Integrated Genomic and Metabolomic Approach to the Discovery of Potential Anti-Quorum Sensing Natural Products from Microbes Associated with Marine Samples from Singapore

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
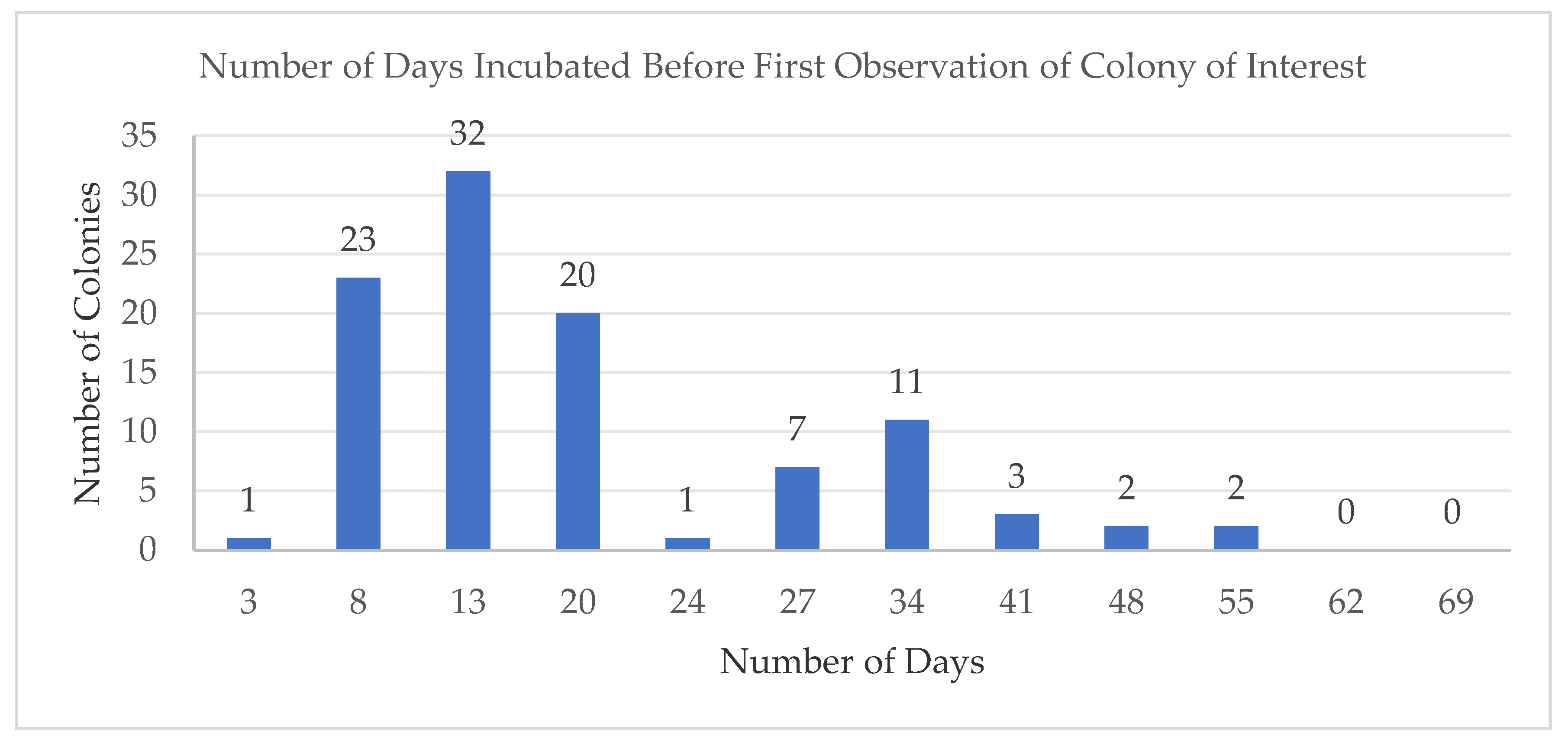
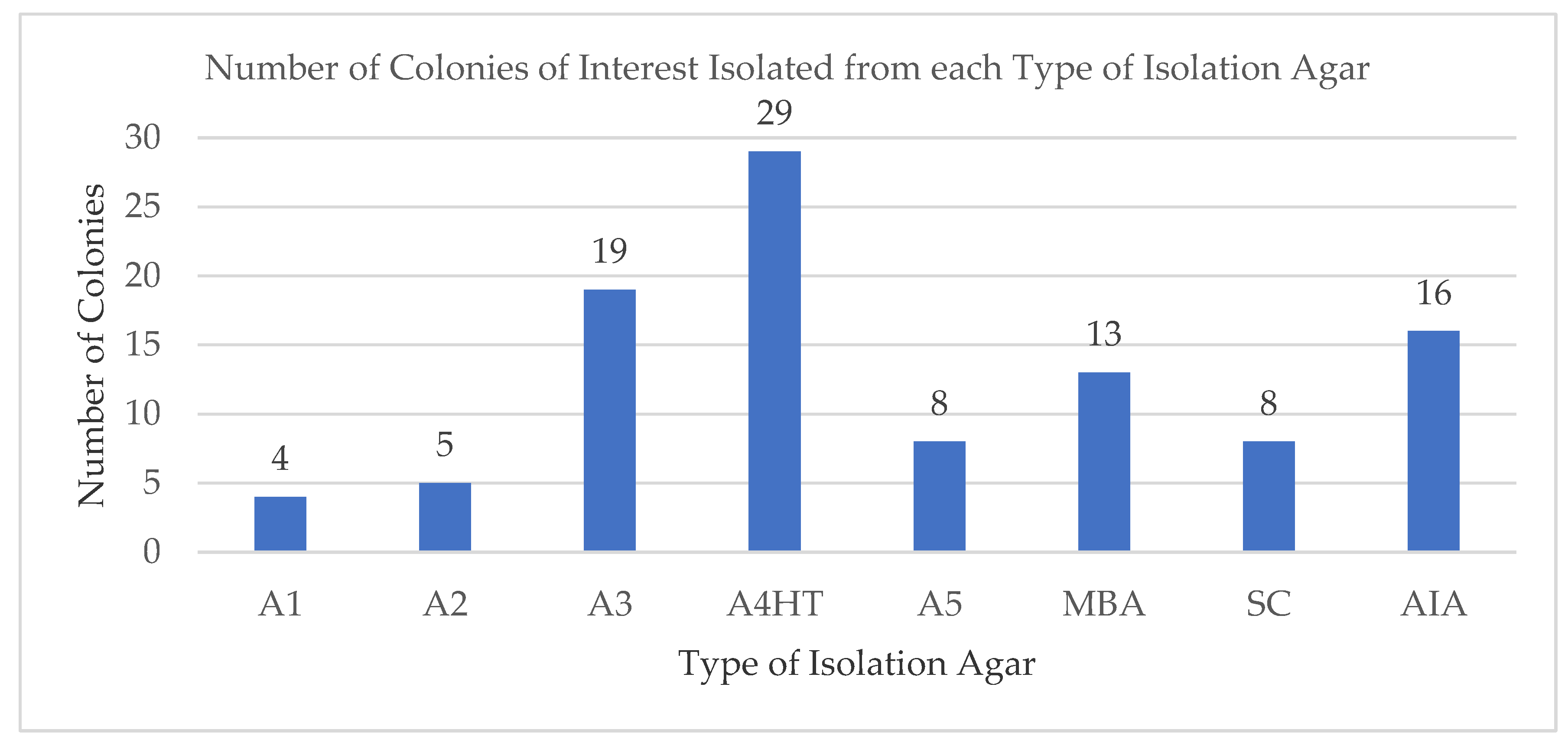
2.1. Isolated Microbes Associated with Deep Water Marine Samples
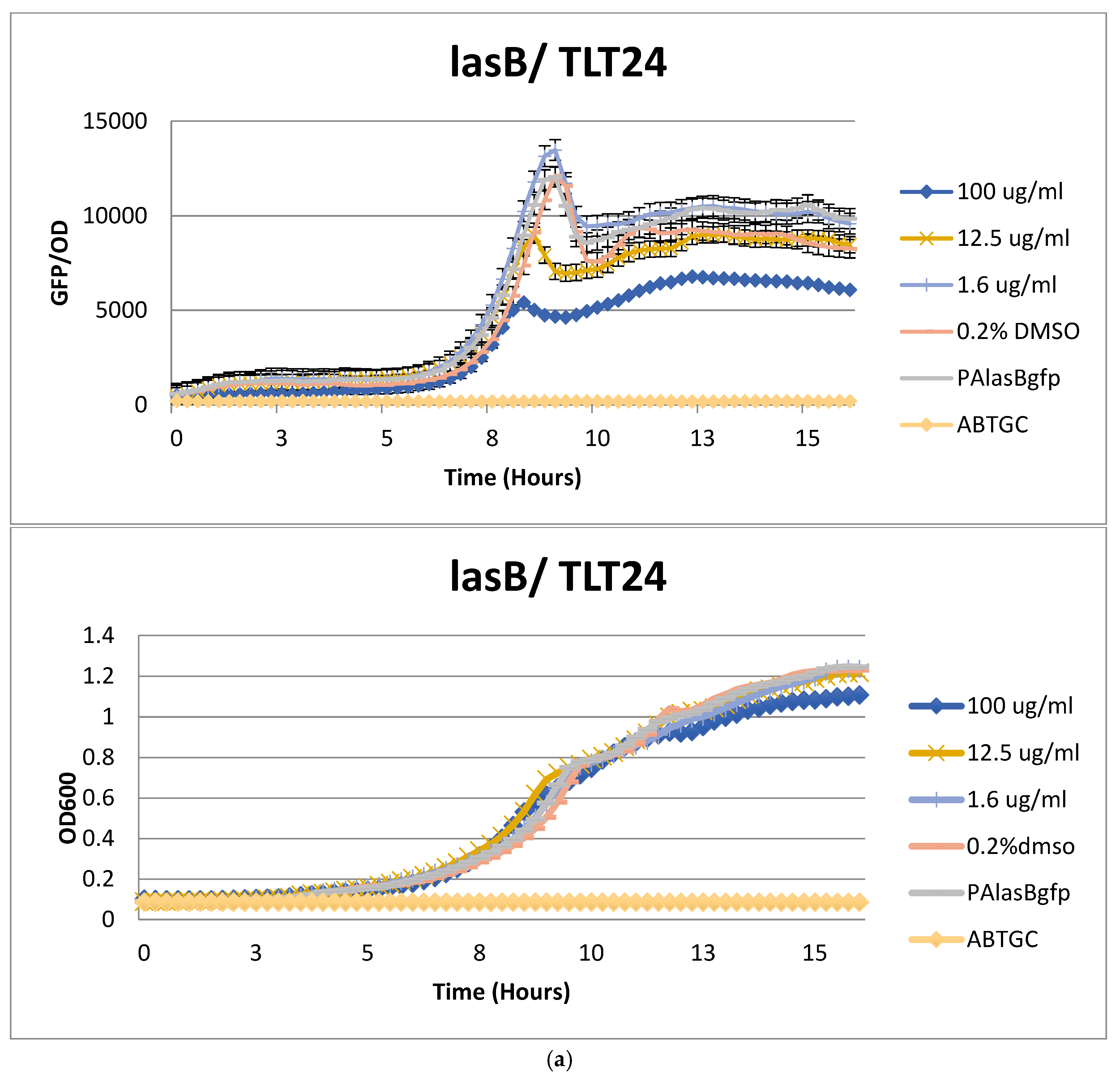
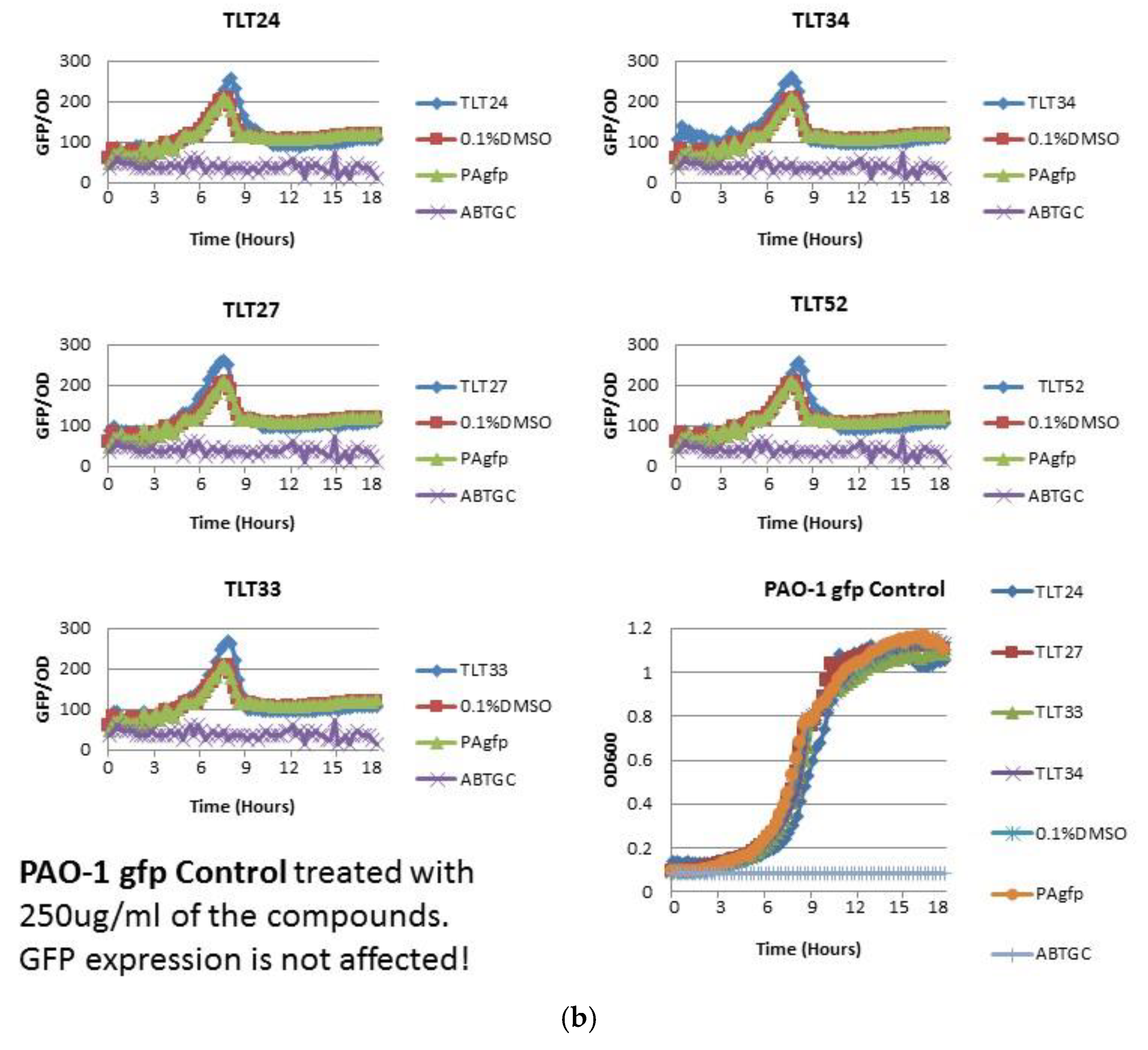
2.2. Anti-Quorum Sensing Activity of Marine Bacterial Extracts
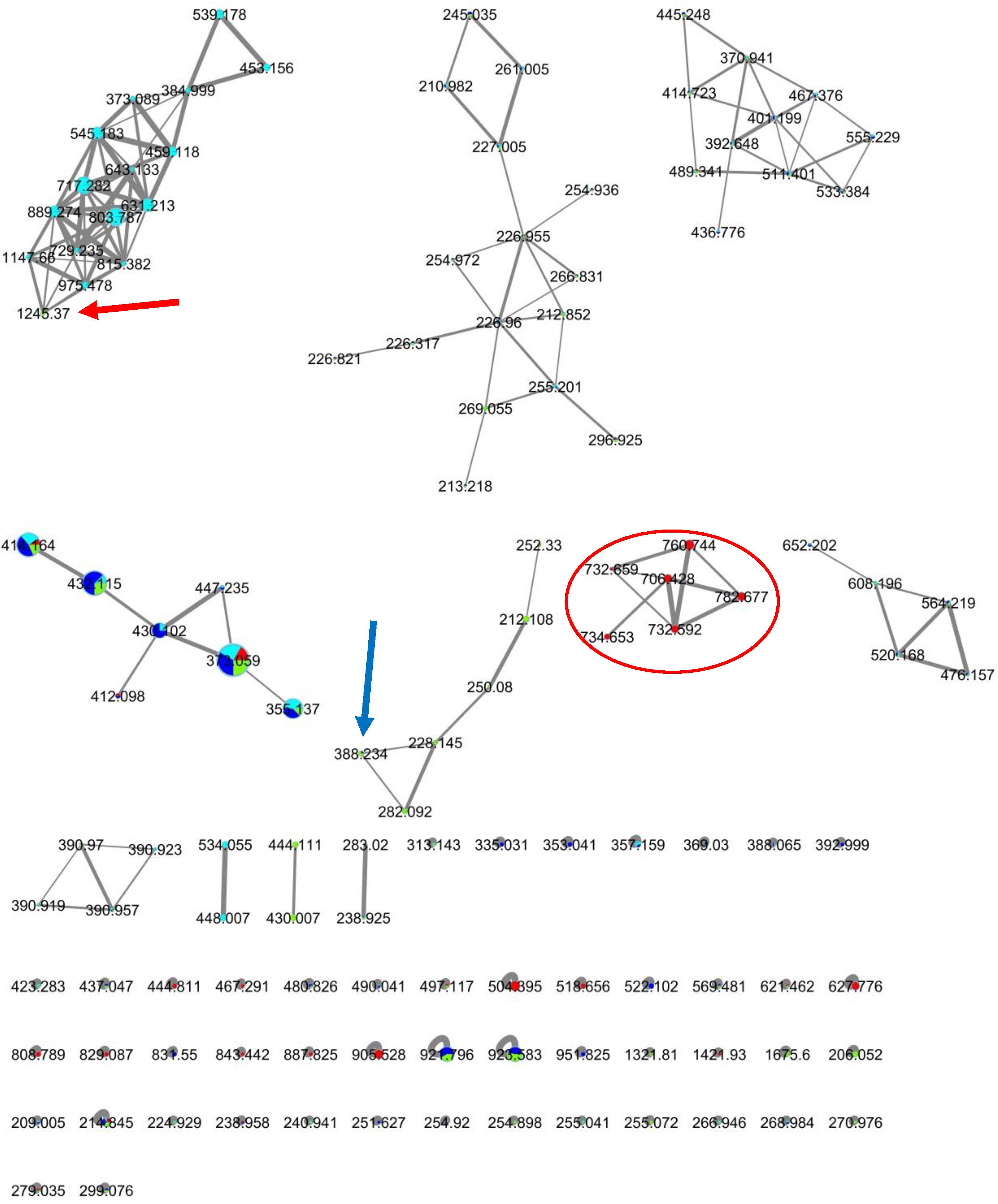
2.3. MS-Based Molecular Networking of Organic Extracts Derived from Selected Marine Bacterial Strains
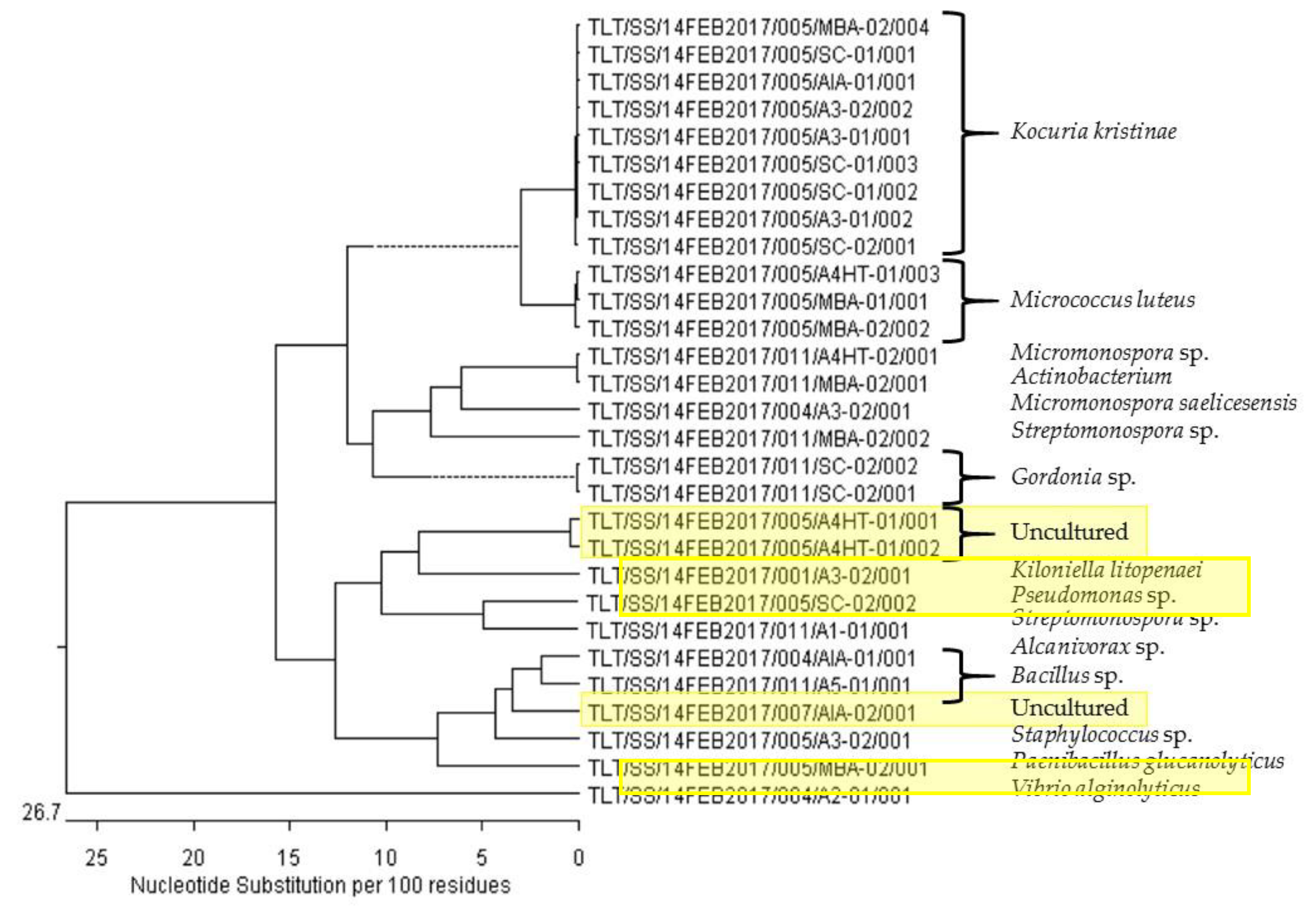
2.4. Phylogenetic Analysis of Selected Marine Bacterial Strains
2.5. Annotation of Biosynthetic Gene Clusters of Selected Marine Bacterial Genome
3. Experimental
3.1. Sample Collection and Processing
3.2. Cultivable Microbial Isolation
3.3. Molecular Identification and Phylogenetic Analysis
3.4. Organic Extract Preparation and Anti-Quorum Sensing Bioassay
3.5. Mass Spectrometry-Based Molecular Networking
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Obst, U.; Schwartz, T.; Volkmann, H. Antibiotic resistant pathogenic bacteria and their resistance genes in bacterial biofilms. Int. J. Artif. Organs. 2006, 29, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Escaich, S. Novel agents to inhibit microbial virulence and pathogenicity. Expert Opin. Ther. Pat. 2010, 20, 1401–1418. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Zhang, X.H. Quorum quenching agents: Resources for antivirulence therapy. Mar. Drugs 2014, 12, 3245–3282. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Tomita, H. Interference of bacterial cell-to-cell communication: A new concept of antimicrobial chemotherapy breaks antibiotic resistance. Front. Microbiol. 2013, 4, 114. