Antioxidative Peptides from Proteolytic Hydrolysates of False Abalone (Volutharpa ampullacea perryi): Characterization, Identification, and Molecular Docking




Abstract
1. Introduction
2. Results and Discussion
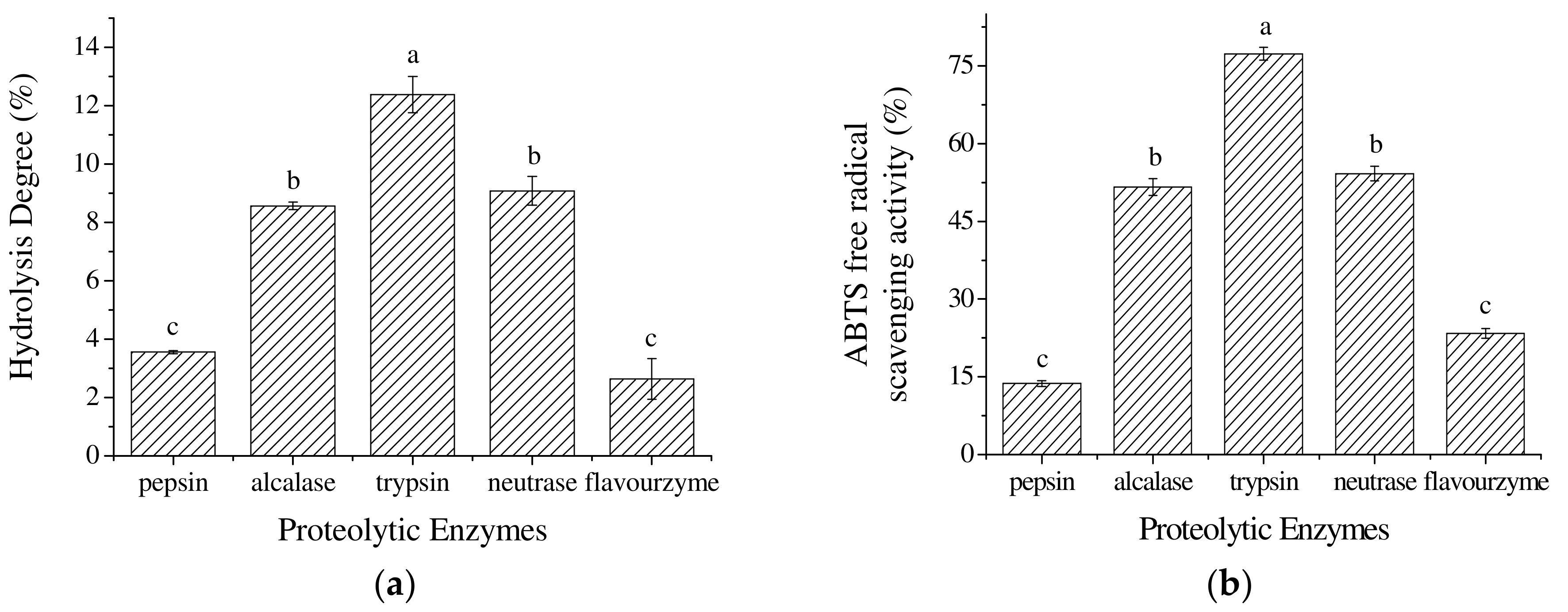
2.1. The Selection of Proteolytic Enzymes
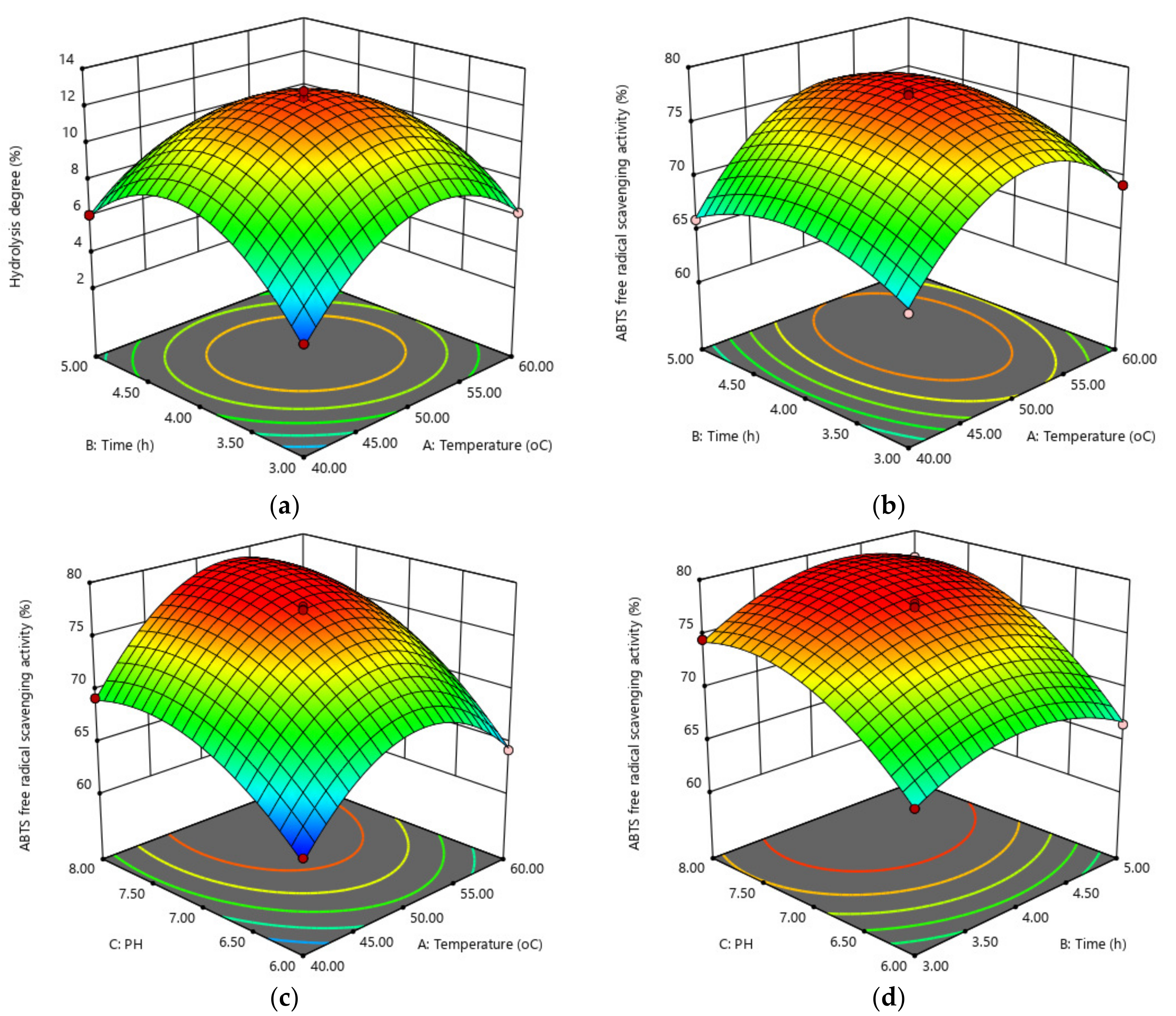
2.2. Optimum Hydrolysis Conditions
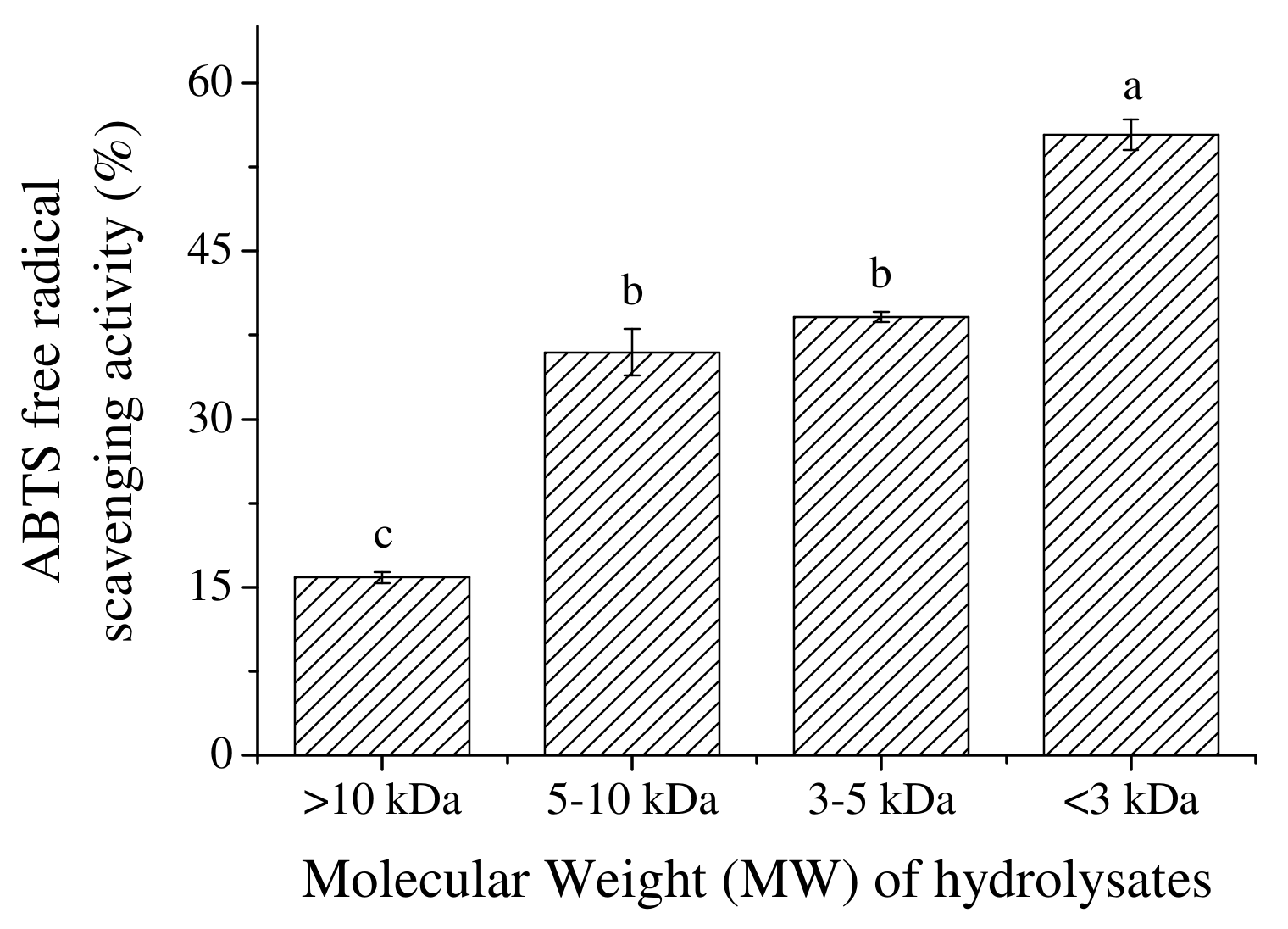
2.3. The Effect of MW
2.4. Identification of Peptide Sequences by LC–MS/MS
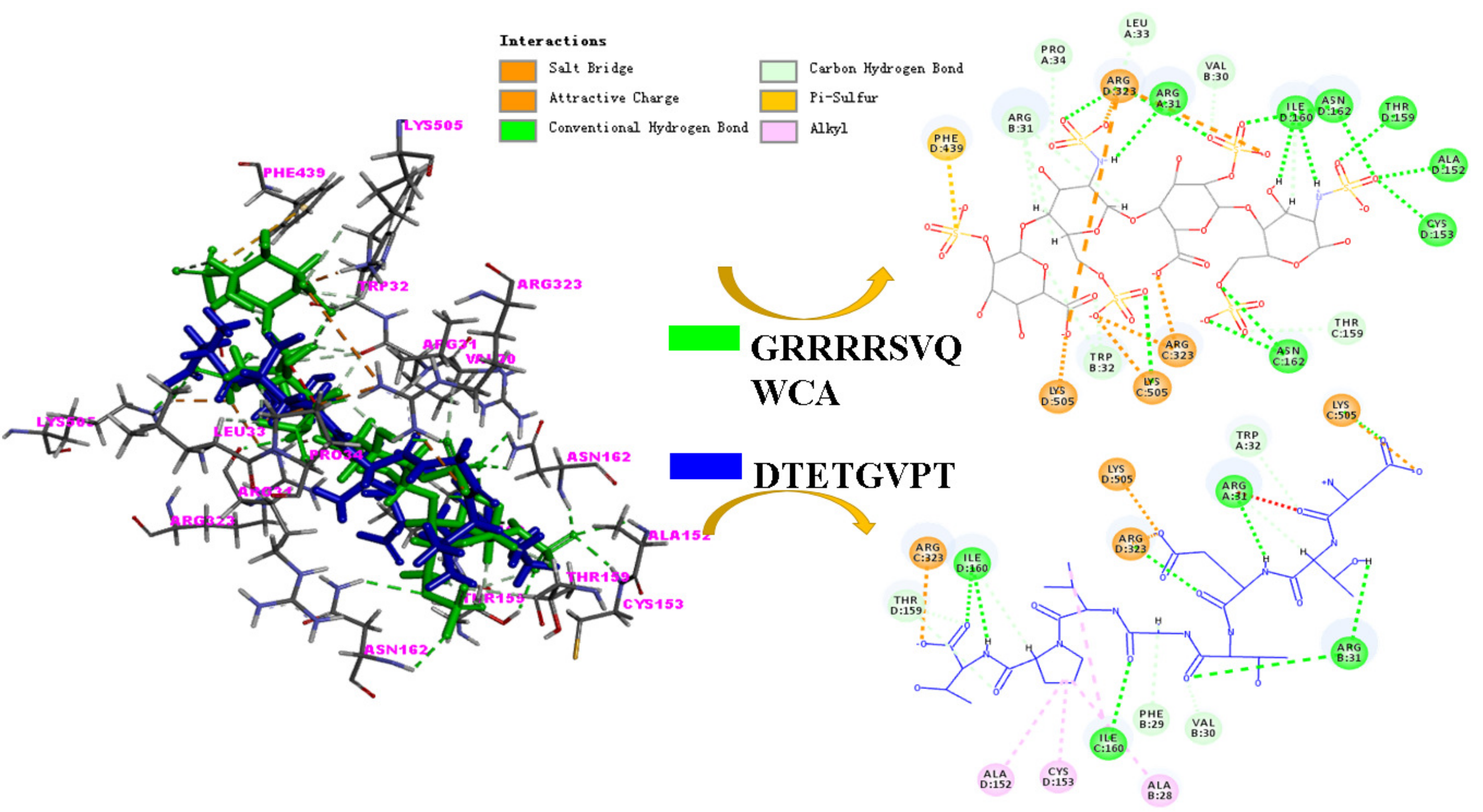
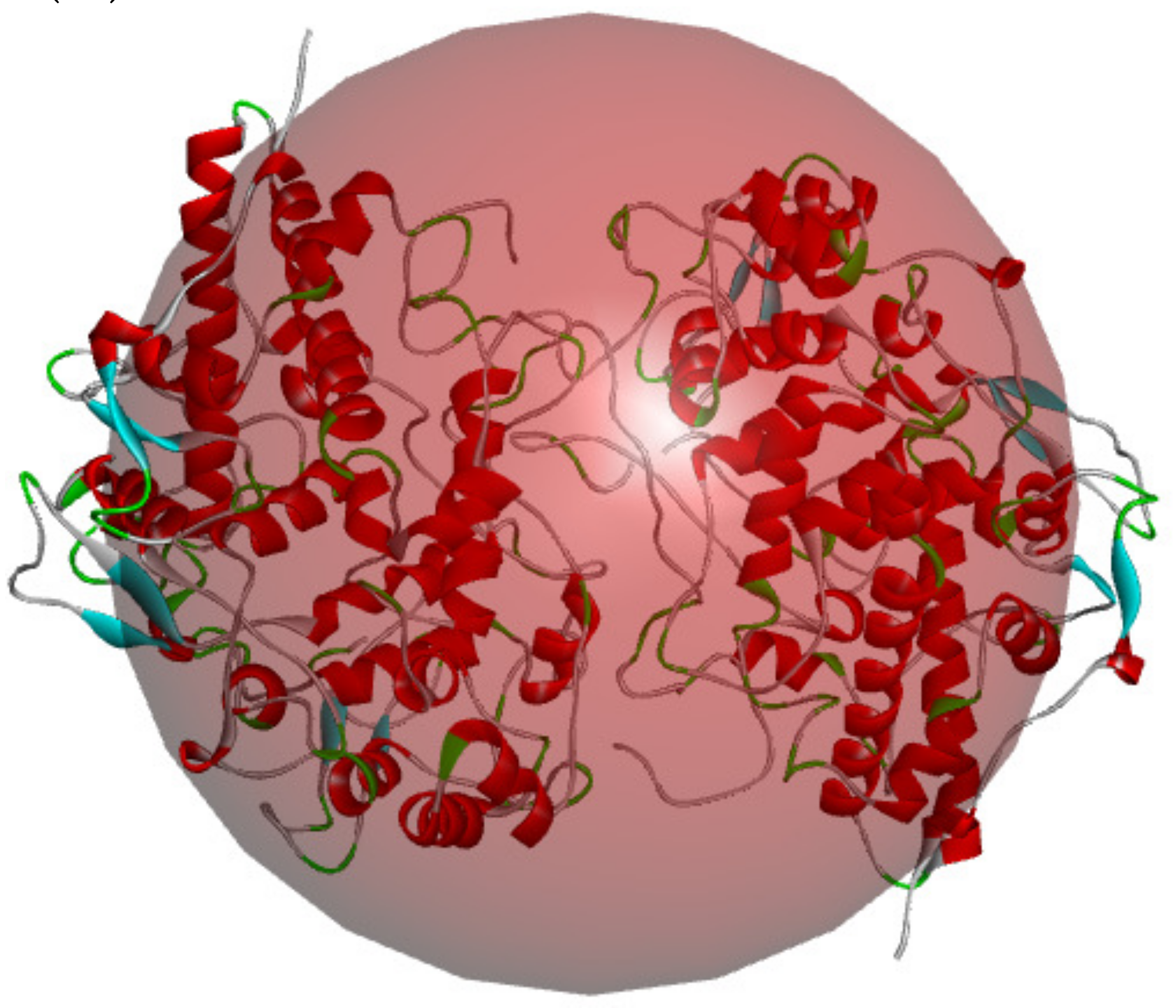
2.5. Molecular Mechanism
3. Materials and Methods
3.1. Materials and Reagents
3.2. Proteolytic Hydrolysis of False Abalone
3.3. Determination of Protein Concentration and Hydrolysis Degree
3.4. Antioxidant Activity Determination of Hydrolysates Using ABTS+• Assay
3.5. Selection of Proteolytic Enzyme
3.6. Response Surface Methodology
3.7. Fractionation of the Optimized Hydrolysates
3.8. LC–MS/MS Assay
3.9. Molecular Docking
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sun, M.L.; Zhang, Q.; Ma, Q.; Fu, Y.-H.; Jin, W.G.; Zhu, B.-W. Affinity purification of angiotensin-converting enzyme inhibitory peptides from Volutharpa ampullacea perryi protein hydrolysate using Zn-SBA-15 immobilized ACE. Eur. Food Res. Technol. 2018, 244, 457–468. [Google Scholar] [CrossRef]
- He, S.; Sun, X.; Du, M.; Chen, H.; Tan, M.; Sun, H.; Zhu, B. Effects of muscle protein denaturation and water distribution on the quality of false abalone (Volutharpa ampullacea perryi) during wet heating. J. Food Process Eng. 2018, e12932. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, J.; Li, D.; Wen, C.; Liu, H.; Song, S.; Zhu, B. Comparison of polysaccharides of Haliotis discus hannai and Volutharpa ampullacea perryi by PMP-HPLC-MSn analysis upon acid hydrolysis. Carbohyd. Res. 2015, 415, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Gómez-Guillén, M.; Giménez, B.; López-Caballero, M.A.; Montero, M. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocolloid. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Kim, S.K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Malle, E.; Furtmüller, P.; Sattler, W.; Obinger, C. Myeloperoxidase: A target for new drug development? Br. J. Pharmacol. 2007, 152, 838–854. [Google Scholar]
- Guerra-Vargas, M.A.; Rosales-Hernández, M.C.; Martínez-Fonseca, N.; Padilla-Martínez, I.; Fonseca-Sabater, Y.; Martínez-Ramos, F. 2-Acetyl-4-aminoresorcinol derivatives: synthesis, antioxidant activity and molecular docking studies. Med. Chem. Res. 2018, 27, 1186–1197. [Google Scholar] [CrossRef]
- Van der Does, A.M.; Hensbergen, P.J.; Bogaards, S.J.; Cansoy, M.; Deelder, A.M.; van Leeuwen, H.C.; Drijfhout, J.W.; van Dissel, J.T.; Nibbering, P.H. The human lactoferrin-derived peptide hLF1-11 exerts immunomodulatory effects by specific inhibition of myeloperoxidase activity. J. Immunol. 2012, 1102777. [Google Scholar] [CrossRef]
- Tavano, O.L. Protein hydrolysis using proteases: An important tool for food biotechnology. J. Mol. Catal. B: Enzym. 2013, 90, 1–11. [Google Scholar] [CrossRef]
- Hou, H.; Li, B.; Zhao, X.; Zhang, Z.; Li, P. Optimization of enzymatic hydrolysis of Alaska pollock frame for preparing protein hydrolysates with low-bitterness. LWT-Food Sci. Technol. 2011, 44, 421–428. [Google Scholar] [CrossRef]
- Mao, X.Y.; Cheng, X.; Wang, X.; Wu, S.J. Free-radical-scavenging and anti-inflammatory effect of yak milk casein before and after enzymatic hydrolysis. Food Chem. 2011, 126, 484–490. [Google Scholar] [CrossRef]
