?-Carrageenan Oligosaccharides of Distinct Anti-Heparanase and Anticoagulant Activities Inhibit MDA-MB-231 Breast Cancer Cell Migration

 and
and
Abstract
:
1. Introduction
2. Results and Discussion
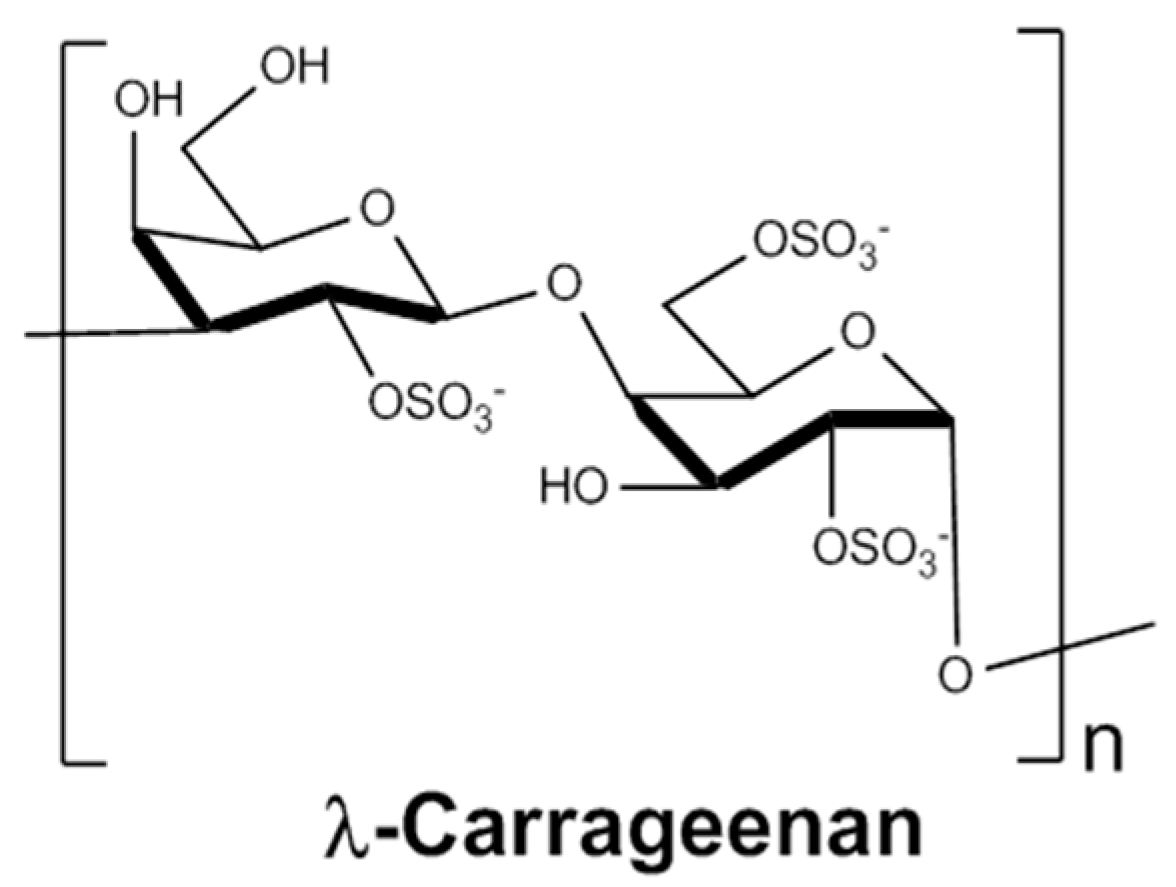
2.1. Depolymerisation of λ-Carrageenan
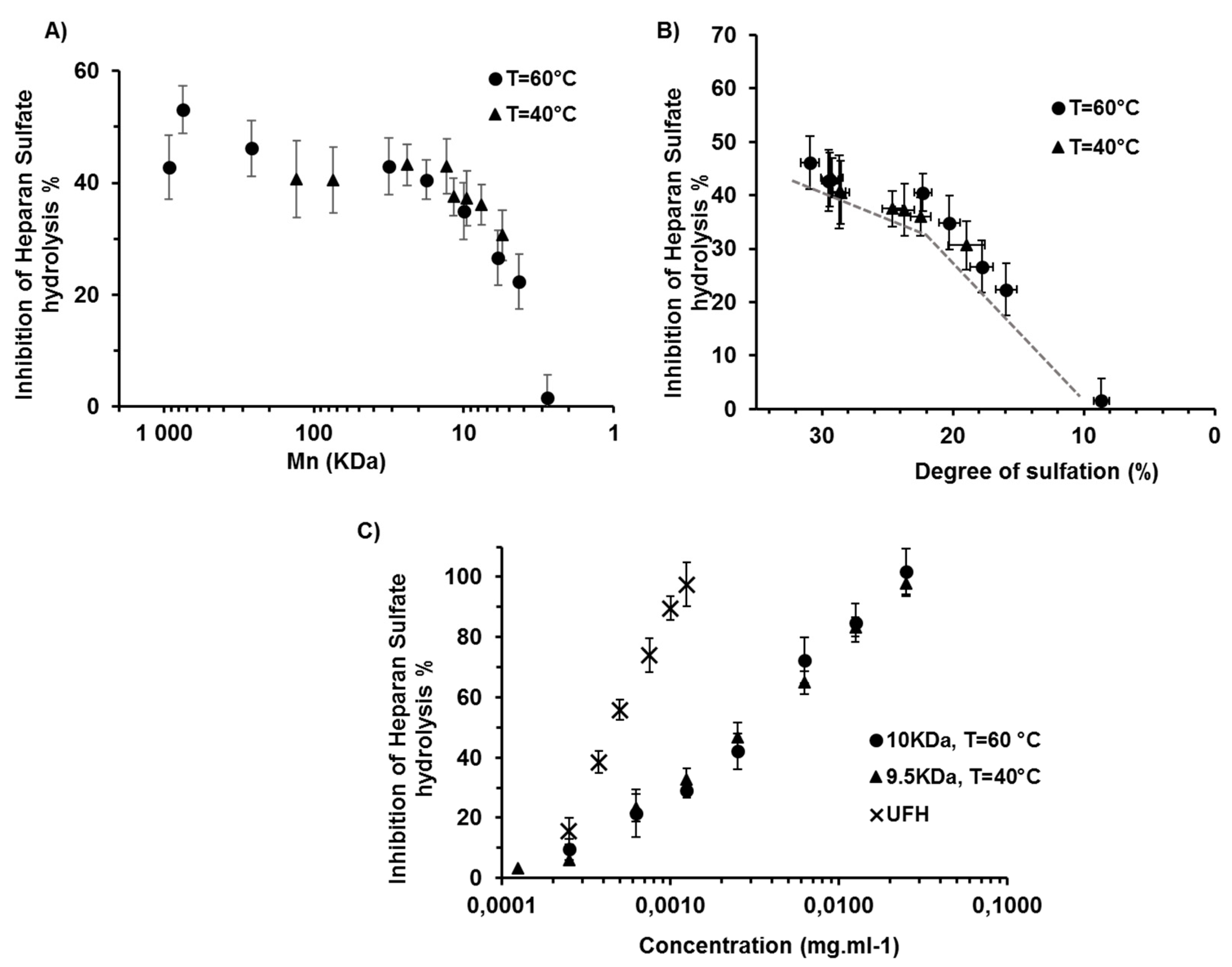
2.2. Effects of the Degree of Depolymerisation and Sulfation on the Anti-Heparanase Activity of λ-CO
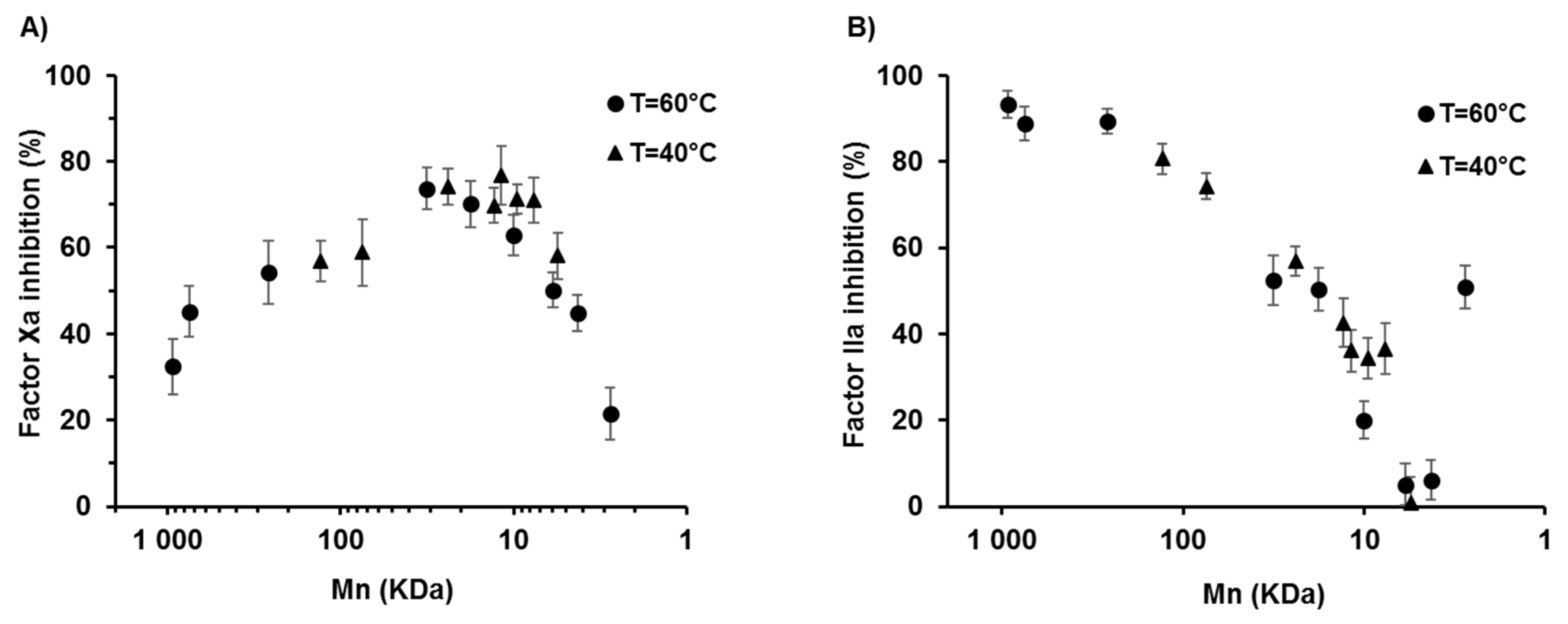
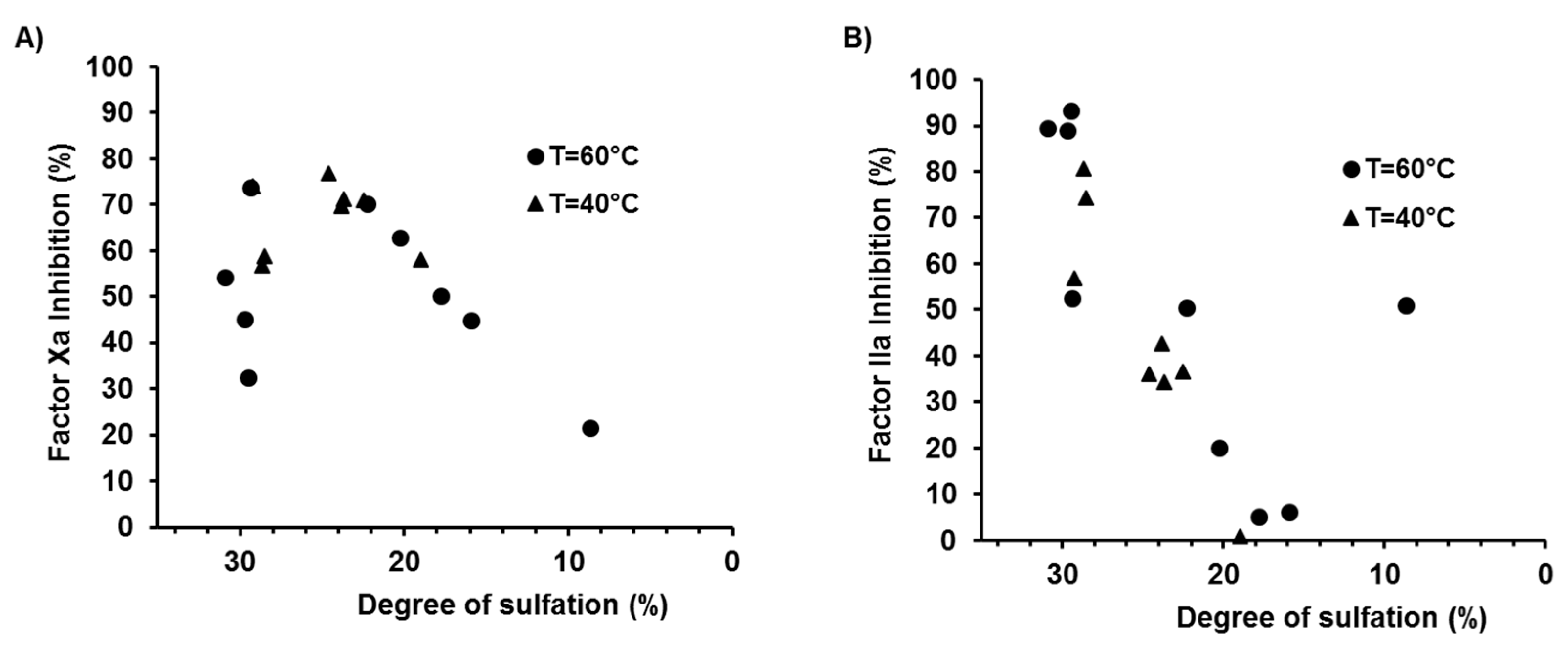
2.3. Effects of the Degree of Depolymerisation and Sulfation on the Anticoagulant Activity of λ-CO
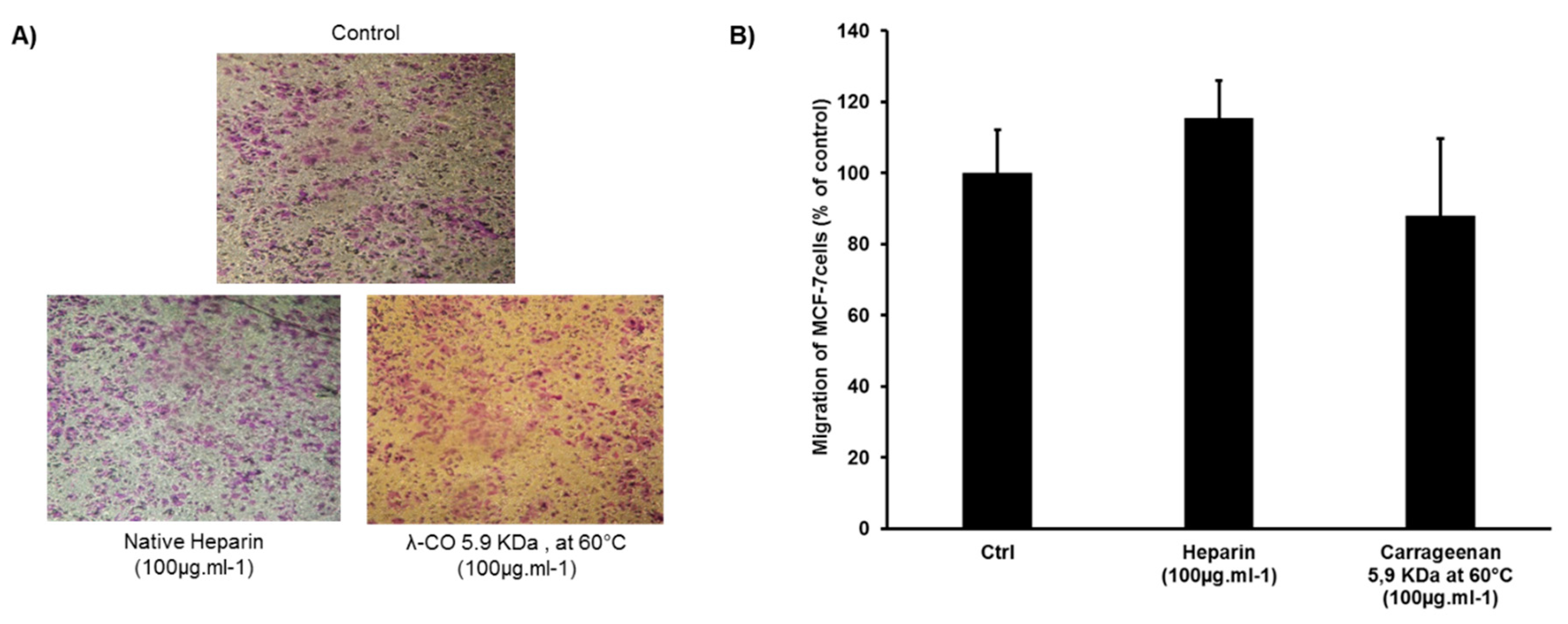
2.4. Effect of λ-CO Candidates on the Migration of MDA-MB-231 Breast Cancer Cells In Vitro
3. Materials and Methods
3.1. Depolymerisation of λ-Carrageenan for the Production of Oligosaccharides (λ-CO)
3.2. Structural and Quantitative Analysis of λ-CO by Size Exclusion Chromatography (SEC)
3.3. Quantification of the Sulfation Degree of λ-CO
3.4. Anti-Heparanase Activity of λ-CO
3.5. Anticoagulant Activity of λ-CO
3.6. MDA-MB-231 Cell Migration Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A




References
- Belli, C.; Trapani, D.; Viale, G.; D’Amico, P.; Duso, B.A.; Della Vigna, P.; Orsi, F.; Curigliano, G. Targeting the microenvironment in solid tumors. Cancer Treat. Rev. 2018, 65, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhuang, X.; Lin, L.; Yu, P.; Wang, Y.; Shi, Y.; Hu, G.; Sun, Y. New horizons in tumor microenvironment biology: Challenges and opportunities. BMC Med. 2015, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F.R.; Capasso, M.; Hagemann, T. The tumor microenvironment at a glance. J. Cell Sci. 2012, 125, 5591–5596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivara, S.; Milazzo, F.M.; Giannini, G. Heparanase: A rainbow pharmacological target associated to multiple pathologies including rare diseases. Future Med. Chem. 2016, 8, 647–680. [Google Scholar] [CrossRef] [PubMed]
- Vlodavsky, I.; Friedmann, Y. Molecular properties and involvement of heparanase in cancer metastasis and angiogenesis. J. Clin. Investig. 2001, 108, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Nadir, Y.; Brenner, B. Heparanase multiple effects in cancer. Thromb. Res. 2014, 133, S90–S94. [Google Scholar] [CrossRef]
