Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro

 and
and
Abstract
1. Introduction
2. Results
2.1. Actinomycin V Inhibits the Proliferation of Human Breast Cancer Cells
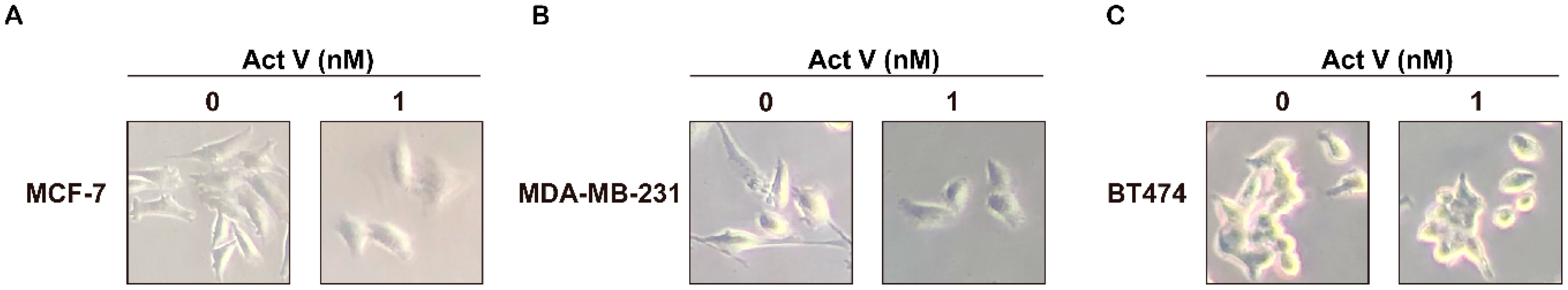
2.2. Actinomycin V’s Effects on Morphological Changes in Breast Cancer Cells
2.3. Actinomycin V Inhibits the Migration and Invasion of Human Breast Cancer Cells
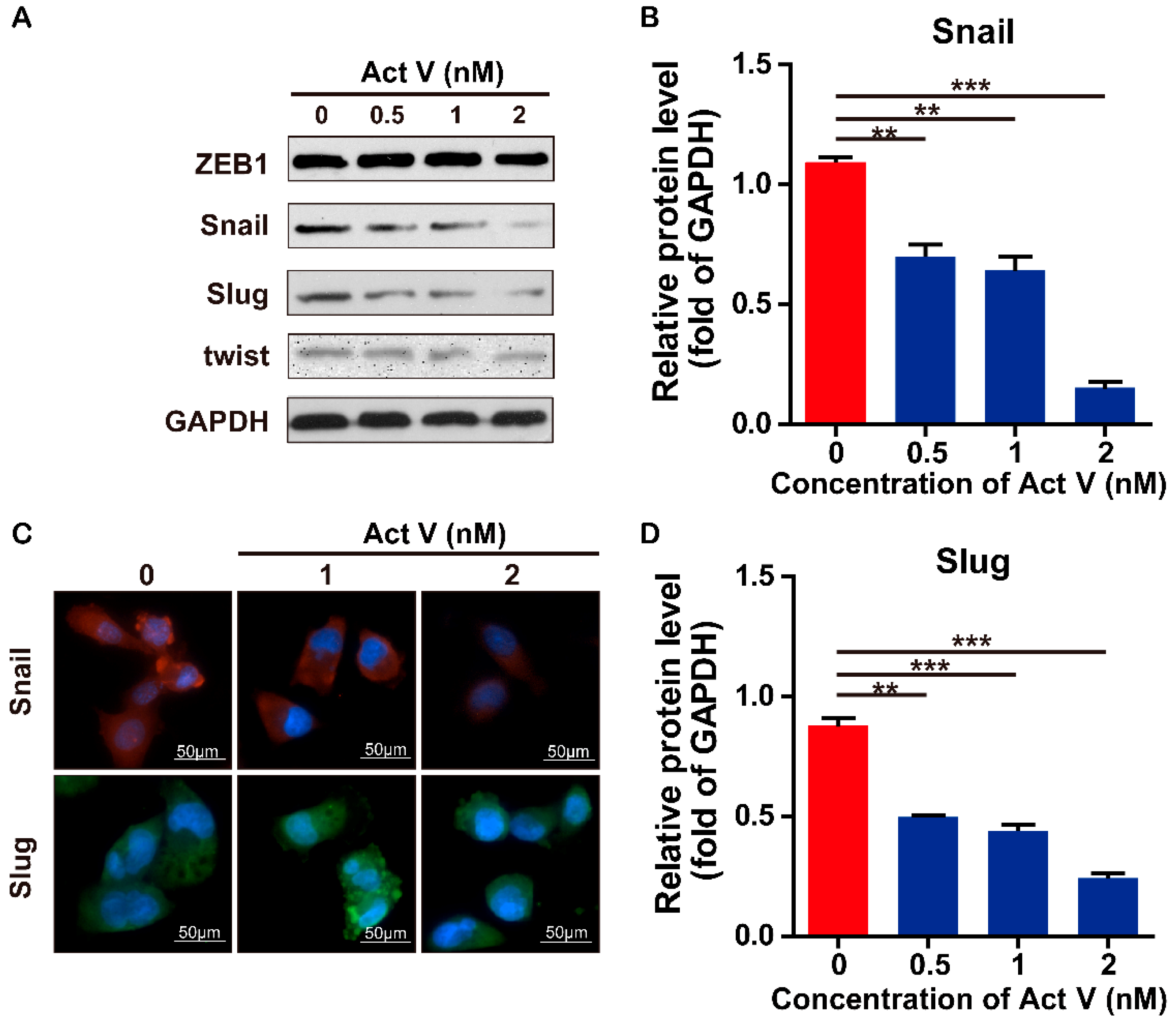
2.4. Effects of Actinomycin V on the Expression of EMT-Associated Proteins and mRNA in MDA-MB-231 Cells
2.5. Actinomycin V Suppresses the EMT Process by Reducing the Expression of SNAIL and SLUG
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines and Cell Culture
4.3. MTT Assay
4.4. Wound-Healing Assay
4.5. Transwell Assay
4.6. Immunofluorescence Staining
4.7. Western Blot Analysis
4.8. RNA Extraction and Relative Quantification by Real Time PCR
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Goding Sauer, A.; Ortiz, A.P.; Fedewa, S.A.; Pinheiro, P.S.; Tortolero-Luna, G.; Martinez-Tyson, D.; Jemal, A.; Siegel, R.L. Cancer Statistics for Hispanics/Latinos, 2018. CA Cancer J. Clin. 2018, 68, 425–445. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.-J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Gonen, M.; Kim, H.J.; Michor, F.; Polyak, K. Cellular and genetic diversity in the progression of in situ human breast carcinomas to an invasive phenotype. J. Clin. Investig. 2010, 120, 636–644. [Google Scholar] [CrossRef] [PubMed]
- Fehm, T.; Muller, V.; Alix-Panabieres, C.; Pantel, K. Micrometastatic spread in breast cancer: Detection, molecular characterization and clinical relevance. Breast Cancer Res. 2008, 10, S1. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Guan, J.L. Focal adhesion kinase: A prominent determinant in breast cancer initiation, progression and metastasis. Cancer Lett. 2010, 289, 127–139. [Google Scholar] [CrossRef]
