Anticoagulant and Antithrombotic Properties of Three Structurally Correlated Sea Urchin Sulfated Glycans and Their Low-Molecular-Weight Derivatives


Abstract
:
1. Introduction
2. Results and Discussion
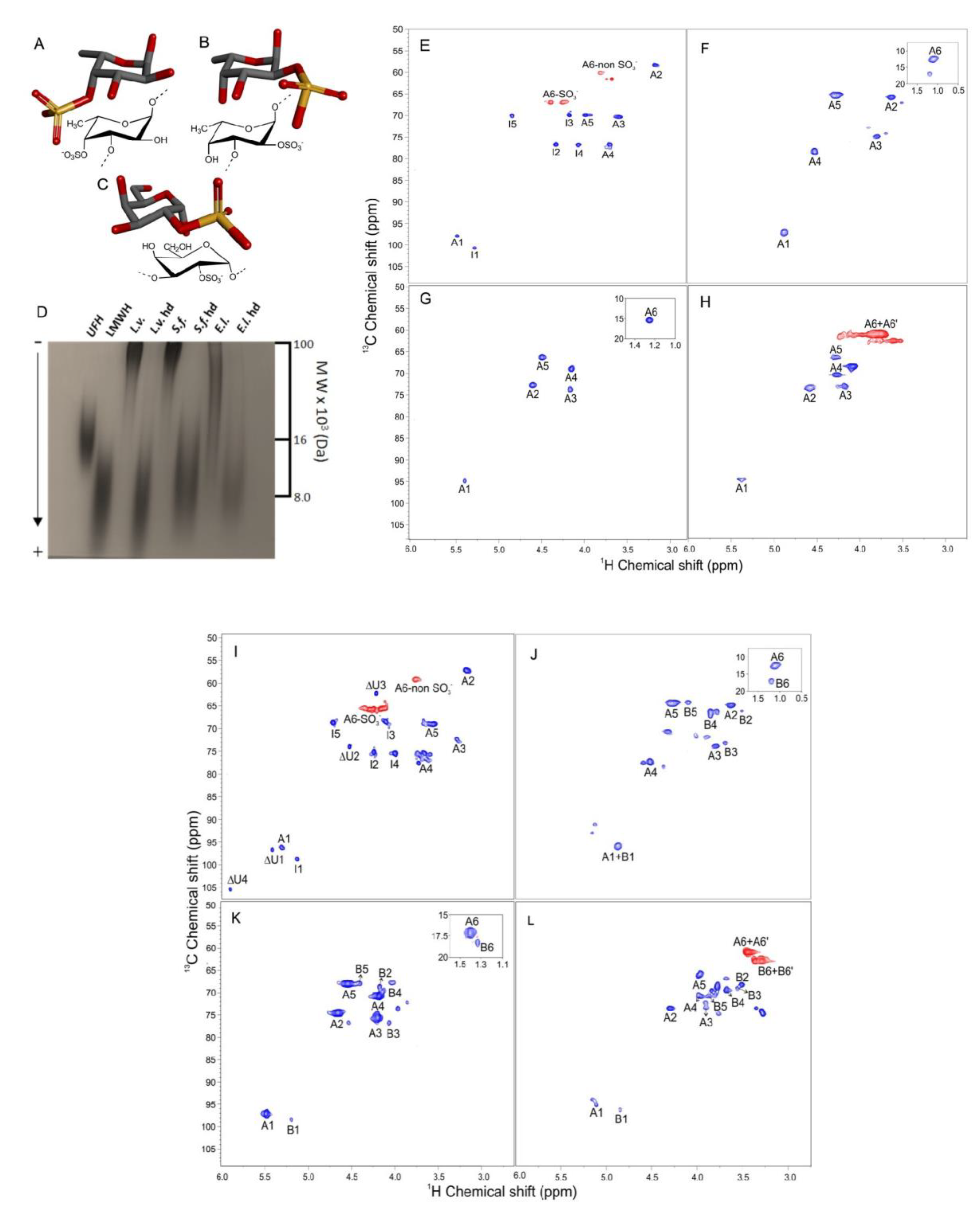
2.1. Compounds and Structural Integrity
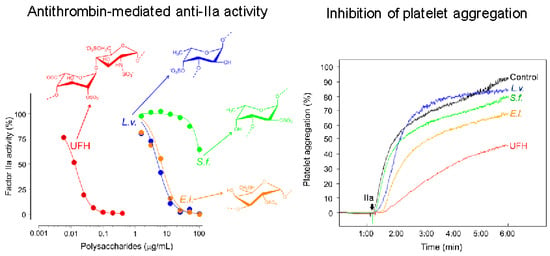
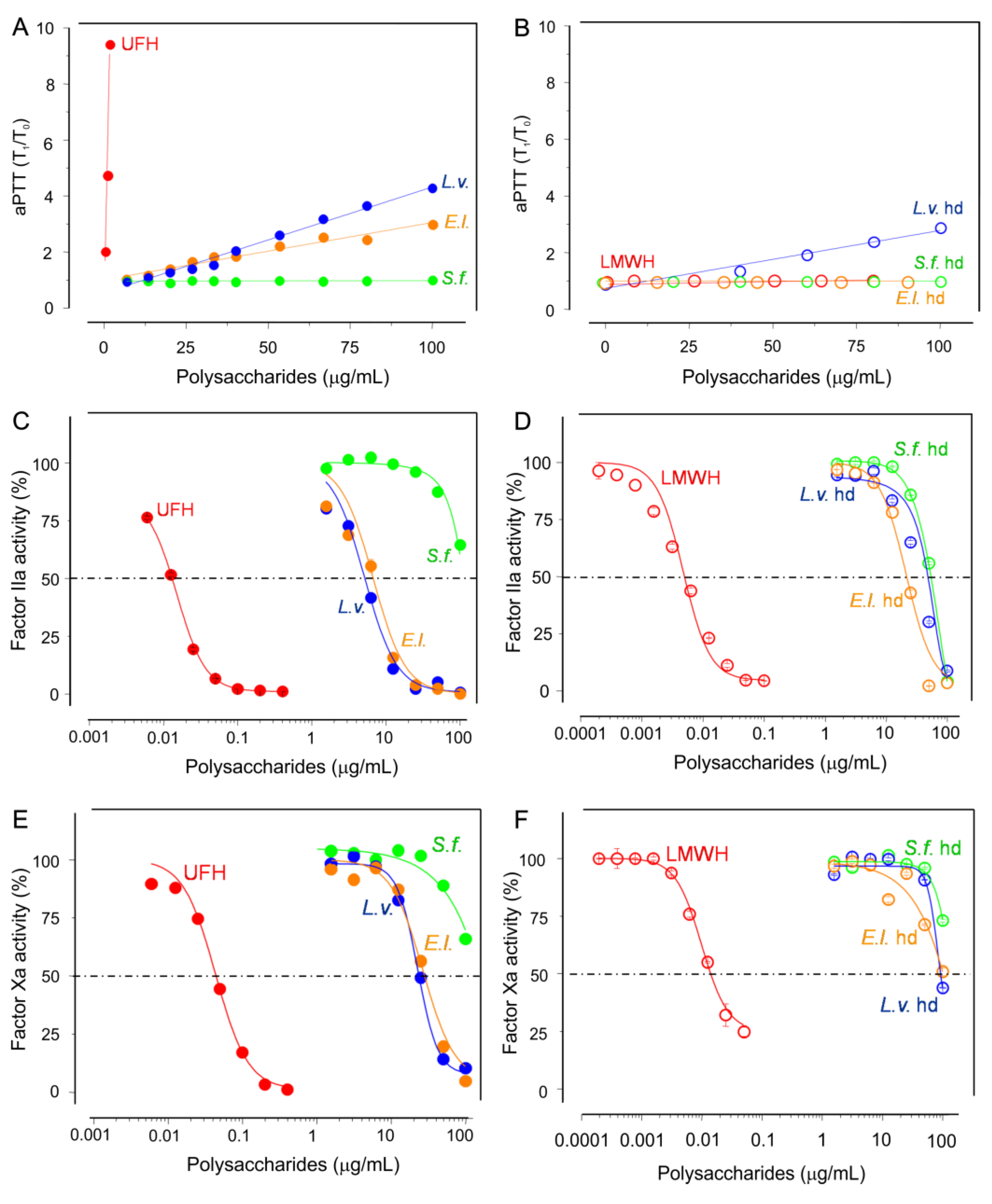
2.2. Anticoagulation
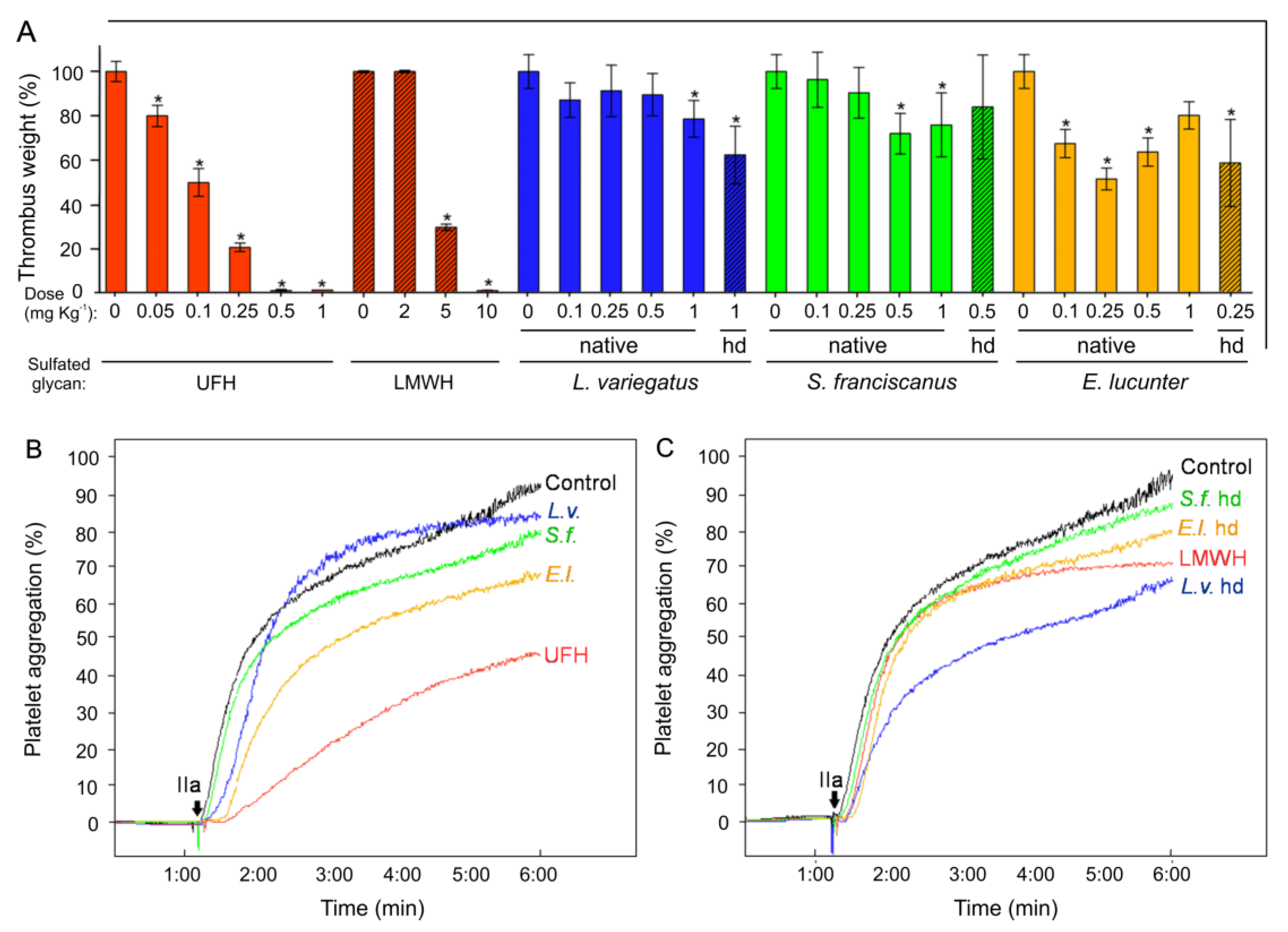
2.3. Antithrombosis
2.4. Platelet Aggregation
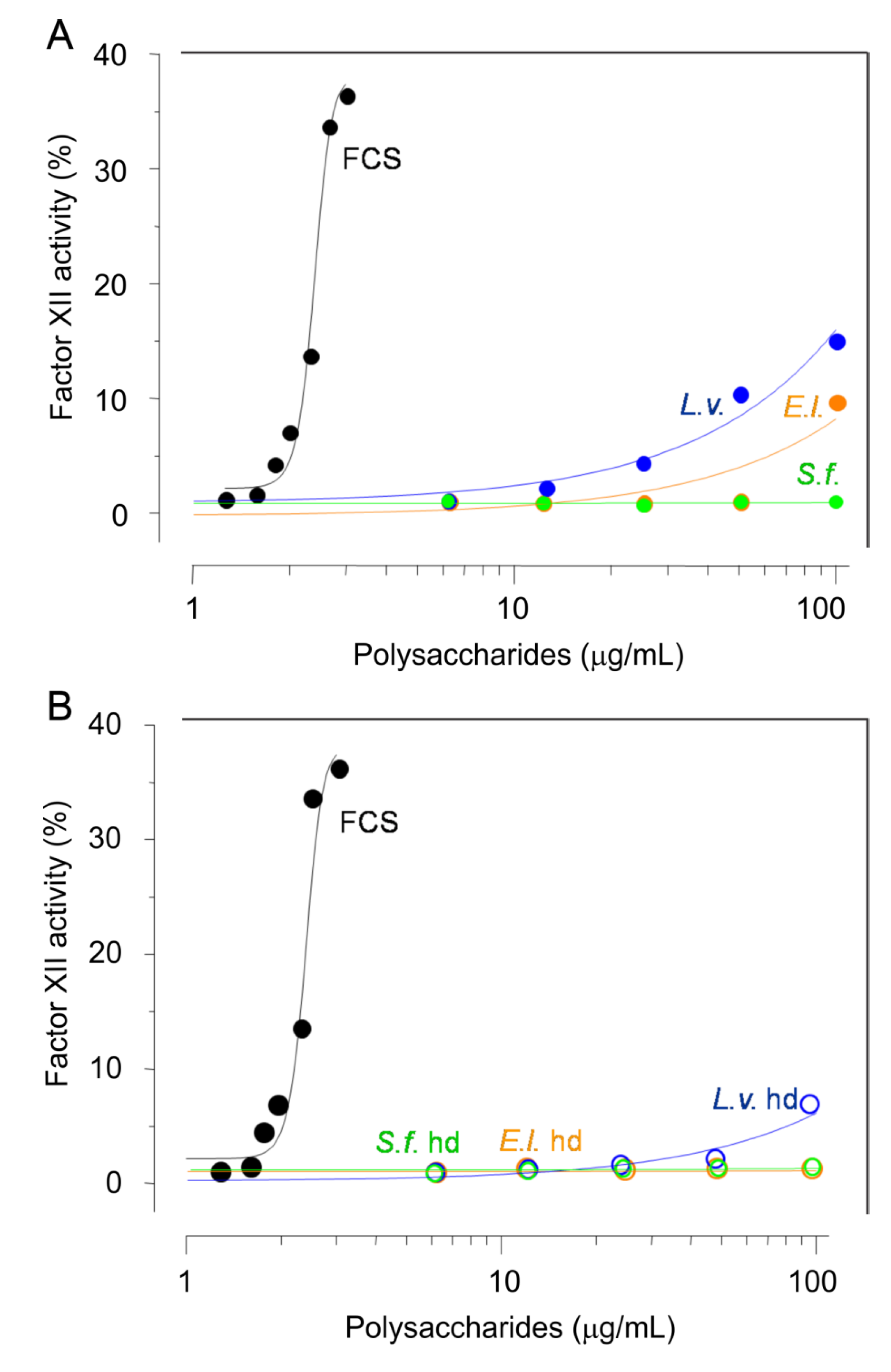
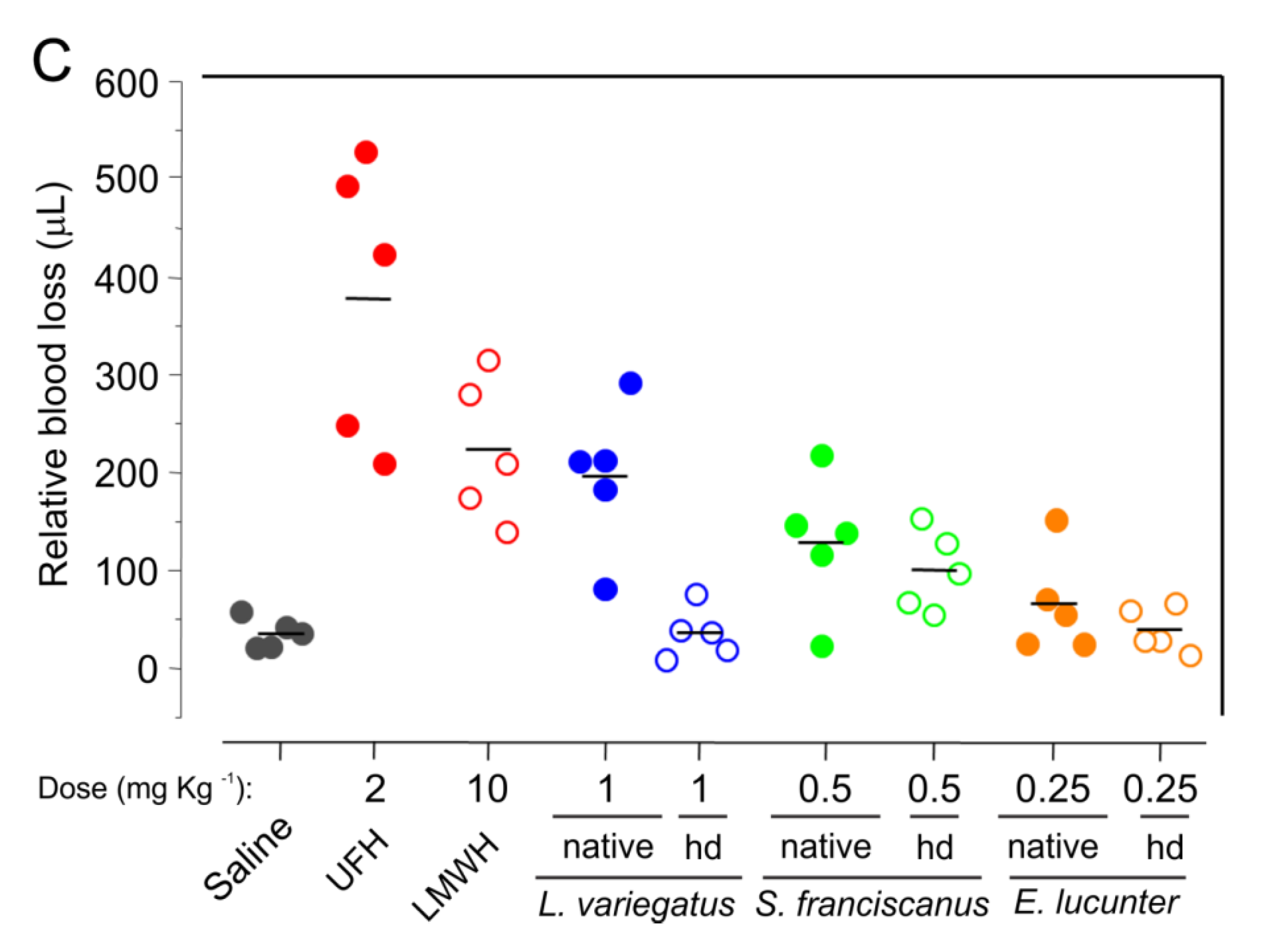
2.5. Potential Side Effects
3. Materials and Methods
3.1. Purification of the Sea Urchin Sulfated Glycans
3.2. Production of the Low-MW Derivatives
3.3. PAGE
3.4. NMR 1H-13C Dept-HSQC
3.5. Activated Partial Thromboplastin Time (aPTT)
3.6. Inhibition of Thrombin or Factor Xa by Antithrombin in the Presence of Sulfated Glycans
3.7. Venous Thrombosis
3.8. Platelet Aggregation
3.9. Bleeding
3.10. Activation of Factor XII
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raskob, G.E.; Angchaisuksiri, P.; Blanco, A.N.; Buller, H.; Gallus, A.; Hunt, B.J.; Hylek, E.M.; Kakkar, A.; Konstantinides, S.V.; McCumber, M.; et al. Thrombosis: A major contributor to global disease burden. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2363–2371. [Google Scholar] [CrossRef] [PubMed]