- Masola, V.; Secchi, M.F.; Gambaro, G.; Onisto, M. Heparanase as a target in cancer therapy. Curr. Cancer Drug Targets 2014, 14, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Ma, S. Recent advances in the discovery of heparanase inhibitors as anti-cancer agents. Eur. J. Med. Chem. 2016, 121, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Vlodavsky, I.; Ilan, N.; Naggi, A.; Casu, B. Heparanase: Structure, biological functions, and inhibition by heparin-derived mimetics of heparan sulfate. Curr. Pharm. Des. 2007, 13, 2057–2073. [Google Scholar] [CrossRef] [PubMed]
- Warkentin, T.E.; Levine, M.N.; Hirsh, J.; Horsewood, P.; Roberts, R.S.; Gent, M.; Kelton, J.G. Heparin-Induced Thrombocytopenia in Patients Treated with Low-Molecular-Weight Heparin or Unfractionated Heparin. N. Engl. J. Med. 1995, 332, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Naggi, A.; Casu, B.; Perez, M.; Torri, G.; Cassinelli, G.; Penco, S.; Pisano, C.; Giannini, G.; Ishai-Michaeli, R.; Vlodavsky, I. Modulation of the Heparanase-inhibiting Activity of Heparin through Selective Desulfation, Graded N-Acetylation, and Glycol Splitting. J. Biol. Chem. 2005, 280, 12103–12113. [Google Scholar] [CrossRef] [PubMed]
- Arvatz, G.; Weissmann, M.; Ilan, N.; Vlodavsky, I. Heparanase and cancer progression: New directions, new promises. Hum. Vaccines Immunother. 2016, 12, 2253–2256. [Google Scholar] [CrossRef] [PubMed]
- Loka, R.S.; Yu, F.; Sletten, E.T.; Nguyen, H.M. Design, synthesis, and evaluation of heparan sulfate mimicking glycopolymers for inhibiting heparanase activity. Chem. Commun. 2017, 53, 9163–9166. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Viola, C.M.; Brzozowski, A.M.; Davies, G.J. Structural characterization of human heparanase reveals insights into substrate recognition. Nat. Struct. Mol. Biol. 2015, 22, 1016–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; El Hadri, A.; Richard, S.; Denis, F.; Holte, K.; Duffner, J.; Yu, F.; Galcheva-Gargova, Z.; Capila, I.; Schultes, B.; et al. Synthesis and Biological Evaluation of a Unique Heparin Mimetic Hexasaccharide for Structure–Activity Relationship Studies. J. Med. Chem. 2014, 57, 4511–4520. [Google Scholar] [CrossRef] [PubMed]