- Kong, B.; Xiong, Y.L. Antioxidant activity of zein hydrolysates in a liposome system and the possible mode of action. J. Agric. Food Chem. 2006, 54, 6059–6068. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chi, Y.J.; Zhao, M.Y.; Xu, W. Influence of degree of hydrolysis on functional properties, antioxidant and ACE inhibitory activities of egg white protein hydrolysate. Food Sci. Biotechnol. 2012, 21, 27–34. [Google Scholar] [CrossRef]
- Ovissipour, M.; Kenari, A.A.; Motamedzadegan, A.; Nazari, R.M. Optimization of enzymatic hydrolysis of visceral waste proteins of yellowfin tuna (Thunnus albacares). Food Bioprocess Tech. 2012, 5, 696–705. [Google Scholar] [CrossRef]
- Cheison, S.C.; Schmitt, M.; Leeb, E.; Letzel, T.; Kulozik, U. Influence of temperature and degree of hydrolysis on the peptide composition of trypsin hydrolysates of β-lactoglobulin: Analysis by LC–ESI-TOF/MS. Food Chem. 2010, 121, 457–467. [Google Scholar] [CrossRef]
- You, L.; Regenstein, J.M.; Liu, R.H. Optimization of hydrolysis conditions for the production of antioxidant peptides from fish gelatin using response surface methodology. J. Food Sci. 2010, 75, C582–C587. [Google Scholar] [CrossRef] [PubMed]
- Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of peptide size on antioxidant properties of African yam bean seed (Sphenostylis stenocarpa) protein hydrolysate fractions. Int. J. Mol. Sci. 2011, 12, 6685–6702. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.Y.; Lee, J.H.; Samarakoon, K.; Kim, J.S.; Jeon, Y.J. Purification and determination of two novel antioxidant peptides from flounder fish (Paralichthys olivaceus) using digestive proteases. Food Chem. Toxicol. 2013, 52, 113–120. [Google Scholar] [CrossRef]
- Chen, M.; Li, B. The effect of molecular weights on the survivability of casein-derived antioxidant peptides after the simulated gastrointestinal digestion. Innov. Food Sci. Emerg. 2012, 16, 341–348. [Google Scholar] [CrossRef]
- Wang, B.; Li, L.; Chi, C.F.; Ma, J.H.; Luo, H.Y.; Xu, Y.f. Purification and characterisation of a novel antioxidant peptide derived from blue mussel (Mytilus edulis) protein hydrolysate. Food Chem. 2013, 138, 1713–1719. [Google Scholar] [CrossRef] [PubMed]
- Najafian, L.; Babji, A.S. Production of bioactive peptides using enzymatic hydrolysis and identification antioxidative peptides from patin (Pangasius sutchi) sarcoplasmic protein hydolysate. J. Funct. Foods 2014, 9, 280–289. [Google Scholar] [CrossRef]
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Kouzuma, Y.; Yonekura, M. Structures and properties of antioxidative peptides derived from royal jelly protein. Food Chem. 2009, 113, 238–245. [Google Scholar] [CrossRef]
- Memarpoor-Yazdi, M.; Asoodeh, A.; Chamani, J. A novel antioxidant and antimicrobial peptide from hen egg white lysozyme hydrolysates. J. Funct. Foods 2012, 4, 278–286. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Kim, S.K. Antioxidant properties of a radical-scavenging peptide purified from enzymatically prepared fish skin gelatin hydrolysate. J. Agric. Food Chem. 2005, 53, 581–587. [Google Scholar] [CrossRef]
- Chen, H.M.; Muramoto, K.; Yamauchi, F. Structural analysis of antioxidative peptides from Soybean. beta.-Conglycinin. J. Agric. Food Chem. 1995, 43, 574–578. [Google Scholar] [CrossRef]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef]
- Shazly, A.B.; He, Z.; El-Aziz, M.A.; Zeng, M.; Zhang, S.; Qin, F.; Chen, J. Fractionation and identification of novel antioxidant peptides from buffalo and bovine casein hydrolysates. Food Chem. 2017, 232, 753–762. [Google Scholar] [CrossRef]