- The Top 10 Causes of Death. Available online: http://www.who.int/mediacentre/factsheets/fs310/en/ (accessed on 15 July 2018).
- Spyropoulos, A.C. Brave new world: The current and future use of novel anticoagulants. Thromb. Res. 2008, 123, S29–S35. [Google Scholar] [CrossRef] [PubMed]
- Mannucci, P.M.; Franchini, M. Old and new anticoagulant drugs: A minireview. Ann. Med. 2011, 43, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Fanikos, J.; Cina, J.L.; Baroletti, S.; Fiumara, K.; Matta, L.; Goldhaber, S.Z. Adverse drug events in hospitalized cardiac patients. Am. J. Cardiol. 2007, 100, 1465–1469. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.J.; Cohen, M.R.; Furberg, C.D. Serious adverse drug events reported to the Food and Drug Administration, 1998–2005. Arch. Intern. Med. 2007, 167, 1752–1759. [Google Scholar] [CrossRef] [PubMed]
- Mulloy, B.; Hogwood, J.; Gray, E.; Lever, R.; Page, C.P. Pharmacology of heparin and related drugs. Pharmacol. Rev. 2016, 68, 76–141. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Majeed, A.; Powell, R. Heparin induced thrombocytopenia: Diagnosis and management update. Postgrad. Med. J. 2007, 83, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Baroletti, S.A.; Goldhaber, S.Z. Heparin-induced thrombocytopenia. Circulation 2006, 114, e355–e356. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Lassepas, M.; Quinones, M.R. Heparin therapy for stroke: Hemorrhagic complications and risk factors for intracerebral hemorrhage. Neurology 1984, 34, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Clark, W.M.; Madden, K.P.; Lyden, P.D.; Zivin, J.A. Cerebral hemorrhagic risk of aspirin or heparin therapy with thrombolytic treatment in rabbits. Stroke 1991, 22, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Fucanomics and galactanomics: Current status in drug discovery, mechanisms of action and role of the well-defined structures. Biochim. Biophys. Acta 2012, 1820, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Marine non-glycosaminoglycan sulfated glycans as potential pharmaceuticals. Pharmaceuticals 2015, 8, 848–864. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. A dilemma in the glycosaminoglycan-based therapy: Synthetic or naturally unique molecules? Med. Res. Rev. 2015, 35, 1195–1219. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Holothurian fucosylated chondroitin sulfate. Mar. Drugs 2014, 12, 232–254. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Medical gains of chondroitin sulfate upon fucosylation. Curr. Med. Chem. 2015, 22, 4166–4176. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.S.; Vilela-Silva, A.C.E.S.; Valente, A.P.; Mourão, P.A.S. A 2-sulfated, 3-linked α-L-galactan is an anticoagulant polysaccharide. Carbohydr. Res. 2002, 337, 2231–2238. [Google Scholar] [CrossRef]
