahren; Universitaet Greifswald: Greifswald, Germany, 1873. [Google Scholar]
- Haussmann, D. Kann die Erweiterung des Verengten Muttermundes Durch Pressschwamm die Empfangniss Erleichtern? Gesellschaft für Geburtshilfe und Gynäkologie: Berlin, Germany, 1878; pp. 311–327. [Google Scholar]
- Herberger, J. Zur chemischen Kenntnis der gebrannten Kropfschwamme. Repert. Pharm. 1835, 52, 309–327. [Google Scholar]
- Müller, W.E.G.; Batel, R.; Schröder, H.C.; Müller, I.M. Sustainable exploitation of biodiversity (sponges and invertebrates) in the Adriatic Sea in Rovinj (Croatia). Evid. Based Complement. Altern. Med. 2004, 1, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Petrus, C. Naauwkeurige Afbelding en Beschryving van Eene Geheel en al Verloorene, Maar Door Kunst Herstelde Neus en Verhemelte; Seep–Boekverkooper: Amsterdam, The Netherlands, 1771. [Google Scholar]
- Thomson, W. Reporton surgery: On sponge grafting. Dublin J. Med. Sci. 1881, 71, 215–216. [Google Scholar]
- Hamilton, D. On sponge-grafting. Edinb. Med. J. 1881, 27, 385–413. [Google Scholar]
- Ferguson, J. On a modification of sponge-grafting. Br. Med. J. 1882, 16, 1202–1203. [Google Scholar] [CrossRef]
- Sanctuary, T. Three successful cases of sponge-grafting. Br. Med. J. 1883, 16, 1202. [Google Scholar] [CrossRef]
- Case, P. Sponge-grafting. Br. Med. J. 1883, 27, 51–52. [Google Scholar] [CrossRef]
- Kendal, F. On so-called sponge-grafting. J. Anat. Physiol. 1883, 17, 349–362. [Google Scholar]
- Brunett, J. Sponge-grafting. Ann. Surg. 1885, 1, 550–558. [Google Scholar] [CrossRef]
- Acland, T.D. Two cases of sponge grafting. Br. Med. J. 1888, 1, 205–206. [Google Scholar]
- Clark, H.E. Hospital and surgical practice in the hospitals and asylums of Great Britain and Ireland: Compound fracture of skull: Trephining, sponge-grafting. Br. Med. J. 1883, 1, 767–768. [Google Scholar]
- Richter, E. Ueber die Schwammkohle (Spongia tosta), deren Zusammensetzung und der aus ihr bereiteten Tinktur. Apotheker-Zeitung 1911, 33, 317–319. [Google Scholar]
- Smith, J.G. Sponges as a fertilizer. Ind. Eng. Chem. 1913, 5, 850. [Google Scholar] [CrossRef]
- Pronzato, R. Mediterranean sponge fauna: A biologica, historical and cultural heritage. Biogeographia 2003, 24, 91–99. [Google Scholar] [CrossRef]
- Stevely, J.M.; Thompson, J.C.; Warner, R.E. The biology and utilization of Florida’s commercial sponges. Tech. Pap. 1978, 8, 1–49. [Google Scholar]
- Moore, H.F. The commercial sponges and the sponge fisheries. In Proceedings of the Fourth International Fishery Congress, Washington, DC, USA, 22–26 September 1908. [Google Scholar]
- Alcolado, P.M.; Grovas-Hernandez, A.; Marcos, Z. General comments on species inventory, fisheries, culture and some community features of the porifera in Cuba. Boll. Mus. Int. Biol. Univ. Genova 2004, 68, 175–186. [Google Scholar]
- Stevely, J.M.; Sweat, D.E. A preliminary evaluation of the commercial sponge resources of Belize with reference to the location of the Turneffe Island sponge farm. Atoll Res. Bull. 1994, 424, 1–22. [Google Scholar] [CrossRef]
- Stevely, J.; Sweat, D. The Biology and Fishery of Florida’s Commercial Sponges, Sea Grant Marine Agent for Manatee. 2001. Available online: http://edis.ifas.ufl.edu/sg095 (accessed on 15 Juanary 2018).