- Zhuang, H.; Tang, N.; Yuan, Y. Purification and identification of antioxidant peptides from corn gluten meal. J. Funct. Foods 2013, 5, 1810–1821. [Google Scholar] [CrossRef]
- Taheri, A.; Farvin, K.S.; Jacobsen, C.; Baron, C.P. Antioxidant activities and functional properties of protein and peptide fractions isolated from salted herring brine. Food Chem. 2014, 142, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Hunter, H.N.; Demcoe, A.R.; Jenssen, H.; Gutteberg, T.J.; Vogel, H.J. Human lactoferricin is partially folded in aqueous solution and is better stabilized in a membrane mimetic solvent. Antimicrob. Agents Ch. 2005, 49, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Battistuzzi, G.; Bellei, M.; Zederbauer, M.; Furtmüller, P.G.; Sola, M.; Obinger, C. Redox thermodynamics of the Fe (III)/Fe (II) couple of human myeloperoxidase in its high-spin and low-spin forms. Biochemistry 2006, 45, 12750–12755. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, T.J.; Davey, C.A.; Fenna, R.E. X-ray crystal structure and characterization of halide-binding sites of human myeloperoxidase at 1.8 Å resolution. J. Biol. Chem. 2000, 275, 11964–11971. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Wong, C.C.; Cheng, K.W.; Chen, F. Antioxidant properties in vitro and total phenolic contents in methanol extracts from medicinal plants. LWT Food Sci. Technol. 2008, 41, 385–390. [Google Scholar] [CrossRef]
- Carpena, X.; Vidossich, P.; Schroettner, K.; Callisto, B.M.; Banerjee, S.; Stampler, J.; Soudi, M.; Furtmüller, PG.; Rovira, C.; Fita, I. Essential role of proximal histidine-asparagine interaction in mammalian peroxidases. J. Biol. Chem. 2009, 284, 25929–25937. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Independent Variables | Responses | |||
|---|---|---|---|---|---|
| A: Temperature (oC) | B: Time (h) | C: pH | Response 1: Hydrolysis Degree (%) | Response 2: ABTS+• Scavenging Activity (%) | |
| 1 | 50 | 4 | 7 | 12.44 | 77.56 |
| 2 | 50 | 4 | 7 | 12.81 | 77.49 |
| 3 | 40 | 5 | 7 | 6.11 | 65.99 |
| 4 | 50 | 5 | 8 | 10.09 | 77.34 |
| 5 | 50 | 3 | 8 | 9.22 | 74.53 |
| 6 | 50 | 4 | 7 | 12.23 | 77.28 |
| 7 | 60 | 3 | 7 | 6.24 | 69.25 |
| 8 | 50 | 3 | 6 | 3.33 | 66.37 |
| 9 | 40 | 4 | 6 | 4.03 | 62.15 |
| 10 | 60 | 5 | 7 | 5.37 | 72.39 |
| 11 | 60 | 4 | 8 | 10.16 | 75.23 |
| 12 | 50 | 5 | 6 | 4.25 | 66.57 |
| 13 | 50 | 4 | 7 | 12.35 | 77.85 |
| 14 | 60 | 4 | 6 | 5.06 | 64.25 |
| 15 | 50 | 4 | 7 | 12.35 | 77.26 |
| 16 | 40 | 3 | 7 | 3.88 | 65.14 |
| 17 | 40 | 4 | 8 | 8.67 | 69.25 |
| Source | Response 1 | Response 2 | ||||
|---|---|---|---|---|---|---|
| Mean Square | F Value | p Value | Mean Square | F Value | p Value | |
| Model | 22.13 | 189.74 | <0.0001 | 56.43 | 334.76 | <0.0001 |
| A | 2.14 | 18.37 | 0.0036 | 43.20 | 256.27 | <0.0001 |
| B | 1.24 | 10.63 | 0.0138 | 6.13 | 36.34 | 0.0005 |
| C | 57.62 | 493.99 | <0.0001 | 171.22 | 1015.74 | <0.0001 |
| AB | 2.40 | 20.60 | 0.0027 | 1.31 | 7.78 | 0.0270 |
| AC | 0.0529 | 0.4535 | 0.5223 | 3.76 | 22.33 | 0.0021 |
| BC | 0.0006 | 0.0054 | 0.9437 | 1.70 | 10.10 | 0.0155 |
| A2 | 48.37 | 414.66 | <0.0001 | 171.87 | 1019.61 | <0.0001 |
| B2 | 55.99 | 480.06 | <0.0001 | 35.57 | 211.01 | <0.0001 |
| C2 | 17.99 | 154.19 | <0.0001 | 48.07 | 285.20 | <0.0001 |
| Residual | 0.1166 | 0.1686 | ||||
| Lack of fit | 0.2065 | 4.19 | 0.1000 | 0.3162 | 5.46 | 0.0673 |
| Pure error | 0.0493 | 0.0579 | ||||
| Peptide no. | Peptide Sequence | Consensus Score | −CDOCKER Interaction Energy | Interactions with A Chain of MPO | Interactions with B Chain of MPO | Interactions with C Chain of MPO | Interactions with D Chain of MPO |
|---|---|---|---|---|---|---|---|