- Xu, Y.J.; Bismar, T.A.; Su, J.; Xu, B.; Kristiansen, G.; Varga, Z.; Teng, L.H.; Ingber, D.E.; Mammoto, A.; Kumar, R.; Alaoui-Jamali, M.A. Filamin A regulates focal adhesion disassembly and suppresses breast cancer cell migration and invasion. J. Exp. Med. 2010, 207, 2421–2437. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ye, X.; Chai, W.; Lian, X.-Y.; Zhang, Z. New Metabolites and Bioactive Actinomycins from Marine-Derived Streptomyces sp. ZZ338. Mar. Drugs 2016, 14, 181. [Google Scholar] [CrossRef] [PubMed]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2004, 21, 519–538. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef]
- Brockmann, H. actinomycines. Angew. Chem. Int. Edit. 1960, 72, 447–939. [Google Scholar]
- Muller, W.; Crothers, D.M. Studies of binding of actinomycin and related compounds to DNA. J. Mol. Biol. 1968, 35, 251–290. [Google Scholar] [CrossRef]
- Cai, W.L.; Wang, X.C.; Elshahawi, S.I.; Ponomareva, L.V.; Liu, X.D.; McErlean, M.R.; Cui, Z.; Arlinghaus, A.L.; Thorson, J.S.; Van Lanen, S.G. Antibacterial and Cytotoxic Actinomycins Y-6–Y-9 and Zp from Streptomyces sp. Strain Go–GS12. J. Nat. Prod. 2016, 79, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Farber, S.; Dangio, G.; Evans, A.; Mitus, A. Clinical significance.3. clinical studies of actinomycin–D with special reference to wilms tumor in children. Ann. N. Y. Acad. Sci. 1960, 89, 421–425. [Google Scholar] [CrossRef]
- Sliva, D.; Rizzo, M.T.; English, D. Phosphatidylinositol 3-kinase and NF-kappa B regulate motility of invasive MDA-AM-231 human breast cancer cells by the secretion of urokinase-type plasminogen activator. J. Biol. Chem. 2002, 277, 3150–3157. [Google Scholar] [CrossRef] [PubMed]
- Morioka, H.; Takezawa, M.; Shibai, H. Actinomycin-V as a potent differentiation inducer of F5-5 friend-leukemia cells. Agric. Biol. Chem. 1985, 49, 2835–2842. [Google Scholar]
- Wang, D.; Wang, C.; Gui, P.; Liu, H.; Khalaf, S.M.H.; Elsayed, E.A.; Wadaan, M.A.M.; Hozzein, W.N.; Zhu, W. Identification, Bioactivity, and Productivity of Actinomycins from the Marine-Derived Streptomyces heliomycini. Front. Microbiol. 2017, 8, 1147–1158. [Google Scholar] [CrossRef]
- Blanco, M.J.; Moreno-Bueno, G.; Sarrio, D.; Locascio, A.; Cano, A.; Palacios, J.; Nieto, M.A. Correlation of Snail expression with histological grade and lymph node status in breast carcinomas. Oncogene 2002, 21, 3241–3246. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.S.T.; Gumbiner, B.M. Adhesion-independent mechanism for suppression of tumor cell invasion by E-cadherin. J. Cell Biol. 2003, 161, 1191–1203. [Google Scholar] [CrossRef]