- Alekseeva, A.; Casu, B.; Cassinelli, G.; Guerrini, M.; Torri, G.; Naggi, A. Structural features of glycol-split low-molecular-weight heparins and their heparin lyase generated fragments. Anal. Bioanal. Chem. 2014, 406, 249–265. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Elli, S.; Naggi, A.; Guerrini, M.; Torri, G.; Petitou, M. Investigating Glycol-Split-Heparin-Derived Inhibitors of Heparanase: A Study of Synthetic Trisaccharides. Molecules 2016, 21, 1602. [Google Scholar] [CrossRef] [PubMed]
- Pala, D.; Rivara, S.; Mor, M.; Milazzo, F.M.; Roscilli, G.; Pavoni, E.; Giannini, G. Kinetic analysis and molecular modeling of the inhibition mechanism of roneparstat (SST0001) on human heparanase. Glycobiology 2016, 26, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Roy, S.; Cochran, E.; Zouaoui, R.; Chu, C.L.; Duffner, J.; Zhao, G.; Smith, S.; Galcheva-Gargova, Z.; Karlgren, J.; et al. M402, a Novel Heparan Sulfate Mimetic, Targets Multiple Pathways Implicated in Tumor Progression and Metastasis. PLoS ONE 2011, 6, e21106. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.-J.; Yuan, L.; Hu, J.; Yu, H.-H.; Li, T.; Lin, S.-F.; Tang, S.-B. Phosphomannopentaose sulfate (PI-88) suppresses angiogenesis by downregulating heparanase and vascular endothelial growth factor in an oxygen-induced retinal neovascularization animal model. Mol. Vis. 2012, 18, 1649–1657. [Google Scholar] [PubMed]
- Guerrini, M.; Shriver, Z.; Bisio, A.; Naggi, A.; Casu, B.; Sasisekharan, R.; Torri, G. The tainted heparin story: An update. Thromb. Haemost. 2009, 102, 907–911. [Google Scholar] [CrossRef] [PubMed]
- Szajek, A.Y.; Chess, E.; Johansen, K.; Gratzl, G.; Gray, E.; Keire, D.; Linhardt, R.J.; Liu, J.; Morris, T.; Mulloy, B.; et al. The US regulatory and pharmacopeia response to the global heparin contamination crisis. Nat. Biotechnol. 2016, 34, 625–630. [Google Scholar] [CrossRef] [PubMed]
- de Jesus Raposo, M.; de Morais, A.; de Morais, R. Marine Polysaccharides from Algae with Potential Biomedical Applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Necas, J.; Bartosikova, L. Carrageenan: A review. Veterinární Med. 2013, 58, 187–205. [Google Scholar] [CrossRef]
- Campo, V.L.; Kawano, D.F.; da Silva, D.B.; Carvalho, I. Carrageenans: Biological properties, chemical modifications and structural analysis—A review. Carbohydr. Polym. 2009, 77, 167–180. [Google Scholar] [CrossRef]
- Pangestuti, R.; Kim, S.-K. Biological Activities of Carrageenan. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 72, pp. 113–124. ISBN 978-0-12-800269-8. [Google Scholar]