| hLF1-11 | GRRRRSVQWCA | - | 120.282 | ARG31, LEU33, PRO34 | VAL30, ARG31, TRP32 | THR159, ASN162, ARG323, LYS505 | ALA152, CYS153, ILE160, ASN162, PHE439, LYS505 |
| 39 | DTETGVPT | 3 | 121.516 | AGR31, TRP32 | ALA28, PHE29. VAL30, ARG31, TRP32 | ILE160, ARG323, LYS505 | ALA152, CYS153, THR159, ILE160, ARG323, LYS505 |
| 1 | NDNIQR | 2 | 96.9148 | ARG31, PRO34 | ARG31, TRP32, ALA35 | ILE160, LYS505 | ILE160, ASN162, ARG323, LYS505 |
| 2 | IELLLL | 2 | 112.572 | ARG31, TRP32, PRO34 | ALA28, ARG31, PRO34, ALA35 | ILE160, ASN162, PHE439 | ALA152, CYS153, THR159, ILE160, ASN162, ARG323 |
| 4 | LADEIR | 2 | 101.61 | ARG31, TRP32, PRO34 | VAL30, ARG31, TRP32 | ILE160, ASN162, LYS505 | ALA152, CYS153, ARG323 |
| 5 | LLKDQL | 2 | 106.002 | ALA28, VAL30, ARG31, PRO34 | ALA28, TRP32 | ILE160, ARG323, LYS505 | ARG323, LYS505 |
| 6 | LEILNT | 2 | 95.5421 | VAL30, ARG31 | ALA28, ARG31 | ALA152, CYS153, ILE160, ASN162, ARG323, LYS505 | THR159, ILE160, ASN162, ARG323 |
| 8 | QDPLNR | 2 | 105.976 | PHE29, ARG31, TRP32, PRO34, ALA35 | TRP32 | ILE160, ASN162 | ASP321, ARG323, LYS505 |
| 9 | QVQNVR | 2 | 94.6778 | ARG31, PRO34 | ARG31, TRP32, PRO34 | ILE160, ASN162, ARG323 | ILE160, ASN162, SER319, ARG323, LYS505 |
| 10 | GTELFR | 2 | 97.3112 | ARG31, TRP32, PRO34 | ARG31, TRP32, ALA35 | ASN162, ARG323, LYS505 | ILE160, ASN162, ARG323, LYS505 |
| 12 | ISAAELR | 2 | 99.0441 | ARG31, TRP32, ALA35 | ARG31 | ILE160, ARG323, LYS505 | ILE160, ASN162, ARG323 |
| 13 | FPSIVGR | 2 | 89.5673 | VAL30, ARG31 | ALA28, PHE29, VAL30, ARG31, PRO34, ALA35 | ILE160, ASN162, SER319, ARG323, LYS505 | ALA152, CYS153, ILE160 |
| 14 | LTGMAFR | 2 | 90.0454 | ARG31, LEU33 | ARG31, TRP32, PRO34 | ILE160, ARG323 | ILE160, ASN162, ARG323, LYS505 |
| 16 | FAPQLLT | 2 | 105.345 | ALA28, ARG31 | ALA28, PHE29, VAL30, ARG31, PRO34 | ILE160, ARG323 | ILE160, ASN162, ARG323 |
| 20 | LEVNLMT | 2 | 106.656 | PHE29, ARG31, TRP32, PRO34 | ALA28, ARG31, TRP32, LEU33 | ILE160, ASN162, ARG323, LYS505 | ALA152, ILE160, ASN162, ARG323 |
| 22 | HLQLAIR | 2 | 98.3054 | ALA28, ARG31, TRP32 | ARG31, TRP32 | THR159, ILE160, ARG323 | SER156, THR159, ILE160, ASP321, ARG323 |
| 23 | IAKEGFA | 2 | 92.2588 | PHE29, ARG31, LEU33, PRO34 | ARG31, TRP32 | THR159, ARG323, LYS505 | ILE160, ASN162 |
| 24 | NLNPTTK | 2 | 95.8118 | ARG31, TRP32, PRO34 | PHE29, ARG31, ALA35 | CYS153, ARG323 | ILE160, ASN162, ARG323 |
| 25 | VSHYSTK | 2 | 90.9066 | ARG31 | ALA28, ARG31, TRP32, ALA35 | ILE160, ASN162, ARG323 | ALA152, IEL160, ARG323 |
| 28 | GLLLHWS | 2 | 99.7525 | ALA28, VAL30, ARG31, TRP32 | ALA28, PHE29, ARG31, TRP32, PRO34 | ALA152, CYS153, ILE160, ARG323, LYS505 | ILE160, ASN162, ARG323, LYS505 |
| 29 | LQLISAG | 2 | 89.9116 | ALA28, ARG31, TRP32 | VAL30, ARG31 | CYS153, ILE160, ARG161, ASN162, ASP321, ARG323, LYS505 | ILE160, ASN162 |
| 30 | QCELNFK | 2 | 103.547 | ARG31, PRO34 | - | ILE160, ASN162, ARG323, LYS505 | ALA152, CYS153, ASN162, ARG323 |
| 31 | GIHETTY | 2 | 120.866 | VAL30, ARG31, PRO34 | ALA28, ARG31, TRP32, PRO34 | THR159, ILE160, ASN162, ARG323 | ALA152, ILE160, ARG323 |
| 45 | VASFSTHK | 2 | 97.4559 | ALA28, ARG31 | ALA28, VAL30, ARG31, TRP32 | ILE160 | ALA152, SER319, ARG323, LYS505 |
| 46 | AVAGLGIL | 2 | 94.3888 | ARG31, TRP32 | PHE29, ARG31, TRP32 | ALA152, CYS153, ILE160, ASN162, ARG323, LYS505 | CYS153, ILE160, ASN162, ARG323 |
| 48 | VGDEAQSK | 2 | 114.397 | ARG31 | ALA28, TRP32 | YHR159, ILE160, ARG323 | THR159, ILE160, ASN162, ARG323, LYS505 |
| 49 | SSSVVAAL | 2 | 95.2041 | ARG31 | ARG31, TRP32 | ASN162, ARG323, LYS505 | ALA152, CYS153, ASN162, ARG323 |