- Geho, D.H.; Bandle, R.W.; Clair, T.; Liotta, L.A. Physiological mechanisms of tumor-cell invasion and migration. Physiology 2005, 20, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Kobayashi, E.; Takamatsu, Y.; Li, S.H.; Hatano, T.; Sakagami, H.; Kusama, K.; Satoh, K.; Sugita, D.; Shimura, S.; Itoh, Y.; Yoshida, T. Polyphenols from Eriobotrya japonica and their cytotoxicity against human oral tumor cell lines. Chem. Pharm. Bull. 2000, 48, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Oliveira, P.; Cabral, J.; Carvalho, S.; Huntsman, D.; Garter, F.; Seruca, R.; Reis, C.A.; Oliveira, C. Loss and Recovery of Mgat3 and GnT-III Mediated E-cadherin N-glycosylation Is a Mechanism Involved in Epithelial-Mesenchymal-Epithelial Transitions. PLoS ONE 2012, 7, e33191. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yan, Y.; Cheng, Z.; Hu, Y.; Liu, T. Sotetsuflavone suppresses invasion and metastasis in non–small-cell lung cancer A549 cells by reversing EMT via the TNF-alpha/NF-kappaB and PI3K/AKT signaling pathway. Cell Death Discov. 2018, 4, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Derycke, L.D.M.; Bracke, M.E. N-cadherin in the spotlight of cell-cell adhesion, differentiation, embryogenesis, invasion and signalling. Int. J. Dev. Biol. 2004, 48, 463–476. [Google Scholar] [CrossRef]
- Satelli, A.; Li, S.L. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell. Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [PubMed]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, ZEB and bHLH factors in tumour progression: An alliance against the epithelial phenotype? Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Alidadiani, N.; Ghaderi, S.; Dilaver, N.; Bakhshamin, S.; Bayat, M. Epithelial mesenchymal transition Transcription Factor (TF): The structure, function and microRNA feedback loop. Gene 2018, 674, 115–120. [Google Scholar] [CrossRef]