- Milanese, M.; Sara, A.; Manconi, R.; Ben Abdalla, A.; Pronzato, R. Commercial sponge fishing in Libya: Historical records, present status and perspectives. Fish. Res. 2009, 89, 90–96. [Google Scholar] [CrossRef]
- Crawshay, L.R. Studies in the market sponges I. Growth from the planted cutting. J. Mar. Biol. Assoc. UK 1939, 23, 553–574. [Google Scholar] [CrossRef]
- Moore, F.H. A practical method of sponge culture. In Proceedings of the Fourth International Fishery Congress, Washington, DC, USA, 22–26 September 1908. [Google Scholar]
- Osinga, R.; Tramper, J.; Wijffels, R. Cultivation of marine sponges. Mar. Biotechnol. 1999, 1, 509–532. [Google Scholar] [CrossRef] [PubMed]
- Pronzato, R. Sponge—Fishing, disease and farming in the Mediterranean Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 1999, 9, 485–493. [Google Scholar] [CrossRef]
- Handley, S.; Kelly, S.; Kelly, M. Non-destructive video image analysis method for measuring growth in sponge farming: Preliminary results from the New Zealand bath-sponge Spongia (Heterofibria) manipulatus. N. Z. J. Mar. Freshw. Res. 2003, 37, 613–621. [Google Scholar] [CrossRef]
- Corriero, G.; Longo, C.; Mercurio, M.; Nonnis Marzano, C.; Lembo, G.; Spedicato, M.T. Rearing performance of Spongia officinalis on suspended ropes off the Southern Italian Coast (Central Mediterranean Sea). Aquaculture 2004, 238, 195–205. [Google Scholar] [CrossRef]
- Verdenal, B.; Vecelet, J. Sponge culture on vertical ropes in the northwestern Mediterranean Sea. In New Perspectives in Sponge Biology; Rutzler, K., Ed.; Smithsonian Institution Press: Washington, DC, USA, 1991; ISBN 0874747848. [Google Scholar]
- Pronzato, R.; Bavestrello, G.; Cerrano, C.; Magninio, G.; Manconi, R.; Pantelis, J.; Sara, A.; Sidri, M. Sponge farming in the Mediterranean Sea: New perspectives. Mem. Qld. Museum 1999, 44, 485–491. [Google Scholar]
- Schiefenhovel, K.; Kunzmann, A. Sponge farming trials: Survival, attachment, and growth of two Indo-Pacific sponges, Neopetrosia sp. and Stylissa massa. J. Mar. Biol. 2012, 2012, 417360. [Google Scholar] [CrossRef]
- Duckworth, A.; Wolff, C.; Evans-Illidge, E. Farming bath sponges in tropical Australia. World Aquac. 2009, 40, 20–22. [Google Scholar]
- Kelly, M.; Handley, S.; Page, M.; Butterfield, P.; Hartill, B.; Kelly, S. Aquaculture trials of the New Zealand bath-sponge Spongia (Heterofibria) manipulatus using lanterns. N. Z. J. Mar. Freshw. Res. 2004, 38, 231–241. [Google Scholar] [CrossRef]
- Friday, S. A Study of Sponge Aquaculture in Jambiani: Is Shallow Farming Feasible? Independent Study Project (ISP) Collection: Washington, DC, USA, 2011; p. 1194. [Google Scholar]
- Schatton, W.; Schatton, M.; Müller-Zahn, I.; Müller, W.E.G. Method Using MariBattery (RTM) for Aquatic Cultivation of Sponges, Corals and Other Invertebrates to Improve Water Quality or Provide Raw Materials. Patent No. DE 19834949 A1, 17 February 2000. [Google Scholar]
- Osinga, R. Bioreactor System for the Cultivation of Aquatic Organisms. Patent No. EP 1508272 A1, 23 February 2005. [Google Scholar]
- Amblard, F. Support Pour l’Aquaculture par Bouturage d’Animaux Aquatiques. Patent No. EP 1488696 A1, 22 November 2004. [Google Scholar]
- Van Treeck, P.; Paster, M. Aquakulturzuchteinheit. Patent No. DE 10161174 C1, 12 June 2003. [Google Scholar]