| 50 | FAGDDAPR | 2 | 113.395 | ARG31, TRP32, PRO34 | PHE29, VAL30, ARG31, TRP32, PRO34 | ILE160, ASN162, ARG323 | THR159, ILE160, ASN162, ARG323 |
| 55 | KTKDDMHL | 2 | 111.666 | ARG31, TRP32, PRO34, ALA35 | ARG31, TRP32, PRO34 | ILE160, ARG323, LYS505 | CYS153, THR159, ILE160, ARG323 |
| 58 | VIPELNGK | 2 | 108.917 | ALA28, ARG31 | ALA28, ARG31, ALA35 | ARG323, LYS505 | ILE160, ASN163, ARG323 |
| 62 | GFAGDDAPR | 2 | 128.195 | ALA28, ARG31, TRP32, ALA35 | ALA28, ARG31, TRP32 | ILE160, SER319, ARG323, LYS505 | ILE160, ASN162, ARG323 |
| 64 | VATVGPISV | 2 | 96.6785 | ALA28, ARG31, PRO34 | ALA28, VAL30, ARG31, TRP32, PRO34 | THR159, ILE160, ARG323 | ILE160, ASN162, ARG323, LYS505 |
| 68 | SSSVALHKH | 2 | 78.6093 | ARG31, TRP32, PRO34 | ARG31, TRP32 | PRO151, CYS153, THR159, ILE160, ARG161, ASN162, ARG323, LYS505 | THR159, ILE160, ARG323 |
| 71 | GGKNLDELE | 2 | 145.154 | ARG31, PRO34 | ALA28, PHE29, ARG31, TRP32 | ILE160, ARG323 | CYS153, SER156, THR159, ILE160, ASN162, ASP321, ARG323, LYS505 |
| 111 | VGSSFVGGFG | 2 | 103.741 | VAL30, ARG31, TRP32 | ALA28, ARG31 | ILE160, ASN162, LYS505 | PRO151, CYS153, SER156, THR159, ILE160, ARG161, ASN162, ARG323 |
| 112 | DVNSLKSALA | 2 | 103.755 | ARG31, TRP32 | VAL30, ARG31, TRP32 | CYS153, ILE160, ARG323, LYS505 | ALA152, CYS153, ILE160, ARG161, ASN162 |
| 118 | IGGIGTVPVGR | 2 | 106.314 | ARG31, PRO34, SER42 | VAL30, ARG31, TRP32, PRO34 | PRO151, ALA152, CYS153, SER156, ILE160, ASN162, ASN317, SER319 | ARG323, ARG438, PHE439 |
| 125 | AGLFVSSFFSV | 2 | 116.238 | ALA28, ARG31, TRP32 | ALA28, VAL30, ARG31, PHE41 | CYS153, LYS308, ASP321, ARG323, PHE439, LYS505 | CYS153, SER156, THR159, ILE160, ARG323 |
| 148 | LTASGPSIGARP | 2 | 122.053 | ARG31 | ALA28, TRP32, ASP39 | ARG323, LYS505, PHE439 | PRO123, ALA152, CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162 |
| 182 | ILVGAAVCFFCLILA | 2 | 116.4 | ALA28, PRO103 | ARG31, TRP32 | PRO151, CYS153, PRO154, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ARG323 | ILE160, ASN162, LYS308, ASP321, ARG323, PHE439, LYS505 |
| 183 | STAGDTHLGGEDFDNR | 2 | 140.863 | ALA28, TRP32 | ARG31, PRG41, SER42, LEU43 | LYS308, ASP321, ARG323, PHE439, CYS440, CYS497, CYS505, LYS505 | PRO151, CYS153, GLY155, THR159, ILE160, ARG161, ASN162, ARG323 |
| 7 | NWDLVG | 1 | 99.433 | PHE29, VAL30, ARG31 | ALA28, PHE29, VAL30, ARG31, TRP32 | ARG323 | ARG323 |
| 11 | LLVKLL | 1 | 86.1708 | ALA28, VAL30, ARG31, TRP32, PRO34 | PHE29, VAL30, ARG31, PRO34 | ALA152, CYS153, ASN162, ARG323, LYS505 | ILE160, ASN162 |
| 15 | FPGIADR | 1 | 88.1065 | VAL30, ARG31 | PHE29, VAL30, ARG31, TRP32, | ASN162, ARG323 | ILE160, SER319, ARG323 |
| 17 | IIAPPER | 1 | 96.3472 | VAL30, ARG31, TRP32, PRO34 | ALA28, ARG31, TRP32 | ALA152, CYS153, IEL160, ASN162, ARG323, LYS505 | ILE160, ARG323, LYS505 |
| 21 | LIILELL | 1 | 98.6814 | ALA28, ARG31, TRP32, LEU33, PRO34 | ALA28, VAL30, ARG31, PRO34 | ILE160, ARG323, LYS505 | ALA152, CYS153, ILE160, ASN162 |
| 26 | NLLNIPK | 1 | 86.0812 | VAL30, ARG31, PRO34 | ARG31, TRP32, PRO34 | ILE160, ARG323 | THR159, ILE160 |
| 27 | SFRENNT | 1 | 102.146 | ARG31, PRO34 | PHE29, ARG31, TRP32 | ILE160, ASN162, LYS505 | ASN162, ASP321, ARG323, LYS505 |
| 32 | TEAPLNPK | 1 | 101.636 | VAL30, ARG31, TRP32, PRO34 | ALA28, ARG31, PRO34 | THR159, ILE160, ASN162, LYS505 | ILE160, ASN162, ARG323 |
| 34 | IILLLLV | 1 | 89.3092 | ALA28, VAL30, ARG31, TRP32, PRO34, ALA35 | VAL30, TRP32, PRO34 | ILE160, ASN162, ARG323, LYS505 | ILE160 |
| 35 | SFTTTAER | 1 | 111.511 | ARG31, TRP32 | PHE29, VAL30, ARG31, TRP32 | THR159, LYS505 | CYS153, SER156, THR159, ASN162 |