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Coenye, T. Quorum sensing inhibitors as anti-biofilm agents. Curr. Pharm. Des. 2015, 21, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Rémy, B.; Mion, S.; Plener, L.; Elias, M.; Chabrière, E.; Daudé, D. Interference in bacterial quorum sensing: A biopharmaceutical perspective. Front. Pharmacol. 2018, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.R.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association—A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [PubMed]
- Hameş-Kocabaş, E.E.; Uzel, A. Isolation strategies of marine-derived actinomycetes from sponge and sediment samples. J. Microbiol. Methods 2012, 88, 342–347. [Google Scholar] [CrossRef]
- Saurav, K.; Costantino, V.; Venturi, V.; Steindler, L. Quorum sensing inhibitors from the sea discovered using bacterial N-acyl-homoserine lactone-based biosensors. Mar. Drugs 2017, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, A.; Josyula, V.R.; Hariharapura, R.C. Exploring the potential of marine microbes in clinical management of Alzheimer’s disease: A road map for bioprospecting and identifying promising isolates. Life Sci. 2018, 208, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the core microbiome in corals’ microbial soup. Trends Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Fieseler, L.; Wehrl, M.; Gernert, C.; Steinert, M.; Hacker, J.; Horn, M. Microbial diversity of marine sponges. Prog. Mol. Subcell. Biol. 2003, 37, 59–88. [Google Scholar]
- Wang, G. Diversity and biotechnological potential of the sponge-associated microbial consortia. J. Ind. Microbiol. Biotechnol. 2006, 33, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef]
