Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects

 ,
,
Abstract
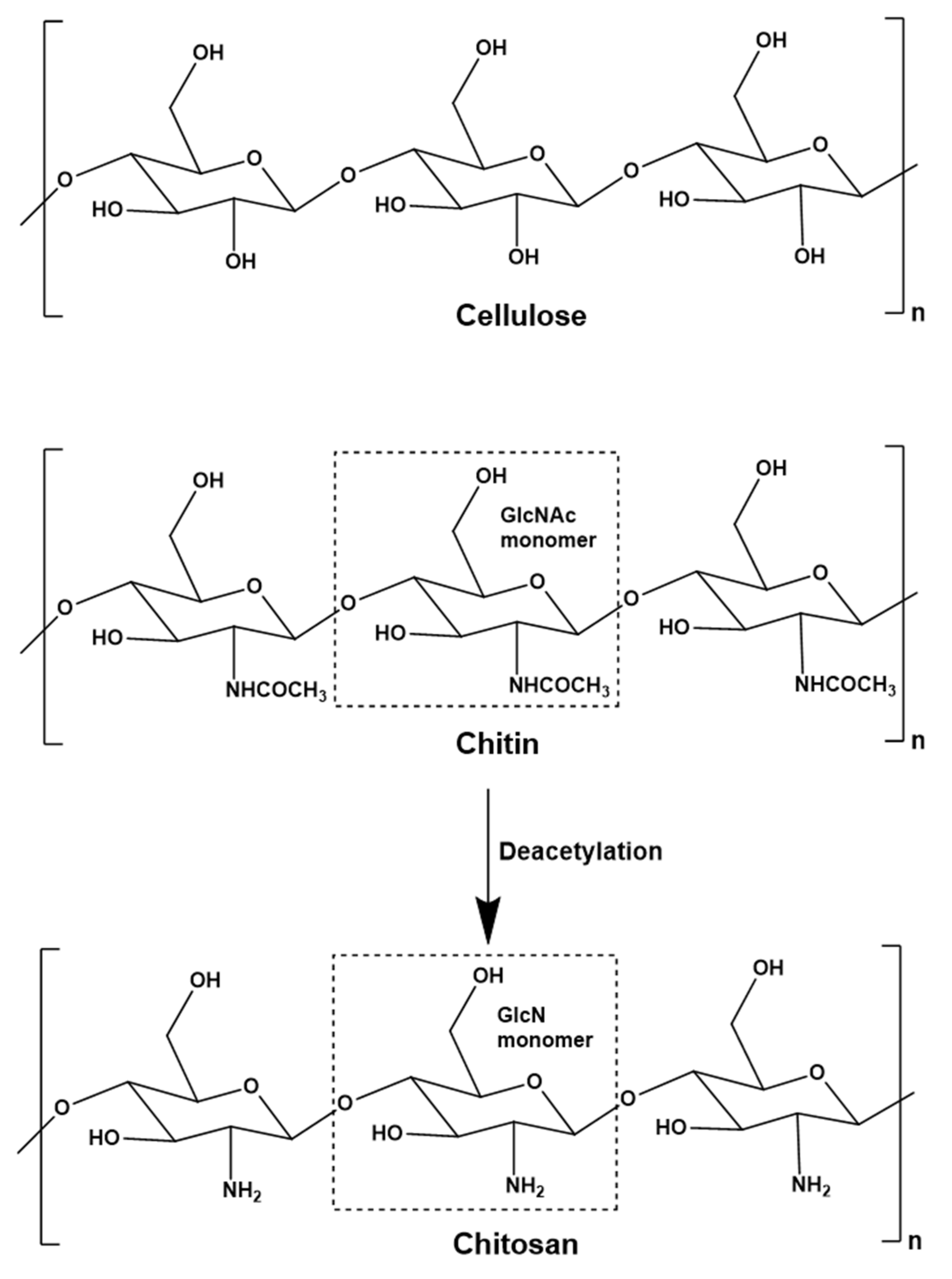