- Pereira, M.S.; Melo, F.R.; Mourão, P.A.S. Is there a correlation between structure and anticoagulant action of sulfated galactans and sulfated fucans? Glycobiology 2002, 12, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, R.J.C.; Santos, G.R.C.; Mourão, P.A.S. Effects of polysaccharides enriched in 2,4-disulfated fucose units on coagulation, thrombosis and bleeding. Practical and conceptual implications. Thromb. Haemost. 2009, 102, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.J.C.; Mourão, P.A.S. Fucosylated chondroitin sulfate as a new oral antithrombotic agent. Thromb. Haemost. 2006, 96, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.J.C.; Sucupira, I.D.; Oliveira, S.N.M.C.G.; Santos, G.R.C.; Mourão, P.A.S. Improved anticoagulant effect of fucosylated chondroitin sulfate orally administered as gastro-resistant tablets. Thromb. Haemost. 2017, 117, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Cinelli, L.P.; Castro, M.O.; Santos, L.L.; Garcia, C.R.; Vilela-Silva, A.C.E.S.; Mourão, P.A.S. Expression of two different sulfated fucans by females of Lytechinus variegatus may regulate the seasonal variation in the fertilization of the sea urchin. Glycobiology 2007, 17, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Vilela-Silva, A.C.E.S.; Alves, A.P.; Valente, A.P.; Vacquier, V.D.; Mourão, P.A.S. Structure of the sulfated alpha-L-fucan from the egg jelly coat of the sea urchin Strongylocentrotus franciscanus: Patterns of preferential 2-O- and 4-O-sulfation determine sperm cell recognition. Glycobiology 1999, 9, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Alves, A.P.; Mulloy, B.; Diniz, J.A.; Mourão, P.A.S. Sulfated polysaccharides from the egg jelly layer are species-specific inducers of acrosomal reaction in sperms of sea urchins. J. Biol. Chem. 1997, 272, 6965–6971. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, I.N.L.; Wang, X.; Glushka, J.N.; Santos, G.R.C.; Valente, A.P.; Prestegard, J.H.; Woods, R.J.; Mourão, P.A.S.; Pomin, V.H. Impact of sulfation pattern on the conformation and dynamics of sulfated fucan oligosaccharides as revealed by NMR and MD. Glycobiology 2015, 25, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, I.N.L.; Vilela-Silva, A.C.E.S.; Pomin, V.H. Oligosaccharides from the 3-linked 2-sulfated alpha-L-fucan and alpha-l-galactan show similar conformations but different dynamics. Glycobiology 2016, 26, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Anticoagulant motifs of marine sulfated glycans. Glycoconj. J. 