- Leal, M.C.; Sheridan, C.; Osinga, R.; Dionísio, G.; Rocha, R.J.; Silva, B.; Rosa, R.; Calado, R. Marine microorganism-invertebrate assemblages: Perspectives to solve the “supply problem” in the initial steps of drug discovery. Mar. Drugs 2014, 12, 3929–3952. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, C.M.; Marker, A.; Kurtböke, D.I. An overview on marine sponge-symbiotic bacteria as unexhausted sources for natural product discovery. Diversity 2017, 9, 40. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef]
- Haque, S.; Ahmad, F.; Dar, S.A.; Jawed, A.; Mandal, R.K.; Wahid, M.; Lohani, M.; Khan, S.; Singh, V.; Akhter, N. Development in strategies for quorum sensing virulence factor inhibition to combat bacterial drug resistance. Microb. Pathog. 2018, 121, 293–302. [Google Scholar] [CrossRef]
- Golberg, K.; Pavlov, V.; Marks, R.S.; Kushmaro, A. Coral-associated bacteria, quorum sensing disrupters, and the regulation of biofouling. Biofouling 2013, 29, 669–682. [Google Scholar] [CrossRef]
- Saurav, K.; Bar-Shalom, R.; Haber, M.; Burgsdorf, I.; Oliviero, G.; Costantino, V.; Morgenstern, D.; Steindler, L. In search of alternative antibiotic drugs: Quorum-quenching activity in sponges and their bacterial isolates. Front. Microbiol. 2016, 7, 416. [Google Scholar] [CrossRef] [PubMed]
- Allard, P.M.; Peresse, T.; Bisson, J.; Gindro, K.; Marcourt, L.; Pham, V.C.; Roussi, F.; Litaudon, M.; Wolfender, J.L. Integration of molecular networking and in-silico MS/MS fragmentation for natural products dereplication. Anal. Chem. 2016, 88, 3317–3323. [Google Scholar] [CrossRef] [PubMed]
- Magarvey, N.A.; Keller, J.M.; Bernan, V.; Dworkin, M.; Sherman, D.H. Isolation and characterization of novel marine-derived actinomycete taxa rich in bioactive metabolites. Appl. Environ. Microbiol. 2004, 70, 7520–7529. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Nam, S.J.; Paul, L.; Beatty, D.; Kauffman, C.A.; Jensen, P.R.; Fenical, W. Previously uncultured marine bacteria linked to novel alkaloid production. Chem. Biol. 2015, 22, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.R.; Crüsemann, M.; Lechner, A.; Sarkar, A.; Li, J.; Ziemert, N.; Wang, M.; Bandeira, N.; Moore, B.S.; Dorrestein, P.C.; et al. Molecular networking and pattern-based genome mining improves discovery of biosynthetic gene clusters and their products from Salinispora species. Chem. Biol. 2015, 22, 460–471. [Google Scholar] [CrossRef]