- Zia, K.M.; Tabasum, S.; Nasif, M.; Sultan, N.; Aslam, N.; Noreen, A.; Zuber, M. A review on synthesis, properties and applications of natural polymer based carrageenan blends and composites. Int. J. Biol. Macromol. 2017, 96, 282–301. [Google Scholar] [CrossRef] [PubMed]
- Tobacman, J.K. Review of harmful gastrointestinal effects of carrageenan in animal experiments. Environ. Health Perspect. 2001, 109, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, B.; Wu, Y.; Liu, Y.; Gu, X.; Zhang, H.; Wang, C.; Cao, H.; Huang, L.; Wang, Z. Structural characterization and antioxidant activities of κ-carrageenan oligosaccharides degraded by different methods. Food Chem. 2015, 178, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yan, X.; Lin, J.; Wang, F.; Xu, W. Depolymerized products of lambda-carrageenan as a potent angiogenesis inhibitor. J. Agric. Food Chem. 2007, 55, 6910–6917. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.-T.; Zhang, D.-S.; Chen, H.-M.; Yan, X.-J. Modulation of the binding of basic fibroblast growth factor and heparanase activity by purified λ-carrageenan oligosaccharides. Carbohydr. Polym. 2015, 125, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Shao, B.; Nie, W.; Wei, X.-W.; Li, Y.-L.; Wang, B.-L.; He, Z.-Y.; Liang, X.; Ye, T.-H.; Wei, Y.-Q. Antitumor and Adjuvant Activity of λ-carrageenan by Stimulating Immune Response in Cancer Immunotherapy. Sci. Rep. 2015, 5, 11062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Sheng, W.; Yao, W.; Wang, C. Effect of low molecular λ-carrageenan from Chondrus ocellatus on antitumor H-22 activity of 5-Fu. Pharmacol. Res. 2006, 53, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Xin, H.; Sheng, W.; Sun, Y.; Li, Z.; Xu, Z. In vivo growth-inhibition of S180 tumor by mixture of 5-Fu and low molecular λ-carrageenan from. Pharmacol. Res. 2005, 51, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-M.; Yan, X.-J.; Mai, T.-Y.; Wang, F.; Xu, W.-F. Lambda-carrageenan oligosaccharides elicit reactive oxygen species production resulting in mitochondrial-dependent apoptosis in human umbilical vein endothelial cells. Int. J. Mol. Med. 2009, 24, 801–806. [Google Scholar] [PubMed]