- Duckworth, A. Farming sponges to supply bioactive metabolites and bath sponges: A review. Mar. Biotechnol. 2009, 11, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Garrone, R.; Pottu, J. Collagen biosynthesis in sponges—Elaboration of spongin by spongocytes. J. Submicrosc. Cytol. 1973, 5, 199–218. [Google Scholar]
- Arndt, W. Die Ältesten Bildlichen Darstellungen von. Schwämmen; Ges. naturforschender Freunde: Berlin, Germany, 1931. [Google Scholar]
- Junqua, S.; Robert, L.; Garrone, R.; Pavans de Ceccatty, M.; Vacelet, J. Biochemical and morphological studies on collagens of horny sponges. Ircinia filaments compared to spongines. Connect. Tissue Res. 1974, 2, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Garrone, R. Phylogenesis of connective tissue. In Morphological Aspects and Biosynthesis of Sponge Intercellular Matrix; Karger: Basel, Switzerland, 1978; Volume 5, ISBN 3805527675. [Google Scholar]
- Simpson, L.S. Cell Biology of Sponges; Springer: New York, NY, USA, 1984; ISBN 9781461252146. [Google Scholar]
- Croockewit, J.H. Ueber die Zusammensetzung des Badeschwammes. Justus Liebigs Ann. Chem. 1843, 48, 43–56. [Google Scholar] [CrossRef]
- Hundeshagen, F. Über jodhaltige Spongien and Jodospongin. Angew. Chem. 1895, 8, 473–476. [Google Scholar] [CrossRef]
- Harnack, E. Ueber das Iodospongin, die Jodhaltige eiweissartige Subsanz aus dem Badeschwamm. Z. Physiol. Chem. 1898, 25, 412–424. [Google Scholar] [CrossRef]
- Clancey, V.J. The constitution of sponges. The common bath sponge, Hippospongia equina. Biochem. J. 1926, 20, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Block, R.; Bolling, D. Amino acid composition of keratins. J. Biol. Chem. 1939, 127, 685–693. [Google Scholar]
- Shoulders, M.D.; Raines, R.T. Collagen structure and stability. Annu. Rev. Biochem. 2009, 1998, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Katzman, R.L.; Halford, M.H.; Reinhold, V.N.; Jeanloz, R.W. Invertebrate connective tissue. IX. Isolation and structure determination of glucosylgalactosylhydroxylysine from sponge and sea anemone collagen. Biochemistry 1972, 11, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Gaino, E.; Pronzato, R. Ultrastructural evidence of bacterial damage to Spongia officinalis fibres (Porifera, Demospongiae). Dis. Aquat. Organ. 1989, 6, 67–74. [Google Scholar] [CrossRef]
- Ehrlich, H.; Maldonado, M.; Hanke, T.; Meissner, H.; Born, R. Spongins: Nanostructural investigations and development of biomimetic material model. VDI Ber. 2003, 1803, 287–292. [Google Scholar]
- Allen, W. Tanned Sponge. U.S. Patent 1,336,806 A, 13 April 1920. [Google Scholar]
- Cohn, M. Method of Dyeing Sponges. U.S. Patent 2,056,166 A, 6 November 1934. [Google Scholar]
- Harlow, H.G. Bleaching of Sponges. U.S. Patent 2,344,987 A, 28 March 1944. [Google Scholar]
- Matthews, J.M. Bleaching and Related Processes as Applied to Textile Fibers and Other Materials; The Chemical Catalog Company: New York, NY, USA, 1921; ISBN 134476357X. [Google Scholar]
- Sterrer, W.; Schoepfer-Sterres, C. Marine Fauna and Flora of Bermuda: A Systematic Guide to the Identification of Marine Organisms; Wiley-Interscience: New York, NY, USA, 1986; ISBN 0471823368. [Google Scholar]
- Wahab, M.A.; de Nys, R.; Whalan, S. Closing the lifecycle for the sustainable aquaculture of the bath sponge Coscinoderma matthewsi. Acquaculture 2012, 324, 281–289. [Google Scholar] [CrossRef]
- Von Ledenfeld, R. A monograph of the horny sponges. Ann. Mag. Nat. Hist. 1889, 6, 418–423. [Google Scholar]