- Maansson, M.; Vynne, N.G.; Klitgaard, A.; Nybo, J.L.; Melchiorsen, J.; Nguyen, D.D.; Sanchez, L.M.; Ziemert, N.; Dorrestein, P.C.; Andersen, M.R.; et al. An integrated metabolomic and genomic mining workflow to uncover the biosynthetic potential of bacteria. mSystems 2016, 1, e00028-15. [Google Scholar] [CrossRef]
- Webster, N.S.; Wilson, K.J.; Blackall, L.L.; Hill, R.T. Phylogenetic diversity of bacteria associated with the marine sponge Rhopaloeides odorabile. Appl. Environ. Microbiol. 2001, 67, 434–444. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Bjarnsholt, T.; Skindersoe, M.E.; Hentzer, M.; Kristoffersen, P.; Kote, M.; Nielsen, J.; Eberl, L.; Givskov, M. Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. J. Bacteriol. 2005, 187, 1799–1814. [Google Scholar] [CrossRef]
- Fong, J.; Yuan, M.; Jakobsen, T.H.; Mortensen, K.T.; Delos Santos, M.M.S.; Chua, S.L.; Yang, L.; Tan, C.H.; Nielsen, T.E.; Givskow, M. Disulfide bond-containing ajoene analogues as novel quorum sensing inhibitors of Pseudomonas aeruginosa. J. Med Chem. 2017, 60, 215–227. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with GNPS. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Garza, D.R.; Dutilh, B.E. From cultured to uncultured genome sequences: Metagenomics and modeling microbial ecosystems. Cell. Mol. Life Sci. 2015, 72, 4287–4308. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de los Santos, E.L.C.; Kim, H.U.; Medema, M.H.; et al. antiSMASH 4.0—Improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Winnikoff, J.R.; Glukhov, E.; Watrous, J.; Dorrestein, P.C.; Gerwick, W.H. Quantitative molecular networking to profile marine cyanobacterial metabolomes. J. Antibiot. 