- Jazzara, M.; Ghannam, A.; Soukkarieh, C.; Murad, H. Anti-Proliferative Activity of λ-Carrageenan Through the Induction of Apoptosis in Human Breast Cancer Cells. Iran. J. Cancer Prev. 2016, 9, e3836. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R. Carrageenans inhibit growth-factor binding. Biochem. J. 1993, 289 Pt 2, 331–334. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, R.; Burns, W.W.; Paper, D.H. Selective inhibition of cell proliferation and DNA synthesis by the polysulphated carbohydrate l-carrageenan. Cancer Chemother. Pharm. 1995, 36, 325–334. [Google Scholar] [CrossRef]
- Kosir, M.A.; Wang, W.; Zukowski, K.L.; Tromp, G.; Barber, J. Degradation of basement membrane by prostate tumor heparanase. J. Surg. Res. 1999, 81, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Parish, C.R.; Coombe, D.R.; Jakobsen, K.B.; Bennett, F.A.; Underwood, P.A. Evidence that sulphated polysaccharides inhibit tumour metastasis by blocking tumour-cell-derived heparanases. Int. J. Cancer 1987, 40, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Poupard, N.; Badarou, P.; Fasani, F.; Groult, H.; Bridiau, N.; Sannier, F.; Bordenave-Juchereau, S.; Kieda, C.; Piot, J.-M.; Grillon, C.; et al. Assessment of Heparanase-Mediated Angiogenesis Using Microvascular Endothelial Cells: Identification of λ-Carrageenan Derivative as a Potent Anti Angiogenic Agent. Mar. Drugs 2017, 15, 134. [Google Scholar] [CrossRef] [PubMed]
- Poupard, N.; Groult, H.; Bodin, J.; Bridiau, N.; Bordenave-Juchereau, S.; Sannier, F.; Piot, J.-M.; Fruitier-Arnaudin, I.; Maugard, T. Production of heparin and λ-carrageenan anti-heparanase derivatives using a combination of physicochemical depolymerization and glycol splitting. Carbohydr. Polym. 2017, 166, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Marine Glycobiology: Principles and Applications; Kim, S.-K. (Ed.) CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: London, UK; New York, NY, USA, 2017; ISBN 978-1-4987-0961-3. [Google Scholar]
- Navarro, D.A.; Flores, M.L.; Stortz, C.A. Microwave-assisted desulfation of sulfated polysaccharides. Carbohydr. Polym. 2007, 69, 742–747. [Google Scholar] [CrossRef]
- Sokolova, E.V.; Byankina, A.O.; Kalitnik, A.A.; Kim, Y.H.; Bogdanovich, L.N.; Solov’eva, T.F.; Yermak, I.M. Influence of red algal sulfated polysaccharides on blood coagulation and platelets activation in vitro: Influence of Red Algal Sulfated Polysaccharides. J. Biomed. Mater. Res. Part A 2014, 102, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Ciancia, M.; Quintana, I.; Cerezo, A.S. Overview of Anticoagulant Activity of Sulfated Polysaccharides from Seaweeds in Relation to their Structures, Focusing on those of Green Seaweeds. Curr. Med. Chem. 2010, 17, 2503–2529. [Google Scholar] [CrossRef] [PubMed]