2014, 31, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Quinderé, A.L.; Santos, G.R.; Oliveira, S.N.; Glauser, B.F.; Fontes, B.P.; Queiroz, I.N.; Benevides, N.M.; Pomin, V.H.; Mourão, P.A. Is the antithrombotic effect of sulfated galactans independent of serpin? J. Thromb. Haemost. 2014, 12, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo, F.R.; Mourão, P.A. An algal sulfated galactan has an unusual dual effect on venous thrombosis due to activation of factor XII and inhibition of the coagulation proteases. Thromb. Haemost. 2008, 99, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Dual and antagonic therapeutic effects of sulfated glycans. Bioorg. Med. Chem. 2016, 24, 3965–3971. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, R.J.C.; Oliveira, S.N.M.C.G.; Pomin, V.H.; Mecawi, A.S.; Araujo, I.G.; Mourão, P.A.S. Effects of oversulfated and fucosylated chondroitin sulfates on coagulation. Challenges for the study of anticoagulant polysaccharides. Thromb. Haemost. 2010, 103, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Pereira, M.S.; Valente, A.P.; Tollefsen, D.M.; Pavão, M.S.; Mourão, P.A.S. Selective cleavage and anticoagulant activity of a sulfated fucan: Stereospecific removal of a 2-sulfate ester from the polysaccharide by mild acid hydrolysis, preparation of oligosaccharides, and heparin cofactor II-dependent anticoagulant activity. Glycobiology 2005, 15, 369–681. [Google Scholar] [CrossRef] [PubMed]