- Castritsi-Catharios, J.; Magli, M.; Vacelet, J. Evaluation of the quality of two commercial sponges by tensile strength measurement. J. Mar. Biol. Assoc. 2007, 87, 1765–1771. [Google Scholar] [CrossRef]
- Lim, Y.; Cha, C.M.; Chang, J.Y. Compressible and monolithic microporous polymer sponges prepared via one-pot synthesis. Sci. Rep. 2015, 5, 15957. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.; Simon, P.; Motylenko, M.; Wysokowski, M.; Bazhenov, V.V.; Galli, R.; Stelling, A.L.; Stawski, D.; Ilan, M.; Stöcker, H.; et al. Extreme Biomimetics: Formation of zirconium dioxide nanophase using chitinous scaffolds under hydrothermal conditions. J. Mater. Chem. B 2013, 1, 5092–5099. [Google Scholar] [CrossRef]
- Mutsenko, V.V.; Gryshkov, O.; Lauterboeck, L.; Rogulska, O.; Tarusin, D.N.; Bazhenov, V.V.; Schütz, K.; Brüggemeier, S.; Gossla, E.; Akkineni, A.R.; et al. Novel chitin scaffolds derived from marine sponge Ianthella basta for tissue engineering approaches based on human mesenchymal stromal cells: Biocompatibility and cryopreservation. Int. J. Biol. Macromol. 2017, 104, 1955–1965. [Google Scholar] [CrossRef] [PubMed]
- Mutsenko, V.V.; Bazhenov, V.V.; Rogulska, O.; Tarusin, D.N.; Schütz, K.; Brüggemeier, S.; Gossla, E.; Akkineni, A.R.; Meißner, H.; Djurovi, M.; et al. 3D chitinous scaffolds derived from cultivated marine demosponge Aplysina aerophoba for tissue engineering approaches based on human mesenchymal stromal cells. Int. J. Biol. Macromol. 2017, 104, 1966–1974. [Google Scholar] [CrossRef] [PubMed]
- Green, D.; Howard, D.; Yang, X.; Kelly, M.; Oreffo, R.O.C. Natural marine sponge fiber skeleton: A biomimetic scaffold for human osteoprogenitor cell attachment, growth, and differentiation. Tissue Eng. 2003, 9, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W. Tissue bionics: Examples in biomimetic tissue engineering. Biomed. Mater. 2008, 3, 34010. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W.; Lee, J.; Jung, H. Marine structural biomaterials in medical biomimicry. Tissue Eng. Part B Rev. 2015, 21, 438–450. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Solomon, K.L.; Zhang, X.; Pavlos, N.J.; Abel, T.; Willers, C. In vitro evaluation of natural marine sponge collagen as a scaffold for bone tissue engineering. Int. J. Biol. Sci. 2011, 7, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Nandi, S.K.; Kundu, B.; Mahato, A.; Thakur, N.L.; Joardar, S.N.; Mandal, B.B. In vitro and in vivo evaluation of the marine sponge skeleton as a bone mimicking biomaterial. Integr. Biol. 2015, 7, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Lüer, S.; Troller, R.; Aebi, C. Antibacterial and antiinflammatory kinetics of curcumin as a potential antimucositis agent in cancer patients. Nutr. Cancer 2012, 64, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Chernomorsky, S.; Rancourt, R.; Virdi, K.; Segelman, A.; Poretz, R.D. Antimutagenicity, cytotoxicity and composition of chlorophyllin copper complex. Cancer Lett. 1997, 120, 141–147. [Google Scholar] [CrossRef]
- Rebecca, O.P.S.; Boyce, A.N.; Chandran, S. Pigment identification and antioxidant properties of red dragon fruit (Hylocereus polyrhizus). Afr. J. Biotechnol. 2010, 9, 1450–1454. [Google Scholar]
