Astaxanthin Restrains Nitrative-Oxidative Peroxidation in Mitochondrial-Mimetic Liposomes: A Pre-Apoptosis Model

 ,
,
Abstract
:
1. Introduction
2. Results
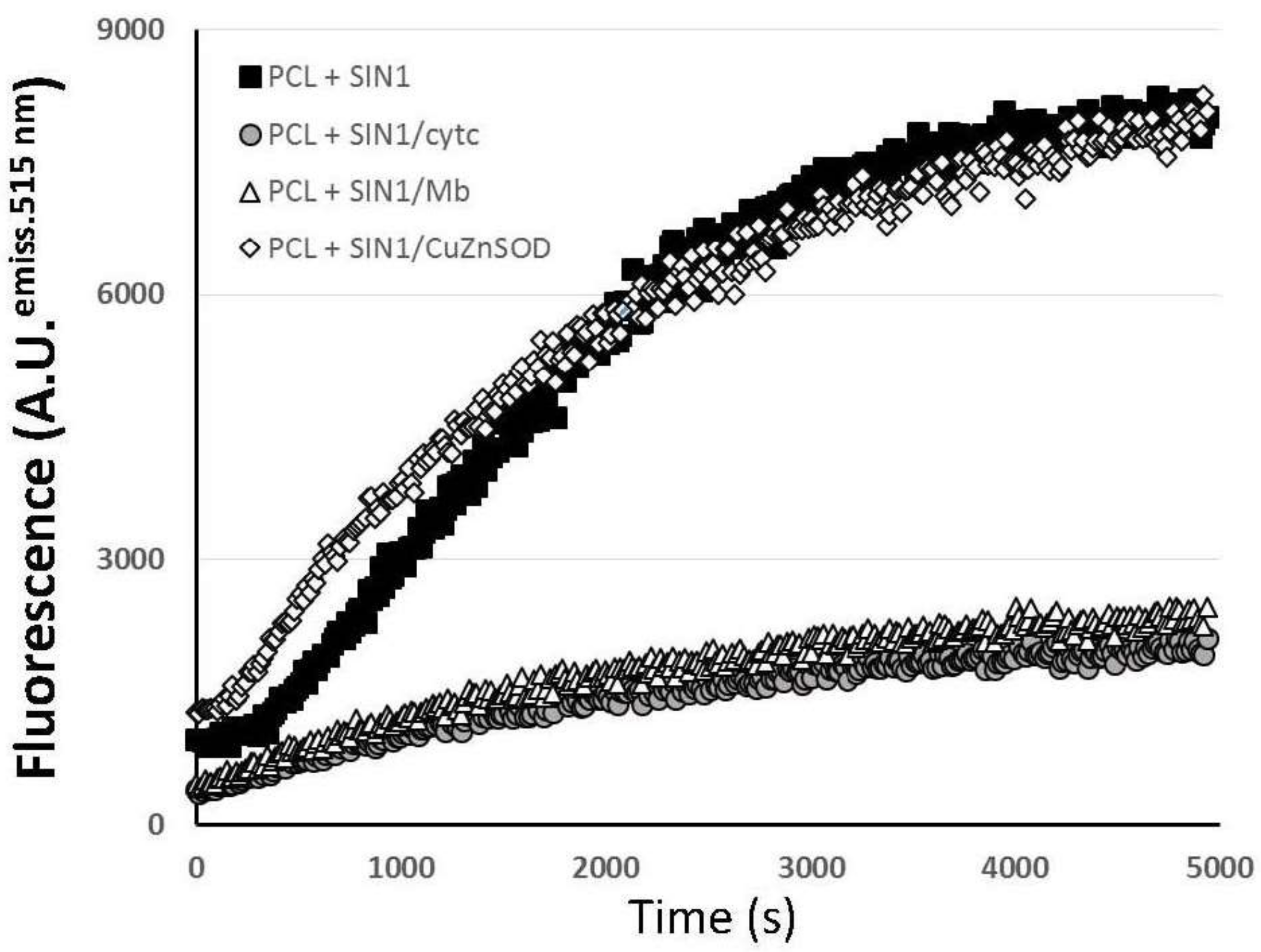
2.1. Oxidative/Nitrative Modifications on Cytochrome C
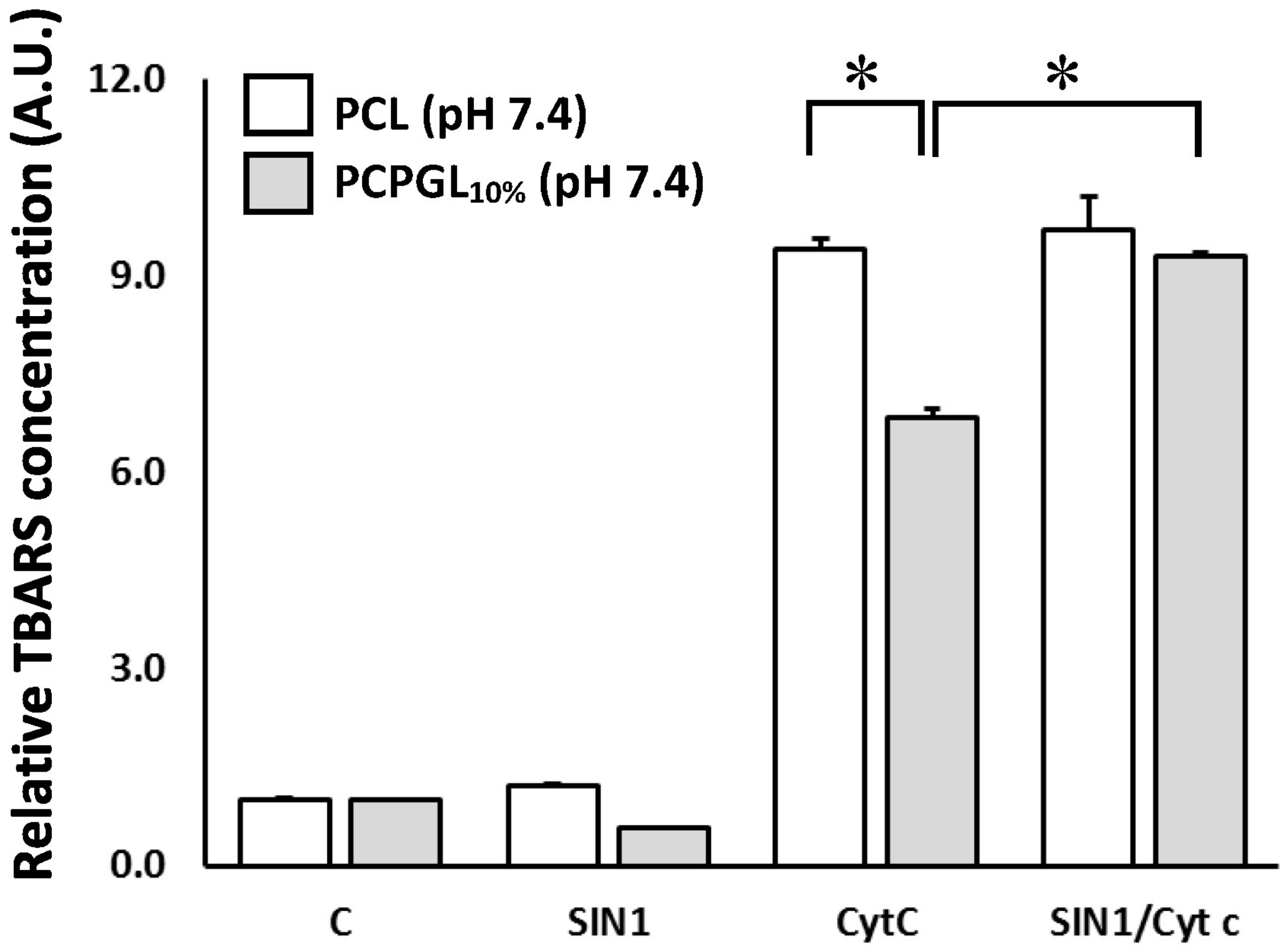
2.2. Peroxidation of Liposomes by the SIN-1/Cyt C System
2.3. Astaxanthin Effect on Liposome Size
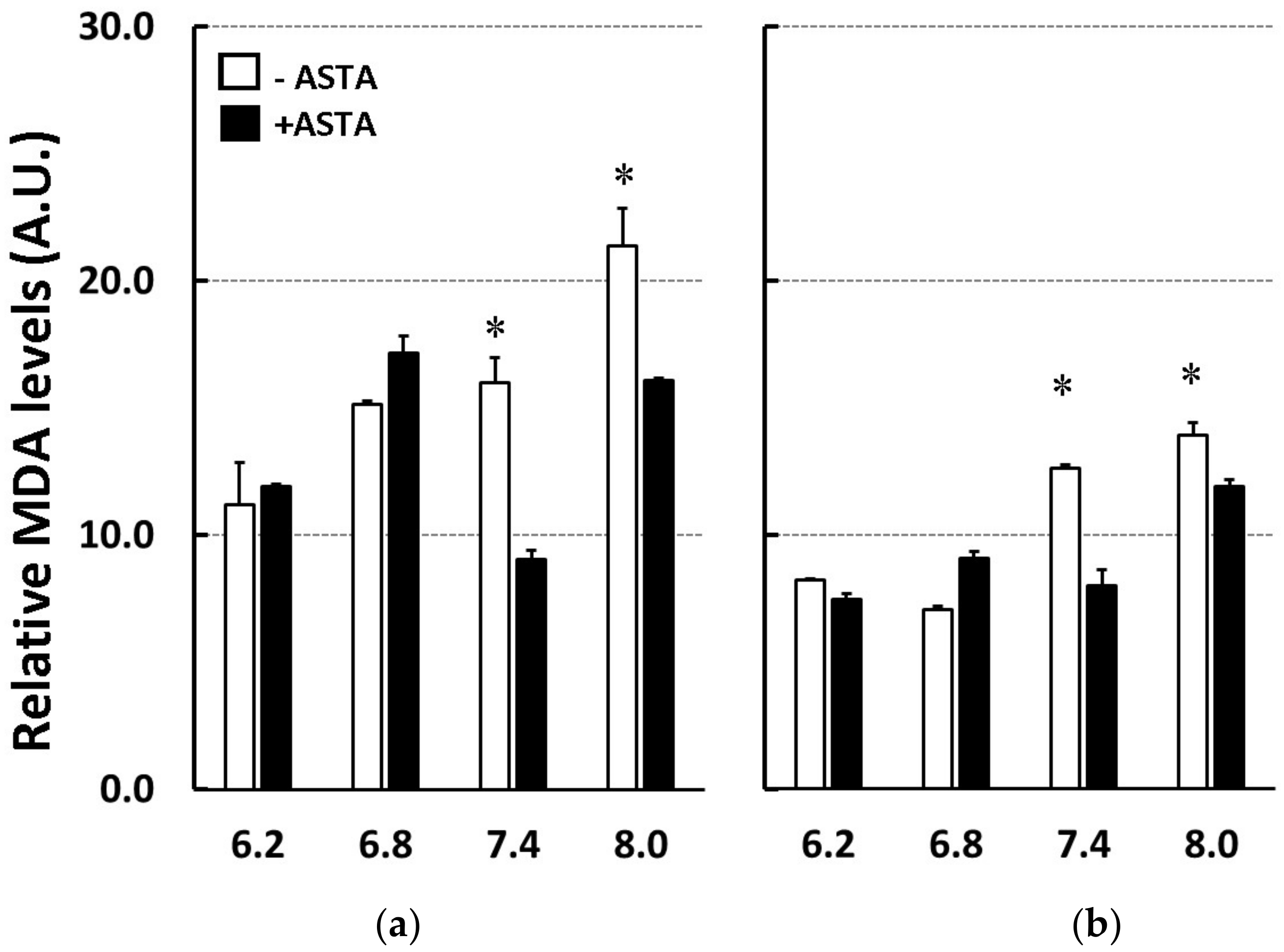
2.4. pH Effect on Lipid Peroxidation and Astaxanthin Antioxidant Properties
3. Discussion
3.1. Cytochrome c-Modified by SIN-1 Induces Liposome Peroxidation
3.2. Liposome Size/Organization is Unaltered by Astaxanthin Incorporation
3.3. Astaxanthin Suppresses Cyt C/SIN-1-Induced Liposome Oxidation at pH > 7.4
3.4. Mechanism of Astaxanthin-Protective Role against Liposome Peroxidation
4. Materials and Methods
4.1. Chemicals
4.2. Astaxanthin Stock Solutions
4.3. Preparation of Unilamellar Liposomes
- 5 mM phosphatidylcholine (5 mM PCL);
- 4.76 mM phosphatidylcholine + 0.24 mM phpsphatidylglycerol (PCPGL 5%); and
- 4.54 mM phosphatidylcholine + 0.46 mM phpsphatidylglycerol (PCPGL 10%).
4.4. Dynamic Light Scattering Studies
4.5. Oxidative/Nitrative Conditions for Liposome Peroxidation
- 5 mM PCL/PCPGL + 50 mM tPi (pH 6.2–8);
- 5 mM PCL/PCPGL + 50 mM tPi (pH 6.2–8) + 150 µM SIN-1;
- 5 mM PCL/PCPGL + 50 mM tPi (pH 6.2–8) + 150 µM SIN-1 + 150 µM cyt c;
- 5 mM PCL/PCPGL + 50 mM tPi (pH 6.2–8) + 150 µM KO2;
- 5 mM PCL/PCPGL + 50 mM tPi (pH 6.2–8) + 150 µM SIN-1 + 0.6 U/mL SOD;
- 5 mM PCL/PCPGL + 50 mM tPi (pH 6.2–8) + 150 µM KO2 + 0.6 U/mL SOD;
4.6. UV-Vis Spectrophotometry
4.7. Nitric Oxide Concentration
4.8. Indices of Lipid Peroxidation