- Wessler, S.; Reimer, S.M.; Sheps, M.C. Biologic assay of a thrombosis-inducing activity in human serum. J. Appl. Physiol. 1959, 14, 943–946. [Google Scholar] [CrossRef] [PubMed]
- Herbert, J.M.; Bernat, A.; Maffrand, J.P. Importance of platelets in experimental venous thrombosis in the rat. Blood 1992, 80, 2281–2286. [Google Scholar] [PubMed]
- Fonseca, R.J.C.; Oliveira, S.N.M.C.G.; Melo, F.R.; Pereira, M.G.; Benevides, N.M.; Mourão, P.A.S. Slight differences in sulfation of algal galactans account for differences in their anticoagulant and venous antithrombotic activities. Thromb. Haemost. 2008, 99, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.F.; Guimarães, J.A.; Mourão, P.A.; Verli, H. Conformation of sulfated galactan and sulfated fucan in aqueous solutions: Implications to their anticoagulant activities. J. Mol. Graph. Model. 2007, 26, 391–399. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycan | AT/IIa | AT/Xa |
|---|---|---|
| UFH | 210 | 204 |
| LMWH | 35 | 100 |
| L.v. | 0.44 | 0.29 |
| L.v. hd | 0.07 | 0.06 |
| S.f. | 0.02 | 0.05 |
| S.f. hd | 0.05 | 0.04 |
| E.l. | 0.56 | 0.27 |