2014, 67, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Kleigrewe, K.; Almaliti, J.; Tian, I.Y.; Kinnel, R.B.; Korobeynikov, A.; Monroe, E.A.; Duggan, B.M.; Marzo, V.D.; Sherman, D.H.; Dorrestein, P.C.; et al. Combining mass spectrometric metabolic profiling with genomic analysis: A powerful approach for discovering natural products from cyanobacteria. J. Nat. Prod. 2015, 78, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Purves, K.; Macintyre, L.; Brennan, D.; Hreggviosson, G.O.; Kuttner, E.; Asgeirsdottir, M.E.; Young, L.C.; Green, D.H.; Edrada-Ebel, R.; Duncan, K.R. Using molecular networking for microbial secondary metabolite bioprospecting. Metabolites 2016, 6, 2. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample # | Marine Bacterial Code Number | Source | Base Pairs # | Identification (% Similarity) | NCBI Accession # |
|---|---|---|---|---|---|
| 1 | TLT/SS/14FEB2017/001/A3-02/001 | Xestospongia sp. | 1433 | Kiloniella litopenaei (98) | LT717346 |
| 8 | TLT/SS/14FEB2017/004/A2-01/001 | Stelletta sp. | 536 | Vibrio alginolyticus (90) | CP017919 |
| 10 | TLT/SS/14FEB2017/004/A3-02/001 | Stelletta sp. | 1400 | Micromonospora saelicesensis (97) | KM37003 |
| 16 | TLT/SS/14FEB2017/004/AIA-01/001 | Stelletta sp. | 1484 | Bacillus sp. (99) | KC953600 |
| 20 | TLT/SS/14FEB2017/005/A3-01/001 | Geodia sp. | 1467 | Kocuria kristinae (99) | KR230389 |
| 21 | TLT/SS/14FEB2017/005/A3-01/002 | Geodia sp. | 1467 | Kocuria kristinae (99) | KR230389 |
| 22 | TLT/SS/14FEB2017/005/A3-02/001 | Geodia sp. | 1491 | Staphylococcus sp. (99) | FR839669 |
| 23 | TLT/SS/14FEB2017/005/A3-02/002 | Geodia sp. | 1466 | Kocuria kristinae (99) | KR230389 |
| 24 | TLT/SS/14FEB2017/005/A4HT-01/001 | Geodia sp. | 1398 | Uncultured (98) | EF574305 |
| 25 | TLT/SS/14FEB2017/005/A4HT-01/002 | Geodia sp. | 1384 | Uncultured (96) | KJ814073 |