- Bonarska-Kujawa, D.; Cyboran, S.; Żyłka, R.; Oszmiański, J.; Kleszczyńska, H. Biological activity of blackcurrant extracts (Ribes nigrum L.) in relation to erythrocyte membranes. Biomed. Res. Int. 2014, 2014, 783059. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, R.N.; Santos, D.T.; Meireles, M.A. Non-thermal stabilization mechanisms of anthocyanins in model and food systems—An overview. Food Res. Int. 2011, 44, 499–509. [Google Scholar] [CrossRef]
- Shahid, M.; Mohammad, F. Recent advancements in natural dye applications: A review. J. Clean. Prod. 2013, 53, 310–331. [Google Scholar] [CrossRef]
- Hug, H.; Bader, M.; Mair, P.; Glatzel, T. Biophotovoltaics: Natural pigments in dye-sensitized solar cells. Appl. Energy 2014, 115, 216–225. [Google Scholar] [CrossRef]
- Norman, M.; Bartczak, P.; Zdarta, J.; Tylus, W.; Szatkowski, T.; Stelling, A.; Ehrlich, H.; Jesionowski, T. Adsorption of C.I. Natural Red 4 onto spongin skeleton of marine demosponge. Materials 2014, 8, 96–116. [Google Scholar] [CrossRef] [PubMed]
- Norman, M.; Bartczak, P.; Zdarta, J.; Ehrlich, H.; Jesionowski, T. Anthocyanin dye conjugated with Hippospongia communis marine demosponge skeleton and its antiradical activity. Dyes Pigments 2016, 134, 541–552. [Google Scholar] [CrossRef]
- Norman, M.; Bartczak, P.; Zdarta, J.; Tomala, W.; Żurańska, B.; Dobrowolska, A.; Piasecki, A.; Czaczyk, K.; Ehrlich, H.; Jesionowski, T. Sodium copper chlorophyllin immobilization onto Hippospongia communis marine demosponge skeleton and its antibacterial activity. Int. J. Mol. Sci. 2016, 17, 1564. [Google Scholar] [CrossRef] [PubMed]
- Arimoto-Kobayashi, S.; Harada, N.; Tokunaga, R.; Odo, J.; Hayatsu, H. Adsorption of mutagens to chlorophyllin—Chitosan, an insoluble form of chlorophyllin. Mutat. Res. 1997, 381, 243–249. [Google Scholar] [CrossRef]
- Hayatsu, H.; Sugiyama, C.; Arimoto-Kobayashi, S.; Negishi, T. Porphyrins as possible preventers of heterocyclic amine carcinogenesis. Cancer Lett. 1999, 143, 185–187. [Google Scholar] [CrossRef]
- Rocha Oliveira, G.; Dias Do Amaral, L.J.; Giovanela, M.; Da Silva Crespo, J.; Fetter, G.; Rivera, J.A.; Sampieri, A.; Bosch, P. Bactericidal performance of chlorophyllin-copper hydrotalcite compounds. Water Air Soil Pollut. 2015, 226, 226–316. [Google Scholar] [CrossRef]
- Azimi, S.; Behin, J.; Abiri, R.; Rajabi, L.; Derakhshan, A.; Karimnezhad, H. Synthesis, characterization and antibacterial activity of chlorophyllin functionalized graphene oxide nanostructures. Sci. Adv. Mater. 2014, 6, 771–781. [Google Scholar] [CrossRef]
- Wiegand, C.; Springer, S.; Abel, M.; Wesarg, F.; Ruth, P.; Hipler, U.C. Application of a drainage film reduces fibroblast ingrowth into large-pored polyurethane foam during negative-pressure wound therapy in an in vitro model. Wound Repair Regen. 2013, 21, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Malmsjo, M.; Ingermansson, R.; Martin, R.; Huddleston, E. Negative-pressure wound therapy using gauze or open-cell polyurethane foam: Similar early effects on pressure transduction and tissue contraction in an experimental porcine wound model. Wound Repair Regen. 2009, 17, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, U.; Turan, A.; Makroc, F.; Erkorkmaz, U.; Elmas, C.; Kostakoglu, N. Loofah sponge as an interface dressong material in negative pressure wound theraphy: Results of an in vitro study. Ostomy Wound Manag. 2014, 60, 37–45. [Google Scholar]