| E.l. hd | 0.12 | 0.08 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasconcelos, A.A.; Sucupira, I.D.; Guedes, A.L.; Queiroz, I.N.; Frattani, F.S.; Fonseca, R.J.; Pomin, V.H. Anticoagulant and Antithrombotic Properties of Three Structurally Correlated Sea Urchin Sulfated Glycans and Their Low-Molecular-Weight Derivatives. Mar. Drugs 2018, 16, 304. https://doi.org/10.3390/md16090304
Vasconcelos AA, Sucupira ID, Guedes AL, Queiroz IN, Frattani FS, Fonseca RJ, Pomin VH. Anticoagulant and Antithrombotic Properties of Three Structurally Correlated Sea Urchin Sulfated Glycans and Their Low-Molecular-Weight Derivatives. Marine Drugs. 2018; 16(9):304. https://doi.org/10.3390/md16090304
Chicago/Turabian StyleVasconcelos, Ariana A., Isabela D. Sucupira, Alessandra L. Guedes, Ismael N. Queiroz, Flavia S. Frattani, Roberto J. Fonseca, and Vitor H. Pomin. 2018. "Anticoagulant and Antithrombotic Properties of Three Structurally Correlated Sea Urchin Sulfated Glycans and Their Low-Molecular-Weight Derivatives" Marine Drugs 16, no. 9: 304. https://doi.org/10.3390/md16090304
APA StyleVasconcelos, A. A., Sucupira, I. D., Guedes, A. L., Queiroz, I. N., Frattani, F. S., Fonseca, R. J., & Pomin, V. H. (2018). Anticoagulant and Antithrombotic Properties of Three Structurally Correlated Sea Urchin Sulfated Glycans and Their Low-Molecular-Weight Derivatives. Marine Drugs, 16(9), 304. https://doi.org/10.3390/md16090304