| 37 | TLEEEKLQ | 1 | 127.638 | ARG31, TRP32, PRO34 | ALA28, TRP32, LEU33, PRO34 | ILE160, ASN162, ARG323, LYS505 | ALA152, CYS153, THR159, ILE160, ARG323 |
| 40 | FLNNNALT | 1 | 88.3879 | ARG31, TRP32 | ARG31 | THR159, ARG323 | ILE160, ASN162, SER319, VAL320, ARG323 |
| 41 | LEVLGVPA | 1 | 96.8835 | PHE29, VAL30, ARG31, TRP32, LEU33, PRO34 | ALA28, VAL30, ARG31, PRO34 | CYS153, THR159, ILE160, ASN162, ARG323, LYS505 | ILE160 |
| 42 | KLDEKIVQ | 1 | 102.044 | VAL30, ARG31, LEU33, PRO34 | ALA28, ARG31, PRO34, SER42 | ARG323 | ALA152, CYS153, ILE160, ARG161, ASN162, SER319, ARG323, LYS505 |
| 44 | TVPIYEGY | 1 | 88.1488 | VAL30, TRP32, SER42 | ALA28, ARG31, TRP32 | ALA152, CYS153, SER156, THR159, ILE160, ARG161, ASN162 | ASN162 |
| 52 | SVKNTAGL | 1 | 89.225 | ARG31, LEU33 | ALA28, ARG31 | CYS153, ASN162, ARG323 | ILE160, ARG323, LYS505 |
| 53 | ADINAADQ | 1 | 121.529 | ARG31 | ARG31, TRP32, LEU33, PRO34 | ALA152, CYS153, ILE160, ASN162 | THR159, ILE160, ASN162, ARG323, LYS505 |
| 54 | KTWVKELQ | 1 | 80.8878 | ALA28, ARG31, PRO34, SER42 | ARG31, TRP32, | CYS153, THR159, ILE160, ASN162 | ILE160, ARG323, LYS505 |
| 57 | EEQVAAIR | 1 | 124.143 | ARG31, TRP32 | PHE29, ARG31, PRO34, ALA35 | THR159, ILE160, ARG323, PHE439 | ALA152, CYS153, THR159, ILE160, SER319, ARG323, LYS505 |
| 63 | MGSTLIMLL | 1 | 120.627 | VAL30, ARG31, TRP32, LEU33, PRO34 | ALA28, VAL30, ARG31 | ILE160, ASN162, PHE439, LYS505 | PRO151, CYS153, SER156, THR159, ILE160, ANS162 |
| 70 | LGKTVPDDV | 1 | 88.3103 | ARG31, TRP32, LEU33, PRO34 | PHE29, ARG31, PRO34 | ILE160, ASN162, ARG323, LYS505 | PRO151, ALA152, CYS153, ILE160, ARG161, ANS162, ARG323 |
| 73 | QVITIGNER | 1 | 113.155 | ALA28, VAL30, ARG31, PRO34 | ALA28, VAL30, ARG31, LEU33, ALA35 | ARG323, LYS505 | ALA152, PRO154, GLY155, SER156, THR159, ILE160, ANS162, SER319, ARG323, LYS505 |
| 74 | LSDLSPFPG | 1 | 82.875 | TRP32 | ALA28, PHE29, ARG31, TRP32, PRO34 | ALA152, CYS153, THR159, ILE160, ASN162, ARG323, LYS505 | ALA152, CYS153, THR159, ILE160, ARG161, ANS162, PHE439 |
| 78 | VGYDALTDQ | 1 | 133.602 | ALA28, VAL30, ARG31, TRP32 | ARG31, PRO34, ALA35 | ALA152, CYS153, ARG323, LYS505 | PRO151, CYS153, SER156, THR159, ILE160, ARG161, ANS162, ARG323, LYS505 |
| 81 | IEAIDQVGS | 1 | 124.082 | ARG31, LEU33 | VAL30, ARG31, SER42 | CYS153, THR159, ASN162, ARG323, PHE439 | CYS153, GLY155, SER156, THR159, ILE160, ASN162, ARG323, LYS505 |
| 82 | MKNPKASVL | 1 | 48.301 | ARG31, TRP32 | - | ILE160, LYS505 | SER156, THR159, ILE160, ASN162, ARG323, PHE439 |
| 84 | LIIIIAAMT | 1 | 80.6012 | VAL30, ARG31, TRP32 | ALA28, ARG31, PRO34, ALA35 | ILE160, ASN162 | CYS153, ILE160, ASN162 |
| 90 | AGFAGDDAPR | 1 | 118.225 | ARG31, PRO34 | ALA28, VAL30, ARG31, PRO34 | ARG323 | CYS153, SER156, THR159, ILE160, ASN162, ARG323 |
| 91 | DAVTYTEHAK | 1 | 130.855 | VAL30, ARG31, PRO34 | ARG31, PHE41 | ILE160, ARG323, LYS505 | CYS153, PRO154, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ARG323, PHE439 |
| 94 | VLMILPSVTG | 1 | 80.7513 | VAL30, ARG31, PRO34, ALA35 | ALA28, ARG31, TRP32, LEU33, PRO34 | ILE160, ASN162, ARG323 | PRO151, ALA152, CYS153, GLY155, THR159, ILE160, ARG161, ARG323, PHE439 |
| 107 | DSGLLTPESV | 1 | 88.9027 | ALA28, ARG31, SER42 | ARG31, TRP32, PRO34 | CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ARG323, LYS505 | ILE160 |
| 109 | VEQEILETGI | 1 | 95.3997 | ARG31, TRP32 | VAL30, ARG31, TRP32, PHE41 | THR159, ASN162, ARG323, LYS505 | ALA152, PRO154, GLY155, SER156, THR159, ILE160, ARG161, ANS162, ARG323, LYS505 |
| 113 | SDDLDLGQVG | 1 | 63.6376 | ARG31, PRO34 | VAL30, ARG31, TRP32, PRO34 | ARG323, LYS505 | ARG148, CYS153, GLY155, SER156, THR159, ILE160, ARG161, ANS162 |