- Liu, J.; Morykwas, M.J.; Argenta, L.C.; Wagner, W.D. Development of a biodegradable foam for use in negative pressure wound therapy. J. Biomed. Mater. Res. B Appl. Biomater. 2011, 98, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Assadian, O.; Assadian, A.; Stadler, M.; Diab-Elschahawi, M.; Kramer, A. Bacterial growth kinetic without the influence of the immune system using vacuum-assisted closure dressing with and without negative pressure in an in vitro wound model. Int. Wound J. 2010, 7, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Ogura, S.; Tabata, K.; Fukushima, K.; Kamachi, T. Development of phthalocyanines for photodynamic therapy. J. Porphyr. Phthalocyanines 2006, 10, 1116–1124. [Google Scholar] [CrossRef]
- Zagal, J.H.; Griveau, S.; Silva, J.F.; Nyokong, T.; Bedioui, F. Metallophthalocyanine-based molecular materials as catalysts for electrochemical reactions. Coord. Chem. Rev. 2010, 254, 2755–2791. [Google Scholar] [CrossRef]
- Lian, K.; Li, R.; Wang, H.; Zhang, J.; Gamota, D. Printed flexible memory devices using copper phthalocyanine. Mater. Sci. Eng. B-Adv. Funct. Solid-State Mater. 2010, 167, 12–16. [Google Scholar] [CrossRef]
- Wen, Z.; Kang, T. Determination of nitrite using sensors based on nickel phthalocyanine polymer modified electrodes. Talanta 2004, 62, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.H.; Wikle, H.C.; Chin, B.A. Passive chemiresistor sensor based on iron(II) phthalocyanine thin films for monitoring of nitrogen dioxide. Sens. Actuators B Chem. 2010, 148, 498–503. [Google Scholar] [CrossRef]
- Amini, M.K.; Shahrokhian, S.; Tangestaninejad, S.; Mirkhani, V. Iron(II) phthalocyanine-modified carbon-paste electrode for potentiometric detection of ascorbic acid. Anal. Biochem. 2001, 290, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Wohrle, D.; Schnurpfeil, G.; Makarov, S.G.; Kazarin, A.; Suvorova, O.N. Practical applications of phthalocyanines—From dyes and pigments to materials for optical, electronic and photo-electronic devices. Macroheterocycles 2012, 5, 191–202. [Google Scholar] [CrossRef]
- Jin, L.; Ding, Z.; Chen, D. Zinc octacarboxylic phthalocyanine/lutein dyads co-adsorbed nanocrystalline TiO2 electrode: Enhancement in photovoltaic performance of dye-sensitized solar cells. J. Mater. Sci. 2013, 48, 4883–4891. [Google Scholar] [CrossRef]
- Stylianakis, M.M.; Konios, D.; Viskadouros, G.; Vernardou, D.; Katsarakis, N.; Koudoumas, E.; Anastasiadis, S.H.; Kymakis, E. Ternary organic solar cells incorporating zinc phthalocyanine with improved performance exceeding 8.5%. Dyes Pigments 2017, 146, 408–413. [Google Scholar] [CrossRef]
- Raja, R.; Ratnasamy, P. Oxidation of cyclohexane over copper phthalocyanines encapsulated in zeolites. Catal. Lett. 1997, 48, 1–10. [Google Scholar] [CrossRef]
- DeOliveira, E.; Neri, C.R.; Ribeiro, A.O.; Garcia, V.S.; Costa, L.L.; Moura, A.O.; Prado, A.G.S.; Serra, O.; Iamamoto, Y. Hexagonal mesoporous silica modified with copper phthalocyanine as a photocatalyst for pesticide 2,4-dichlorophenoxiacetic acid degradation. J. Colloid Interface Sci. 2008, 323, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Vargas, E.; Vargas, R.; Núñez, O. A TiO2 surface modified with copper(II) phthalocyanine-tetrasulfonic acid tetrasodium salt as a catalyst during photoinduced dichlorvos mineralization by visible solar light. Appl. Catal. B Environ. 2014, 156–157, 8–14. [Google Scholar] [CrossRef]