- Wei, Z.; Shan, Z.; Shaikh, Z.A. Epithelial-mesenchymal transition in breast epithelial cells treated with cadmium and the role of Snail. Toxicol. Appl. Pharm. 2018, 344, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Perez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; del Barrio, M.G.; Portillo, F.; Nieto, M.A. The transcription factor Snail controls epithelial–mesenchymal transitions by repressing E-cadherin expression. Nat. Cell Biol. 2000, 2, 76–83. [Google Scholar] [CrossRef]
- Batlle, E.; Sancho, E.; Franci, C.; Dominguez, D.; Monfar, M.; Baulida, J.; de Herreros, A.G. The transcription factor Snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nat. Cell Biol. 2000, 2, 84–89. [Google Scholar] [CrossRef]
- Lu, D.D.; Ren, J.W.; Du, Q.Q.; Song, Y.J.; Lin, S.Q.; Li, X.; Xie, W.D. p-Terhenyls and actinomycins from a Streptomyces sp. associated with the Iarva of mud dauber wasp. Nat. Prod. Res. 2019, in press. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds IC50 (nmol/L) | MCF-7 | MDA-MB-231 | BT474 | MCF-10A | HMLE |
|---|---|---|---|---|---|
| Actinomycin V | 1.92 ± 0.25 | 0.83 ± 0.32 | 4.16 ± 0.25 | 4.07 ± 0.26 | 3.49 ± 0.31 |
| Actinomycin D | 8.23 ± 0.50 | 15.15 ± 0.52 | 37.00 ± 3.15 | 34.01 ± 0.25 | 30.22 ± 0.50 |
| Actinomycin Xob | 149.40 ± 4.03 | 127.33 ± 4.49 | 369.90 ± 0.14 | 248.57 ± 14.69 | 105.83 ± 8.44 |
| Adriamycin | 885.38 ± 13.50 | 942.60 ± 22.50 | 584.70 ± 50.00 | 1489.13 ± 25.50 | 1627 ± 15.50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Mar. Drugs 2019, 17, 305. https://doi.org/10.3390/md17050305
Lin S, Zhang C, Liu F, Ma J, Jia F, Han Z, Xie W, Li X. Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Marine Drugs. 2019; 17(5):305. https://doi.org/10.3390/md17050305
Chicago/Turabian StyleLin, Shiqi, Caiyun Zhang, Fangyuan Liu, Jiahui Ma, Fujuan Jia, Zhuo Han, Weidong Xie, and Xia Li. 2019. "Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro" Marine Drugs 17, no. 5: 305. https://doi.org/10.3390/md17050305
APA StyleLin, S., Zhang, C., Liu, F., Ma, J., Jia, F., Han, Z., Xie, W., & Li, X. (2019). Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Marine Drugs, 17(5), 305. https://doi.org/10.3390/md17050305