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Polotow, T.G.; Poppe, S.C.; Vardaris, C.V.; Ganini, D.; Guariroba, M.; Mattei, R.; Mattei, R.; Hatanaka, E.; Martins, M.F.; Bondan, E.F.; et al. Redox status and neuro inflammation indexes in cerebellum and motor cortex of wistar rats supplemented with natural sources of omega-3 fatty acids and astaxanthin: Fish Oil, Krill Oil, and Algal Biomass. Mar. Drugs 2015, 13, 6117–6137. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Kuroki, T.; Ikeda, S.; Okada, T.; Maoka, T.; Kitamura, A.; Sugimoto, M.; Kume, S. Astaxanthin ameliorates heat stress-induced impairment of blastocyst development in vitro: Astaxanthin colocalization with and action on mitochondria. J. Assist. Reprod. Genet. 2013, 30, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Bolin, A.P.; Macedo, R.C.; Marin, D.P.; Barros, M.P.; Otton, R. Astaxanthin prevents in vitro auto-oxidative injury in human lymphocytes. Cell Biol. Toxicol. 2010, 26, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, K.; Kowshik, J.; Kishore, T.K.; Baba, A.B.; Nagini, S. Astaxanthin inhibits NF-κB and Wnt/β-catenin signaling pathways via inactivation of Erk/MAPK and PI3K/Akt to induce intrinsic apoptosis in a hamster model of oral cancer. Biochim. Biophys. Acta 2013, 1830, 4433–4444. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, W.; Lee, J.H.; Jeon, S.J.; Choi, Y.H.; Kim, B.W.; Chang, H.I.; Nam, S.W. Astaxanthin inhibits H2O2-mediated apoptotic cell death in mouse neural progenitor cells via modulation of P38 and MEK signaling pathways. J. Microbiol. Biotechnol. 2009, 19, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome c-mediated apoptosis. Ann. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Byungki, J.; Sanghwa, H. Biochemical properties of cytochrome c nitrated by peroxynitrite. Biochimie 2006, 88, 53–58. [Google Scholar] [CrossRef]