- Mirzaeian, M.; Rashidi, A.M.; Zare, M.; Ghabezi, R.; Lotfi, R. Mercaptan removal from natural gas using carbon nanotube supported cobalt phthalocyanine nanocatalyst. J. Nat. Gas Sci. Eng. 2014, 18, 439–445. [Google Scholar] [CrossRef]
- Chen, S.; Huang, X.; Xu, Z. Decoration of phthalocyanine on multiwalled carbon nanotubes/cellulose nanofibers nanocomposite for decoloration of dye wastewater. Compos. Sci. Technol. 2014, 101, 11–16. [Google Scholar] [CrossRef]
- Vashurin, A.S.; Badaukaite, R.A.; Futerman, N.A.; Pukhovskaya, S.G.; Shaposhnikov, G.P.; Golubchikov, O.A. Catalytic properties of polymer matrix-immobilized cobalt complexes with sulfonated phthalocyanines. Pet. Chem. 2013, 53, 197–200. [Google Scholar] [CrossRef]
- Sevim, A.M.; Ilgün, C.; Gül, A. Preparation of heterogeneous phthalocyanine catalysts by cotton fabric dyeing. Dyes Pigments 2011, 89, 162–168. [Google Scholar] [CrossRef]
- Norman, M.; Zdarta, J.; Bartczak, P.; Piasecki, A.; Petrenko, I.; Ehrlich, H.; Jesionowski, T. Marine sponge skeleton photosensitized by copper phthalocyanine: A catalyst for Rhodamine B degradation. Open Chem. 2016, 14, 243–254. [Google Scholar] [CrossRef]
- Norman, M.; Żółtowska-Aksamitowska, S.; Zgoła-Grześkowiak, A.; Ehlich, H.; Jesionowski, T. Iron(III) phthalocyanine supported on a spongin scaffold as an advanced photocatalyst in a highly efficient removal process of halophenols and bisphenol A. J. Hazard. Mater. 2018, 347, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H. Biological Materials of Marine Origin: Invertebrates; Springer: Dordrecht, The Netherlands, 2010; ISBN 9789048196401. [Google Scholar]
- Ehrlich, H. (Ed.) Extreme Biomimetics; Springer Nature: Cham, Switzerland, 2017; ISBN 99783319453385. [Google Scholar]
- Wysokowki, M.; Motylenko, M.; Stocker, H.; Bazhenov, V.V.; Langer, E.; Dobrowolska, A.; Czaczyk, K.; Galli, R.; Stelling, A.L.; Behm, T.; et al. An extreme biomimetic approach: Hydrothermal synthesis of β-chitin/ZnO nanostructured composites. J. Mater. Chem. B 2013, 1, 6469–6476. [Google Scholar] [CrossRef]
- Wysokowski, M.; Motylenko, M.; Beyer, J.; Makarova, A.; Stöcker, H.; Walter, J.; Galli, R.; Kaiser, S.; Vyalikh, D.; Bazhenov, V.V.; et al. Extreme biomimetic approach for development of novel chitin-GeO2 nanocomposites with photoluminescent properties. Nano Res. 2015, 8, 2288–2301. [Google Scholar] [CrossRef]
- Wysokowski, M.; Motylenko, M.; Walter, J.; Lota, G.; Wojciechowski, J.; Stöcker, H.; Galli, R.; Stelling, A.L.; Himcinschi, C.; Niederschlag, E.; et al. Synthesis of nanostructured chitin—Hematite composites under extreme biomimetic conditions. RSC Adv. 2014, 4, 61743–61752. [Google Scholar] [CrossRef]
- Wysokowski, M.; Petrenko, I.; Motylenko, M.; Langer, E.; Bazhenov, V.V.; Galli, R.; Stelling, A.L.; Kljajić, Z.; Szatkowski, T.; Kutsova, W.; et al. Renewable chitin from marine sponge as a thermostable biological template for hydrothermal synthesis of hematite nanospheres using principles of extreme biomimetics. Bioinspired Mater. 2015, 1, 12–22. [Google Scholar] [CrossRef]
- Wysokowski, M.; Motylenko, M.; Rafaja, D.; Koltsov, I.; Stöcker, H.; Szalaty, T.; Bazhenov, V.V.; Stelling, A.L.; Beyer, J.; Heitmann, J.; et al. Extreme biomimetic approach for synthesis of nanocrystalline chitin-(Ti, Zr)O2 multiphase composites. Mater. Chem. Phys. 2017, 188, 115–124. [Google Scholar] [CrossRef]