| 114 | HQGVMVGMGQK | 1 | 108.89 | ALA28, VAL30, ARG31, TPR32 | ARG31, ASP39, SER42 | THR159, PHE439, LYS505 | PRO151, ALA152, CYS153, THR159, ILE160, ARG161, ANS162 |
| 122 | LSAAGLEAGNV | 1 | 104.596 | ARG31, TRP32 | ARG31, SER42 | THR159, ARG323 | ALA152, CYS153, PRO154, GLY155, SER156, THR159, ILE160, ANS162, SER319, ASP321, ARG323 |
| 123 | ATAASSSSLEK | 1 | 107.999 | ARG31, TRP32, PRO34 | VAL30, ARG31, ASP39, SER42 | ARG323, PHE439, LYS505 | CYS153, GLY155, THR159, ILE160, ARG161, ASN162 |
| 124 | VVVYGGVAVNH | 1 | 58.4842 | ARG31 | ALA28, ARG31, PRO34, PHE41, SER42 | ASN162, ARG323, PHE439 | ARG148, PRO151, ALA152, CYS153, SER156, THR159, ILE160, ANS162, ARG323 |
| 130 | KAGSERNVLIF | 1 | 127.874 | ARG31, TRP32, PRO34, ALA35, ASP39, SER42 | ARG31, PRO34 | CYS153, PRO154, GLY155, SER156, SER156, THR159, ILE160, ARG161, ASN162, PHE439, LYS505 | ILE160, ARG161, ARG323, LYS505 |
| 137 | IGLFGGAGVGK | 1 | 104.383 | ALA28, ARG31, PRO34 | PHE29, ARG31, PRO34, PHE41 | ARG323, PHE439, LYS505 | PRO151, CYS153, SER156, THR159, ILE160, ARG323 |
| 139 | TLSIQNDQASQR | 1 | 116.278 | ARG31, TRP32 | - | ILE160, ARG161, ASN162, ARG323, PHE439, CYS497, LYS505 | ARG148, CYS153, ARG161, ASN162, SER156, THR159, ILE160, ARG323 |
| 142 | DSYVGDEAQSKR | 1 | 91.8078 | PRO34 | ARG31 | ARG323, PHE439 | CYS153, GLY155, SER156, THR159, ILE160, ARG161, SER319, ARG323, ARG504 |
| 152 | YISHIELAFSSV | 1 | 122.83 | ARG31, TRP32, PRO34 | ALA28, TRP32 | ARG323, PHE439, LYS505 | PRO151, CYS153, PRO154, GLY155, SER156, THR159, ILE160, ASN162 |
| 154 | EGGGIVESIGEG | 1 | 150.197 | ARG31, TRP32, PRO34 | ARG31, TRP32, SER42 | ALA152, ARG323, LYS505 | ALA152, CYS153, SER156, ILE160, ARG161, ARG323, PHE439 |
| 155 | CRPGALESGPAL | 1 | 91.9951 | ALA28, PRO34 | ARG31 | ALA152, CYS153, PRO154, GLY155, SER156, ILE160, ARG161, ASN162, ARG323 | ILE160, ARG323, PHE439, LYS505 |
| 156 | FNLWGLSCSSLL | 1 | 88.0987 | VAL30, ARG31, TRP32, PRO34 | VAL30, ARG31, PRO34, SER42, LEU43 | ASN162 | PRO123, ARG148, PRO151, CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ASP321, ARG323 |
| 161 | AKCGAYQGQVLIF | 1 | 114.346 | ARG31, TRP32 | TRP32 | ILE160, ASN162, LYS308, ARG323, PHE439 | CYS153, SER156, THR159, ILE160, ARG161, ASN162 |
| 169 | FTQAGSEVSALLGR | 1 | 130.617 | ARG31, PRO34 | VAL30, ARG31 | - | CYS153, GLY155, SER156, ILE160, ARG161, ASN162, ARG323, PHE439 |
| 170 | TTGIVLDSGDGVTH | 1 | 95.5298 | ARG31, PRO34 | VAL30, ARG31, PRO34, SER42 | ASN162, ARG323, ARG438, PHE439, GLY441 | PRO123, PRO151, CYS153, PRO154, GLY155, SER156, THR159, ILE160, ARG161, SER319, ARG323 |
| 172 | AHGGYSVFAGVGER | 1 | 106.868 | VAL30, ARG31 | PHE29, VAL30, ARG31, TRP32, SER42 | CYS153, ASN162, PHE439, LYS505 | ARG148, ALA152, CYS153, PRO154, GLY155, THR159, ARG161, ASN162, ARG323, LYS505 |
| 177 | ELPDGQVITIGNER | 1 | 113.61 | ARG31, TRP32, PRO34, ASP39, SER42 | ALA28, ARG31, TRP32, LEU33, PRO34 | CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162, SER319. ARG323, LYS505 | THR159, ASP321, ARG323 |
| 178 | SAYLVLTITIAAMT | 1 | 111.103 | ARG31, TRP32, PRO34 | ALA28, ARG31, PHE41 | CYS153, ILE160, ARG438, PHE439, LYS505 | ALA152, CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ARG323 |
| 180 | WEDLANEIQEELNK | 1 | 107.719 | ARG31, TRP32 | VAL30, ARG31, TRP32, PRO34, PHE41 | THR159, ARG438, CYS440, GLY441, LYS505 | PRO123, ARG148, PRO151, ALA152, CYS 153, THR159, ILE160, ARG161, ASN162, ARG323, LYS505 |
| 186 | EQEGGLGNFMNFMKENG | 1 | 163.68 | TRP32, PRO34 | ALA28, PHE29, ARG31, LEU33, PRO34, PHE41, PRO101, PRO103 | ILE160, ARG323, PHE439, CYS440 | PRO124, PRO151, CYS153, SER156, THR159, ILE160, ARG161, ASN162, SER319 |