| 26 | TLT/SS/14FEB2017/005/A4HT-01/003 | Geodia sp. | 1449 | Micrococcus luteus (99) | KP345957 |
| 29 | TLT/SS/14FEB2017/005/MBA-01/001 | Geodia sp. | 1450 | Micrococcus luteus (99) | KT805418 |
| 30 | TLT/SS/14FEB2017/005/MBA-02/001 | Geodia sp. | 1489 | Paenibacillus glucanolyticus (99) | CP015286 |
| 31 | TLT/SS/14FEB2017/005/MBA-02/002 | Geodia sp. | 1447 | Micrococcus luteus (99) | KF993668 |
| 33 | TLT/SS/14FEB2017/005/MBA-02/004 | Geodia sp. | 1465 | Kocuria kristinae (99) | KR230389 |
| 34 | TLT/SS/14FEB2017/005/SC-01/001 | Geodia sp. | 1468 | Kocuria kristinae (99) | DQ158132 |
| 35 | TLT/SS/14FEB2017/005/SC-01/002 | Geodia sp. | 1461 | Kocuria kristinae (99) | KR230389 |
| 36 | TLT/SS/14FEB2017/005/SC-01/003 | Geodia sp. | 1455 | Kocuria kristinae (99) | DQ158132 |
| 37 | TLT/SS/14FEB2017/005/SC-02/001 | Geodia sp. | 1831 | Kocuria kristinae (98) | KF075509 |
| 38 | TLT/SS/14FEB2017/005/SC-02/002 | Geodia sp. | 1475 | Pseudomonas sp. (99) | KT034415 |
| 39 | TLT/SS/14FEB2017/005/AIA-01/001 | Geodia sp. | 1466 | Kocuria sp. (99) | KR230389 |
| 52 | TLT/SS/14FEB2017/007/AIA-02/001 | Coelocarteria singaporensis | 1481 | Uncultured (99) | KX859231 |
| 64 | TLT/SS/14FEB2017/011/A1-01/001 | Sediment | 1477 | Alcanivorax sp. (99) | KU954765 |
| 66 | TLT/SS/14FEB2017/011/A4HT-02/001 | Sediment | 1447 | Micromonospora sp. (99) | AB738798 |
| 67 | TLT/SS/14FEB2017/011/A5-01/001 | Sediment | 1486 | Bacillus sp. (99) | AJ438301 |
| 68 | TLT/SS/14FEB2017/011/MBA-02/001 | Sediment | 1445 | Actinobacterium (99) | JN049491 |
| 69 | TLT/SS/14FEB2017/011/MBA-02/002 | Sediment | 1466 | Streptomonospora sp. (99) | JX007947 |
| 70 | TLT/SS/14FEB2017/011/SC-02/001 | Sediment | 1455 | Gordonia sp. (99) | EU590659 |
| 71 | TLT/SS/14FEB2017/011/SC-02/002 | Sediment | 1456 | Gordonia sp. (99) | CP002907 |
| TLT/SS/14FEB2017/005/A4HT-01/001 (#24) | ||||
| Cluster | Type | From | To | Most Similar Known Cluster |