- Kawai, C.; Ferreira, J.C.; Baptista, M.S.; Nantes, I.L. Not only oxidation of cardiolipin affects the affinity of cytochrome c for lipid bilayers. J. Phys. Chem. 2014, 118, 11863–11872. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, M.A.; Cooper, C.E. Reaction of nitric oxide with mitochondrial cytochrome c: A novel mechanism for the formation of nitroxyl anion and peroxide. Biochem. J. 1998, 332, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Schonhoff, M.C.; Gaston, B.; Mannick, J.B. Nitrosylation of cytochrome c during apoptosis. J. Biol. Chem. 2003, 278, 18265–18270. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.M.; Asoh, S.; Hiranuma, H.; Ohsawa, I.; Iio, K.; Satou, A.; Ishikura, M.; Ohta, S. Astaxanthin protects mitochondrial redox state and functional integrity against oxidative stress. J. Nutr. Biochem. 2010, 21, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Mano, C.M.; Barros, M.P.; Faria, P.A.; Prieto, T.; Dyszy, F.H.; Nascimento, O.R.; Nantes, I.L.; Bechara, E.J. Superoxide radical protects liposome-contained cytochrome c against oxidative damage promoted by peroxynitrite and free radicals. Free Radic. Biol. Med. 2009, 47, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, B.S.; Edzuma, A.N.; Hough, M.A.; Blundell, K.L.I.M.; Kagan, V.E.; Kapralov, A.A.; Fraser, L.A.; Butt, J.N.; Silkstone, G.G.; Wilson, M.T.; et al. The hydrogen-peroxide-induced radical behavior in human cytochrome c–phospholipid complexes: Implications for the enhanced pro-apoptotic activity of the G41S mutant. Biochem. J. 2013, 456, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Cassina, A.M.; Hodara, R.; Souza, J.M.; Thomson, L.; Castro, L.; Ischiropoulos, H.; Freeman, B.A.; Radi, R. Cytochrome c nitration by peroxynitrite. J. Biol. Chem. 2000, 275, 21409–21415. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Guha, P.; Bhattarai, R.; Nahak, P.; Karmakar, G.; Chettri, P.; Panda, A.K. Influence of lipid composition, pH, and temperature on physicochemical properties of liposomes with curcumin as model drug. J. Oleo Sci. 2016, 65, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Heberle, F.A.; Tristram-Nagle, S.; Szymanski, M.; Koepfinger, M.; Katsaras, J.; Nučerka, N. Molecular structures of fluid phase phosphatidylglycerol bilayers as determined by small angle neutron and X-ray scattering. Biochim. Biophys. Acta 2012, 1818, 2135–2148. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Tan, C.; Zhang, Y.; Abbas, S.; Feng, B.; Zhang, X.; Qin, F. Modulating effect of lipid bilayer–carotenoid interactions on the property of liposome encapsulation. Colloids Surf. B Biointerfaces 2015, 128, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Porcelli, A.M.; Ghelli, A.; Zanna, C.; Pinton, P.; Rizzuto, R.; Rugolo, M. pH difference across the outer mitochondrial membrane measured with a green fluorescent protein mutant. Biochem. Biophys. Res. Commun. 2005, 326, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Llopi, J.; Deveraux, Q.; Tsien, R.; Reed, J. Changes in intramitochondrial and cytosolic pH: Early events that modulate caspase activation during apoptosis. Nat. Cell Biol. 2000, 2, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Matsuyama, S.; Reed, J.C. Mitochondria-dependent apoptosis and cellular pH regulation. Cell Death Differ. 2000, 7, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Moncelli, M.R.; Becucci, L.; Guidelli, R. The intrinsic pKa values for phosphatidylcholine, phosphatidylethanolamine, and phosphatidylserine in monolayers deposited on mercury electrodes. Biophys. J. 1994, 66, 1969–1980. [Google Scholar] [CrossRef]