| 51 | LILLIRAK | 0 | 74.004 | ALA28, PHE29, ARG31, TRP32, PRO34 | ALA28, ARG31, PRO34, SER42 | ILE160, PHE439, LYS505 | ALA152, CYS153, SER156, ILE160, ARG161, ANS162 |
| 59 | LARDKAAN | 0 | 94.6022 | ARG31, TRP32 | PHE29, VAL30, ARG31, LEU33 | THR159, ILE160, ASN162, ARG323, LYS505 | THR159 |
| 67 | VLFPLTTLQ | 0 | 72.9901 | VAL30 | ARG31, TRP32, SER42 | CYS153, LYS505 | PRO151, ALA152, CYS153, SER156, ARG161, ANS162, ARG323 |
| 72 | SMSFFGLIM | 0 | 86.9571 | - | ALA28, ARG31, TRP32, PRO34 | ASN162, ARG323, PHE439 | PRO151, ALA152, ILE160, ARG161, ASN162, ASP321, LYS505 |
| 85 | ISGLIYEETR | 0 | 75.3523 | ARG31, TRP32, SER42 | ALA28, ARG31, LEU33, PRO34 | CYS153, PRO154, GLY155, SER156, THR159, ILE160, ASN162, ARG323, LYS505 | CYS153, ARG323 |
| 99 | VVTSGNSLNG | 0 | 93.2669 | VAL30, ARG31, TRP32 | PHE29, ARG31, TRP32 | ILE160, ARG323, PHE439 | ALA152, CYS153, ILE160, ANS162, LYS505 |
| 117 | VEIIANDQGNR | 0 | 114.499 | ARG31, PRO34 | VAL30, ARG31, SER42 | ASN162, ARG323, PHE439 | ARG148, CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ARG323 |
| 121 | PTKVGINGFGR | 0 | 81.1789 | ALA28, PHE29, ARG31, TRP32 | ALA28, SER42 | LYS505 | CYS153, GLY155, SER156, THR159, ILE160 |
| 140 | ELISNASDALDK | 0 | 74.9956 | ARG31, TRP32, TRP47 | LEU33 | ARG148, PRO151, CYS153, PRO154, THR159, ILE160, ARG161, ASN162, ARG323, LYS505 | ARG323 |
| 162 | LLTLATCVGDGPA | 0 | 72.2681 | VAL30, ARG31, TRP32, SER42 | ARG31 | CYS153, GLY155, SER156, THR159, ILE160, ARG161, ASN162 | CYS153, LYS505 |
| 164 | RSLGDFACQLEHL | 0 | 75.728 | ARG31, SER42 | TRP32 | PRO151, CYS153, GLY155, SER156, THR159, ILE160, ARG161, LYS505 | THR159, LYS308, ARG323, PHE439, CYS440 |
| 171 | LISWYDNEYGYSNR | 0 | 137.019 | ALA28, ARG31, TRP32, PRO34 | PHE29, PHE41, SER42 | THR159, ASN162 | ALA152, CYS153, PRO154, SER156, THR159, ARG161, ASN162, VAL320, ASP321, ARG323 |
| 191 | LVQDVANNTNEEAGDGTTTATVLAR | 0 | 120.381 | VAL30, ARG31, TRP32, LEU33 | VAL30, TRP32, LEU43 | THR159, ILE160, PHE439 | CYS153, GLY155, SER156, ASN157, THR159, ILE160, ARG161, ASN317, SER319, ARG323, ARG504, LYS505 |
| 192 | AQTISYEVSMALVLLFPLFLGGSFSF | 0 | 185.828 | TRP32 | ARG31, TRP32, PRO34, SER42 | ASN162, ARG323, PHE439, GLY441, PRO443, LYS505 | GLN122, PRO123, CYS153, PRO154, GLY155, SER156, THR159, ILE160, ARG161, ASN162, ARG323, LYS505 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, S.; Zhang, Y.; Sun, H.; Du, M.; Qiu, J.; Tang, M.; Sun, X.; Zhu, B. Antioxidative Peptides from Proteolytic Hydrolysates of False Abalone (Volutharpa ampullacea perryi): Characterization, Identification, and Molecular Docking. Mar. Drugs 2019, 17, 116. https://doi.org/10.3390/md17020116
He S, Zhang Y, Sun H, Du M, Qiu J, Tang M, Sun X, Zhu B. Antioxidative Peptides from Proteolytic Hydrolysates of False Abalone (Volutharpa ampullacea perryi): Characterization, Identification, and Molecular Docking. Marine Drugs. 2019; 17(2):116. https://doi.org/10.3390/md17020116
Chicago/Turabian StyleHe, Shudong, Yi Zhang, Hanju Sun, Ming Du, Jianlei Qiu, Mingming Tang, Xianbao Sun, and Beiwei Zhu. 2019. "Antioxidative Peptides from Proteolytic Hydrolysates of False Abalone (Volutharpa ampullacea perryi): Characterization, Identification, and Molecular Docking" Marine Drugs 17, no. 2: 116. https://doi.org/10.3390/md17020116
APA StyleHe, S., Zhang, Y., Sun, H., Du, M., Qiu, J., Tang, M., Sun, X., & Zhu, B. (2019). Antioxidative Peptides from Proteolytic Hydrolysates of False Abalone (Volutharpa ampullacea perryi): Characterization, Identification, and Molecular Docking. Marine Drugs, 17(2), 116. https://doi.org/10.3390/md17020116