- Hb25_Springer Handbook of Marine Biotechnology; Kim, S.-K. (Ed.) Springer: Berlin/Heidelberg, Germany, 2015; ISBN 978-3-642-53970-1. [Google Scholar]
- Melo, F.R.; Pereira, M.S.; Foguel, D.; Mourão, P.A.S. Antithrombin-mediated Anticoagulant Activity of Sulfated Polysaccharides: Different mechanisms for heparin and sulfated galactans. J. Biol. Chem. 2004, 279, 20824–20835. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Suflita, M.; Linhardt, R.J. Bioengineered heparins and heparan sulfates. Adv. Drug Deliv. Rev. 2016, 97, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Araújo, C.A.; Noseda, M.D.; Cipriani, T.R.; Gonçalves, A.G.; Duarte, M.E.R.; Ducatti, D.R.B. Selective sulfation of carrageenans and the influence of sulfate regiochemistry on anticoagulant properties. Carbohydr. Polym. 2013, 91, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Mao, X.; Peng, X.; Tang, S. Effects of sulfate group in red seaweed polysaccharides on anticoagulant activity and cytotoxicity. Carbohydr. Polym. 2014, 101, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Fernández, P.V.; Quintana, I.; Cerezo, A.S.; Caramelo, J.J.; Pol-Fachin, L.; Verli, H.; Estevez, J.M.; Ciancia, M. Anticoagulant activity of a unique sulfated pyranosic (1->3)-β-l-arabinan through direct interaction with thrombin. J. Biol. Chem. 2013, 288, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Matsuura, Y.; Bacic, A.; Liao, M.; Hori, K.; Miyazawa, K. Anticoagulant properties of a sulfated galactan preparation from a marine green alga, Codium cylindricum. Int. J. Biol. Macromol. 2001, 28, 395–399. [Google Scholar] [CrossRef]
- Dai, X.; Yan, J.; Fu, X.; Pan, Q.; Sun, D.; Xu, Y.; Wang, J.; Nie, L.; Tong, L.; Shen, A.; et al. Aspirin Inhibits Cancer Metastasis and Angiogenesis via Targeting Heparanase. Clin. Cancer Res. 2017, 23, 6267–6278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.M.; Shen, Z.H.; Liu, Z.Y.; Wang, F.; Hai, L.; Gao, L.T.; Wang, H.S. Heparanase promotes human gastric cancer cells migration and invasion by increasing Src and p38 phosphorylation expression. Int. J. Clin. Exp. Pathol. 2014, 7, 5609–5621. [Google Scholar] [PubMed]