- Fiuza, B.; Subelzú, N.; Calcerrada, P.; Straliotto, M.R.; Piacenza, L.; Cassina, A.; Rocha, J.B.; Radi, R.; de Bem, A.F.; Peluffo, G. Impact of SIN-1-derived peroxynitrite flux on endothelial cell redox homeostasis and bioenergetics: Protective role of diphenyl diselenide via induction of peroxiredoxins. Free Radic. Res. 2015, 49, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Schafer, F.Q.; Buettner, G.R. Acidic pH amplifies iron-mediated lipid peroxidation in cells. Free Radic. Biol. Med. 2000, 28, 1175–1181. [Google Scholar] [CrossRef]
- Goto, S.; Kogure, K.; Abe, K.; Kimata, Y.; Kitahama, K.; Yamashita, E.; Terada, H. Efficient radical trapping at the surface and inside the phospholipid membrane is responsible for highly potent antiperoxidative activity of the carotenoid astaxanthin. Biochim. Biophys. Acta 2001, 1512, 251–258. [Google Scholar] [CrossRef]
- El-Agamey, A.; Edge, R.; Navaratnam, S.; Land, E.J.; Truscott, T.G. Carotenoid radical anions and their protonated derivatives. Org. Lett. 2006, 8, 4255–4258. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Osawa, T. Cis astaxanthin and especially 9-cis astaxanthin exhibits a higher antioxidant activity in vitro compared to the all-trans isomer. Biochem. Biophys. Res. Commun. 2002, 357, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, A.; Skibsted, L.H. Kinetics and mechanism of the primary steps of degradation of carotenoids by acid in homogeneous solution. J. Agric. Food Chem. 2000, 48, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Begum, S.; Cianci, M.; Durbeej, B.; Falklöf, O.; Hädener, A.; Helliwell, J.R.; Helliwell, M.; Regan, A.C.; Watt, C.l.F. On the origin and variation of colors in lobster carapace. Phys. Chem. Chem. Phys. 2015, 17, 16723–16732. [Google Scholar] [CrossRef] [PubMed]