| Cluster 1 | Other | 704,902 | 746,323 | Bacilysin_biosynthetic_gene_cluster |
| (85% of genes show similarity) | ||||
| Cluster 2 | Terpene-Siderophore | 57,654 | 91,248 | Carotenoid_biosynthetic_gene_cluster |
| (50% of genes show similarity) | ||||
| Cluster 3 | Bacteriocin | 442,553 | 452,879 | - |
| Cluster 4 | Terpene | 56,166 | 78,121 | - |
| Cluster 5 | T3pks | 116,521 | 156,058 | - |
| Cluster 6 | Nrps | 143,837 | 227,604 | Lichenysin_biosynthetic_gene_cluster |
| (85% of genes show similarity) | ||||
| TLT/SS/14FEB2017/007/AIA-02/001 (#52) | ||||
| Cluster | Type | From | To | Most Similar Known Cluster |
| Cluster 1 | Siderophore | 387,071 | 400,778 | Petrobactin_biosynthetic_gene_cluster (100% of genes show similarity) |
| Cluster 2 | Nrps | 687,959 | 737,684 | Bacillibactin_biosynthetic_gene_cluster (46% of genes show similarity) |
| Cluster 3 | Nrps | 57,214 | 116,594 | Polyoxypeptin_biosynthetic_gene_cluster (5% of genes show similarity) |
| Cluster 4 | Terpene | 35,609 | 57,462 | Molybdenum_cofactor_biosynthetic_gene_cluster (11% of genes show similarity) |
| Cluster 5 | Other | 96,159 | 139,740 | - |
| Cluster 6 | Bacteriocin | 156,762 | 170,627 | - |
| Cluster 7 | Nrps | 17,996 | 65,012 | - |
| Cluster 8 | Bacteriocin | 80,566 | 90,814 | - |
| Cluster 9 | Bacteriocin | 3212 | 13,541 | - |
| Cluster 10 | Nrps | 117,416 | 183,324 | - |
| Cluster 11 | Sactipeptide | 1 | 17,542 | Thurincin_H_biosynthetic_gene_cluster (100% of genes show similarity) |
| Cluster 12 | Arylpolyene-Nrps | 46,467 | 94,641 | - |
| Cluster 13 | Bacteriocin | 28,116 | 40,314 | - |
| Strains or Plasmids | Relevant Genotype and/or Characteristics |
|---|---|
| Strains | |
| PAO1 | ATCC Pseudomonas aeruginosa |
| PAO1-gfp | GFP-tagged wild-type Pseudomonas aeruginosa |
| PAO1-lasB-gfp | PAO1 containing lasB-gfp (ASV) translational reporter fusion |
| PAO1 ΔlasIΔrhlI | Gentamicin Resistance; PAO1 lasI and rhlI mutant |