- Li, Y.; Liu, H.; Huang, Y.Y.; Pu, L.J.; Zhang, X.D.; Jiang, C.C.; Jiang, Z.W. Suppression of endoplasmic reticulum stress-induced invasion and migration of breast cancer cells through the downregulation of heparanase. Int. J. Mol. Med. 2013, 31, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.; Harvey, J.R.; Murphy, K.J.; Pye, D.; O’Boyle, G.; Lennard, T.W.J.; Kirby, J.A.; Ali, S. Modulatory effects of heparin and short-length oligosaccharides of heparin on the metastasis and growth of LMD MDA-MB 231 breast cancer cells in vivo. Br. J. Cancer 2007, 97, 761–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettelaie, C.; Fountain, D.; Collier, M.E.W.; Beeby, E.; Xiao, Y.P.; Maraveyas, A. Low molecular weight heparin suppresses tissue factor-mediated cancer cell invasion and migration in vitro. Exp. Ther. Med. 2011, 2, 363–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponert, J.; Gockel, L.; Henze, S.; Schlesinger, M. Unfractionated and Low Molecular Weight Heparin Reduce Platelet Induced Epithelial-Mesenchymal Transition in Pancreatic and Prostate Cancer Cells. Molecules 2018, 23, 2690. [Google Scholar] [CrossRef] [PubMed]
- Nurcombe, V.; Smart, C.E.; Chipperfield, H.; Cool, S.M.; Boilly, B.; Hondermarck, H. The Proliferative and Migratory Activities of Breast Cancer Cells Can Be Differentially Regulated by Heparan Sulfates. J. Biol. Chem. 2000, 275, 30009–30018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, M.; Brüggemann, K.; Karousou, E.; Caon, I.; Caravà, E.; Vigetti, D.; Greve, B.; Stock, C.; De Luca, G.; Passi, A.; et al. MDA-MB-231 breast cancer cell viability, motility and matrix adhesion are regulated by a complex interplay of heparan sulfate, chondroitin−/dermatan sulfate and hyaluronan biosynthesis. Glycoconj. J. 2017, 34, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Mulloy, B.; Hogwood, J. Chromatographic molecular weight measurements for heparin, its fragments and fractions, and other glycosaminoglycans. Methods Mol. Biol. 2015, 1229, 105–118. [Google Scholar] [PubMed]
- Gao, G.; Jiao, Q.; Ding, Y.; Chen, L. Study on quantitative assay of chondroitin sulfate with a spectrophotometric method of azure A. Guang Pu Xue Yu Guang Pu Fen Xi 2003, 23, 600–602. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Time (h) | Mn (KDa) | Mw (KDa) | DP | I | DS% (w/w) |