- Wysokowski, M.; Behm, T.; Born, R.; Bazhenov, V.V.; Meißner, H.; Richter, G.; Szwarc-Rzepka, K.; Makarova, A.; Vyalikh, D.; Schupp, P.; et al. Preparation of chitin—Silica composites by in vitro silicification of two-dimensional Ianthella basta demosponge chitinous scaffolds under modified Stöber conditions. Mater. Sci. Eng. C Mater. Biol. Appl. 2013, 33, 3935–3941. [Google Scholar] [CrossRef] [PubMed]
- Bazhenov, V.V.; Wysokowski, M.; Petrenko, I.; Stawski, D.; Sapozhnikov, P.; Born, R.; Stelling, A.L.; Kaiser, S.; Jesionowski, T. Preparation of monolithic silica-chitin composite under extreme biomimetic conditions. Int. J. Biol. Macromol. 2015, 76, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Wysokowski, M.; Materna, K.; Walter, J.; Petrenko, I.; Stelling, A.L.; Bazhenov, V.V.; Klapiszewski, Ł.; Szatkowski, T.; Lewandowska, O.; Stawski, D.; et al. Solvothermal synthesis of hydrophobic chitin—Polyhedral oligomeric silsesquioxane (POSS) nanocomposites. Int. J. Biol. Macromol. 2015, 78, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Gholipourmaleabadi, M.; Mozafari, M. Fabrication of newly developed pectin—GeO2 nanocomposite using extreme biomimetics route and its antibacterial activities. J. Macromol. Sci. Part A 2017, 51, 655–661. [Google Scholar] [CrossRef]
- Szatkowski, T.; Wysokowski, M.; Lota, G.; Pęziak, D.; Bazhenov, V.V.; Nowaczyk, G.; Walter, J.; Molodtsov, S.L.; Stöcker, H.; Himcinschi, C.; et al. Novel nanostructured hematite–spongin composite developed using an extreme biomimetic approach. RSC Adv. 2015, 5, 79031–79040. [Google Scholar] [CrossRef]
- Szatkowski, T.; Siwińska-Stefańska, K.; Wysokowski, M.; Stelling, A.; Joseph, Y.; Ehrlich, H.; Jesionowski, T. Immobilization of titanium(IV) oxide onto 3D spongin scaffolds of marine sponge origin according to extreme biomimetics principles for removal of C.I. Basic Blue 9. Biomimetics 2017, 2, 4. [Google Scholar] [CrossRef]
- Szatkowski, T.; Jesionowski, T. Hydrothermal synthesis of spongin-based materials. In Extreme Biomimetic; Ehrlich, H., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 251–271. ISBN 9783319453385. [Google Scholar]
- Szatkowski, T.; Kopczyński, K.; Motylenko, M.; Bormann, H.; Mania, B.; Graś, M.; Lota, G.; Bazhenov, V.V.; Rafaja, D.; Roth, F.; et al. Extreme Biomimetics: Carbonized 3D spongin scaffold as a novel support for nanostructured manganese oxide(IV) and its electrochemical applications. Nano Res. 2018. just accepted. [Google Scholar] [CrossRef]













© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesionowski, T.; Norman, M.; Żółtowska-Aksamitowska, S.; Petrenko, I.; Joseph, Y.; Ehrlich, H. Marine Spongin: Naturally Prefabricated 3D Scaffold-Based Biomaterial. Mar. Drugs 2018, 16, 88. https://doi.org/10.3390/md16030088
Jesionowski T, Norman M, Żółtowska-Aksamitowska S, Petrenko I, Joseph Y, Ehrlich H. Marine Spongin: Naturally Prefabricated 3D Scaffold-Based Biomaterial. Marine Drugs. 2018; 16(3):88. https://doi.org/10.3390/md16030088
Chicago/Turabian StyleJesionowski, Teofil, Małgorzata Norman, Sonia Żółtowska-Aksamitowska, Iaroslav Petrenko, Yvonne Joseph, and Hermann Ehrlich. 2018. "Marine Spongin: Naturally Prefabricated 3D Scaffold-Based Biomaterial" Marine Drugs 16, no. 3: 88. https://doi.org/10.3390/md16030088
APA StyleJesionowski, T., Norman, M., Żółtowska-Aksamitowska, S., Petrenko, I., Joseph, Y., & Ehrlich, H. (2018). Marine Spongin: Naturally Prefabricated 3D Scaffold-Based Biomaterial. Marine Drugs, 16(3), 88. https://doi.org/10.3390/md16030088