- Nantes, I.L.; Faljoni-Alário, A.; Nascimento, O.R.; Bandy, B.; Gatti, R.; Bechara, E.J.H. Modifications in heme iron of free and vesicle bound cytochrome c by tert-butyl hydroperoxide: A magnetic circular dichroism and electron paramagnetic resonance investigation. Free Radic. Biol. Med. 2000, 28, 786–796. [Google Scholar] [CrossRef]
- Fraga, C.G.; Leibovitz, B.E.; Tappel, A.L. Lipid peroxidation measured as thiobarbituric acid-reactive substances in tissue slices: Characterization and comparison with homogenates and microsomes. Free Radic. Biol. Med. 1988, 4, 155–161. [Google Scholar] [CrossRef]
- Domijan, A.M.; Ralić, J.; Radić-Brkanac, S.; Rumora, L.; Žanić-Grubišić, T. Quantification of malondialdehyde by HPLC-FL—Application to various biological samples. Biomed. Chromatogr. BMC 2015, 29, 41–46. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liposome | Control (Diameter, nm) | +25 µM ASTA (Diameter, nm) |
|---|---|---|
| PCL | 196.7 ± 0.8 | 184.2 ± 0.6 |
| 5% PCPGL | 191.5 ± 0.3 | 181.2 ± 0.3 |
| 10% PCPGL | 189.1 ± 1.3 | 190.4 ± 0.4 |
| TBARS Levels | pH | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 6.2 | 6.8 | 7.4 | 8.0 | ||||||
| −ASTA | +ASTA | −ASTA | +ASTA | −ASTA | +ASTA | −ASTA | +ASTA | ||
| PCL | C | 1.00 ± 0.04 | 1.00 ± 0.04 | 1.00 ± 0.13 | 1.00 ± 0.01 | 1.00 ± 0.08 | 1.00 ± 0.02 | 1.00 ± 0.01 | 1.00 ± 0.05 |
| SIN1 | 0.77 ± 0.01 | 1.19 ± 0.01 | 0.88 ± 0.02 | 0.95 ± 0.01 | 0.82 ± 0.04 | 1.10 ± 0.02 | 1.14 ± 0.03 | 1.02 ± 0.03 | |
| KO2 | 0.79 ± 0.05 | 1.03 ± 0.03 | 0.77 ± 0.22 | 0.92 ± 0.04 | 1.06 ± 0.07 | 1.04 ± 0.01 | 0.98 ± 0.05 | 0.97 ± 0.02 | |