| Plasmids | |
| PrhlA-gfp | Gentamicin Resistance/Carbenicillin Resistance; pUCPNotI-based plasmid carrying RlhRregulated rhlA-gfp (ASV) translational fusion |
| PpqsA-gfp | Gentamicin Resistance/Carbenicillin Resistance; pUCP22NotI-based plasmid carrying pqsA-gfp (ASV) transcriptional fusion |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ong, J.F.M.; Goh, H.C.; Lim, S.C.; Pang, L.M.; Chin, J.S.F.; Tan, K.S.; Liang, Z.-X.; Yang, L.; Glukhov, E.; Gerwick, W.H.; et al. Integrated Genomic and Metabolomic Approach to the Discovery of Potential Anti-Quorum Sensing Natural Products from Microbes Associated with Marine Samples from Singapore. Mar. Drugs 2019, 17, 72. https://doi.org/10.3390/md17010072
Ong JFM, Goh HC, Lim SC, Pang LM, Chin JSF, Tan KS, Liang Z-X, Yang L, Glukhov E, Gerwick WH, et al. Integrated Genomic and Metabolomic Approach to the Discovery of Potential Anti-Quorum Sensing Natural Products from Microbes Associated with Marine Samples from Singapore. Marine Drugs. 2019; 17(1):72. https://doi.org/10.3390/md17010072
Chicago/Turabian StyleOng, Ji Fa Marshall, Hui Chin Goh, Swee Cheng Lim, Li Mei Pang, Joyce Seow Fong Chin, Koh Siang Tan, Zhao-Xun Liang, Liang Yang, Evgenia Glukhov, William H. Gerwick, and et al. 2019. "Integrated Genomic and Metabolomic Approach to the Discovery of Potential Anti-Quorum Sensing Natural Products from Microbes Associated with Marine Samples from Singapore" Marine Drugs 17, no. 1: 72. https://doi.org/10.3390/md17010072
APA StyleOng, J. F. M., Goh, H. C., Lim, S. C., Pang, L. M., Chin, J. S. F., Tan, K. S., Liang, Z.-X., Yang, L., Glukhov, E., Gerwick, W. H., & Tan, L. T. (2019). Integrated Genomic and Metabolomic Approach to the Discovery of Potential Anti-Quorum Sensing Natural Products from Microbes Associated with Marine Samples from Singapore. Marine Drugs, 17(1), 72. https://doi.org/10.3390/md17010072