|---|---|---|---|---|---|---|
| Depolymerisation at 60 °C | 0 | 2585.9 | 2722.2 | 4463 | 1.1 | ~30 |
| 2 | 931.3 | 1120.2 | 1607 | 1.2 | 29.5 ± 0.5 | |
| 4 | 752.5 | 987.2 | 1299 | 1.3 | 29.7 ± 0.5 | |
| 8 | 261.4 | 425.8 | 451 | 1.6 | 30.9 ± 0.7 | |
| 12 | 31.7 | 42.7 | 55 | 1.4 | 28.0 ± 0.6 | |
| 16 | 17.8 | 23.8 | 31 | 1.3 | 22.3 ± 0.7 | |
| 24 | 10.0 | 12.7 | 17 | 1.3 | 20.3 ± 0.8 | |
| 36 | 5.9 | 8.4 | 10 | 1.4 | 17.8 ± 0.9 | |
| 48 | 4.3 | 6.8 | 7 | 1.6 | 15.9 ± 0.8 | |
| 72 | 2.8 | 3.7 | 5 | 1.4 | 8.7 ± 0.6 | |
| Depolymerisation at 40 °C | 0 | 2585.9 | 2722.2 | 4463 | 1.1 | ~30 |
| 24 | 130.3 | 197.2 | 225 | 1.5 | 27.6 ± 0.5 | |
| 48 | 74.5 | 110.4 | 129 | 1.5 | 27.6 ± 0.7 | |
| 72 | 23.9 | 36.2 | 41 | 1.5 | 29.2 ± 0.8 | |
| 96 | 13.0 | 19.2 | 22 | 1.5 | 23.8 ± 0.6 | |
| 120 | 11.7 | 17.4 | 20 | 1.5 | 24.6 ± 0.8 | |
| 144 | 9.5 | 14.3 | 16 | 1.5 | 23.7 ± 0.8 | |
| 168 | 7.6 | 11.2 | 13 | 1.5 | 22.5 ± 0.8 | |
| 240 | 5.5 | 8.0 | 10 | 1.4 | 19.0 ± 1.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groult, H.; Cousin, R.; Chot-Plassot, C.; Maura, M.; Bridiau, N.; Piot, J.-M.; Maugard, T.; Fruitier-Arnaudin, I. ?-Carrageenan Oligosaccharides of Distinct Anti-Heparanase and Anticoagulant Activities Inhibit MDA-MB-231 Breast Cancer Cell Migration. Mar. Drugs 2019, 17, 140. https://doi.org/10.3390/md17030140
Groult H, Cousin R, Chot-Plassot C, Maura M, Bridiau N, Piot J-M, Maugard T, Fruitier-Arnaudin I. ?-Carrageenan Oligosaccharides of Distinct Anti-Heparanase and Anticoagulant Activities Inhibit MDA-MB-231 Breast Cancer Cell Migration. Marine Drugs. 2019; 17(3):140. https://doi.org/10.3390/md17030140
Chicago/Turabian StyleGroult, Hugo, Rémi Cousin, Caroline Chot-Plassot, Maheva Maura, Nicolas Bridiau, Jean-Marie Piot, Thierry Maugard, and Ingrid Fruitier-Arnaudin. 2019. "?-Carrageenan Oligosaccharides of Distinct Anti-Heparanase and Anticoagulant Activities Inhibit MDA-MB-231 Breast Cancer Cell Migration" Marine Drugs 17, no. 3: 140. https://doi.org/10.3390/md17030140
APA StyleGroult, H., Cousin, R., Chot-Plassot, C., Maura, M., Bridiau, N., Piot, J.-M., Maugard, T., & Fruitier-Arnaudin, I. (2019). ?-Carrageenan Oligosaccharides of Distinct Anti-Heparanase and Anticoagulant Activities Inhibit MDA-MB-231 Breast Cancer Cell Migration. Marine Drugs, 17(3), 140. https://doi.org/10.3390/md17030140