| SIN1+SOD | 0.92 ± 0.01 | 1.18 ± 0.03 | 0.90 ± 0.03 | 0.95 ± 0.07 | 1.09 ± 0.05 | 1.15 ± 0.02 | 1.26 ± 0.03 | 1.06 ± 0.03 | |
| KO2+SOD | 0.71 ± 0.01 | 1.07 ± 0.01 | 1.00 ± 0.03 | 0.95 ± 0.06 | 1.18 ± 0.10 | 1.06 ± 0.01 | 0.84 ± 0.04 | 1.12 ± 0.05 | |
| PCPGL 10% | C | 1.00 ± 0.61 | 1.00 ± 0.19 | 1.00 ± 0.39 | 1.00 ± 0.46 | 1.00 ± 0.03 | 1.00 ± 0.01 | 1.00 ± 0.03 | 1.00 ± 0.03 |
| SIN1 | 1.56 ± 0.79 | 1.21 ± 0.20 | 1.24 ± 0.47 | 1.35 ± 1.00 | 1.38 ± 0.01 | 0.88 ± 0.01 | 1.14 ± 0.01 | 0.90 ± 0.04 | |
| KO2 | 2.25 ± 0.49 | 1.32 ± 0.28 | 1.16 ± 0.26 | 1.93 ± 0.49 | 1.10 ± 0.05 | 0.95 ± 0.02 | 0.95 ± 0.04 | 0.89 ± 0.02 | |
| SIN1+SOD | 1.91 ± 0.60 | 1.11 ± 0.26 | 1.09 ± 0.65 | 0.86 ± 0.37 | 1.53 ± 0.01 | 0.90 ± 0.03 | 1.13 ± 0.01 | 0.92 ± 0.01 | |
| KO2+SOD | 1.48 ± 0.40 | 2.21 ± 0.25 | 1.49 ± 0.47 | 1.30 ± 0.35 | 1.29 ± 0.02 | 0.97 ± 0.02 | 1.07 ± 0.03 | 0.92 ± 0.02 | |
| MDA-HPLC Levels | pH | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 6.2 | 6.8 | 7.4 | 8.0 | ||||||
| −ASTA | +ASTA | −ASTA | +ASTA | −ASTA | +ASTA | −ASTA | +ASTA | ||
| PCL | C | 1.00 ± 0.06 | 1.47 ± 0.15 | 1.00 ± 0.16 | 1.29 ± 0.01 | 1.00 ± 0.01 | 1.07 ± 0.01 | 1.00 ± 0.03 | 1.69 ± 0.30 |
| SIN1 | 1.00 ± 0.03 | 1.37 ± 0.05 | 1.37 ± 0.20 | 1.78 ± 0.06 | 0.65 ± 0.02 | 0.84 ± 0.02 | 1.51 ± 0.01 | 1.72 ± 0.01 | |
| KO2 | 1.17 ± 0.03 | i.d. | 1.29 ± 0.30 | i.d. | 1.28 ± 0.01 | 1.01 ± 0.01 | 1.10 ± 0.16 | 1.63 ± 0.01 | |
| SIN1+SOD | 1.13 ± 0.08 | 1.35 ± 0.07 | 1.44 ± 0.19 | 1.70 ± 0.01 | 0.75 ± 0.01 | 0.87 ± 0.02 | 1.92 ± 0.10 | 1.34 ± 0.01 | |
| KO2+SOD | 0.95 ± 0.19 | i.d. | 1.02 ± 0.03 | 1.92 ± 0.01 | 1.20 ± 0.03 | 1.23 ± 0.02 | 0.99 ± 0.03 | 1.50 ± 0.21 | |
| PCPGL 10% | C | 1.00 ± 0.11 | 1.15 ± 0.10 | 1.00 ± 0.09 | 0.99 ± 0.13 | 1.00 ± 0.08 | 1.11 ± 0.06 | 1.00 ± 0.09 | 1.00 ± 0.08 |
| SIN1 | 1.06 ± 0.18 | 1.27 ± 0.09 | 1.07 ± 0.09 | 1.08 ± 0.09 | 1.09 ± 0.13 | 1.69 ± 0.06 | 0.55 ± 0.01 | 1.35 ± 0.14 | |
| KO2 | 1.06 ± 0.13 | 1.24 ± 0.26 | 1.07 ± 0.11 | 1.01 ± 0.14 | 1.13 ± 0.14 | 1.14 ± 0.04 | 0.97 ± 0.06 | 1.07 ± 0.02 | |
| SIN1+SOD | 1.17 ± 0.18 | 1.18 ± 0.06 | 1.08 ± 0.09 | 1.19 ± 0.08 | 1.04 ± 0.12 | 1.00 ± 0.14 | 1.13 ± 0.01 | 1.07 ± 0.14 | |
| KO2+SOD | 1.20 ± 0.05 | 1.20 ± 0.14 | 1.24 ± 0.10 | 1.21 ± 0.15 | 1.00 ± 0.10 | 1.02 ± 0.06 | 1.07 ± 0.17 | 1.00 ± 0.41 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mano, C.M.; Guaratini, T.; Cardozo, K.H.M.; Colepicolo, P.; Bechara, E.J.H.; Barros, M.P. Astaxanthin Restrains Nitrative-Oxidative Peroxidation in Mitochondrial-Mimetic Liposomes: A Pre-Apoptosis Model. Mar. Drugs 2018, 16, 126. https://doi.org/10.3390/md16040126
Mano CM, Guaratini T, Cardozo KHM, Colepicolo P, Bechara EJH, Barros MP. Astaxanthin Restrains Nitrative-Oxidative Peroxidation in Mitochondrial-Mimetic Liposomes: A Pre-Apoptosis Model. Marine Drugs. 2018; 16(4):126. https://doi.org/10.3390/md16040126
Chicago/Turabian StyleMano, Camila M., Thais Guaratini, Karina H. M. Cardozo, Pio Colepicolo, Etelvino J. H. Bechara, and Marcelo P. Barros. 2018. "Astaxanthin Restrains Nitrative-Oxidative Peroxidation in Mitochondrial-Mimetic Liposomes: A Pre-Apoptosis Model" Marine Drugs 16, no. 4: 126. https://doi.org/10.3390/md16040126
APA StyleMano, C. M., Guaratini, T., Cardozo, K. H. M., Colepicolo, P., Bechara, E. J. H., & Barros, M. P. (2018). Astaxanthin Restrains Nitrative-Oxidative Peroxidation in Mitochondrial-Mimetic Liposomes: A Pre-Apoptosis Model. Marine Drugs, 16(4), 126. https://doi.org/10.3390/md16040126