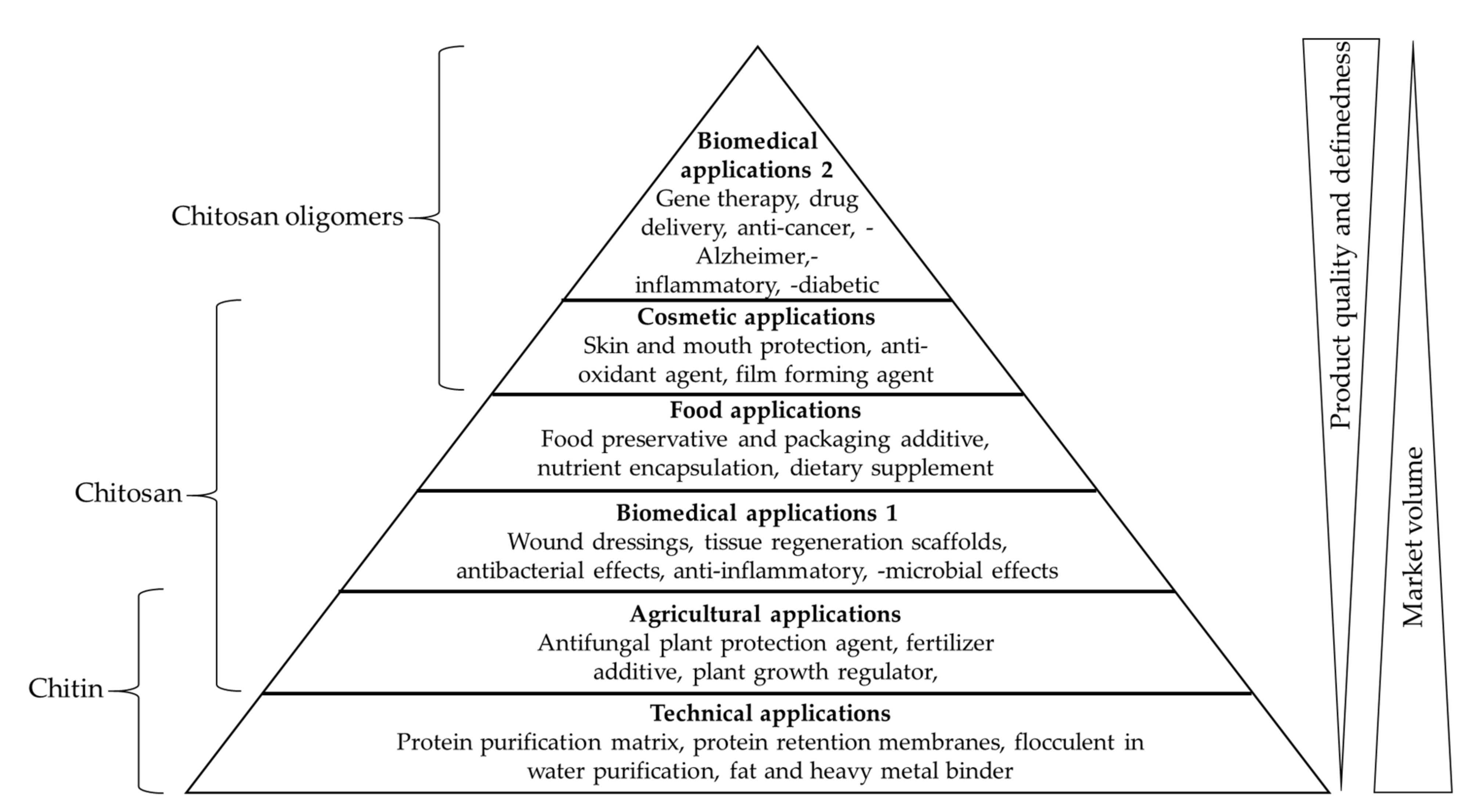
:1. Introduction—Chitin, Chitosan and Chitosan Oligomers
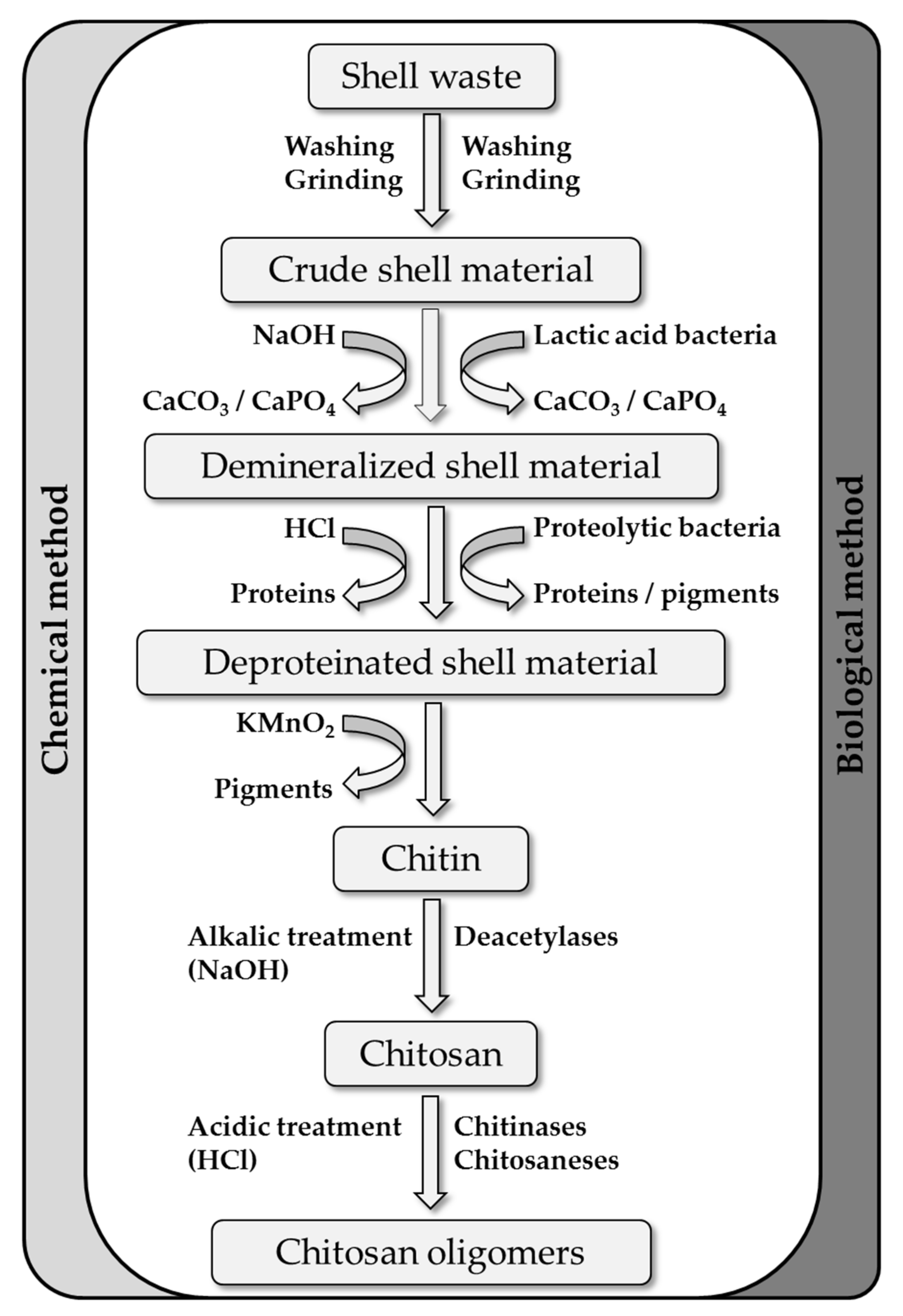
2. Chitin Extraction from Marine Biowaste
3. Chemical Conversion of Chitin to COS
3.1. Chemical Deacetylation of Chitin
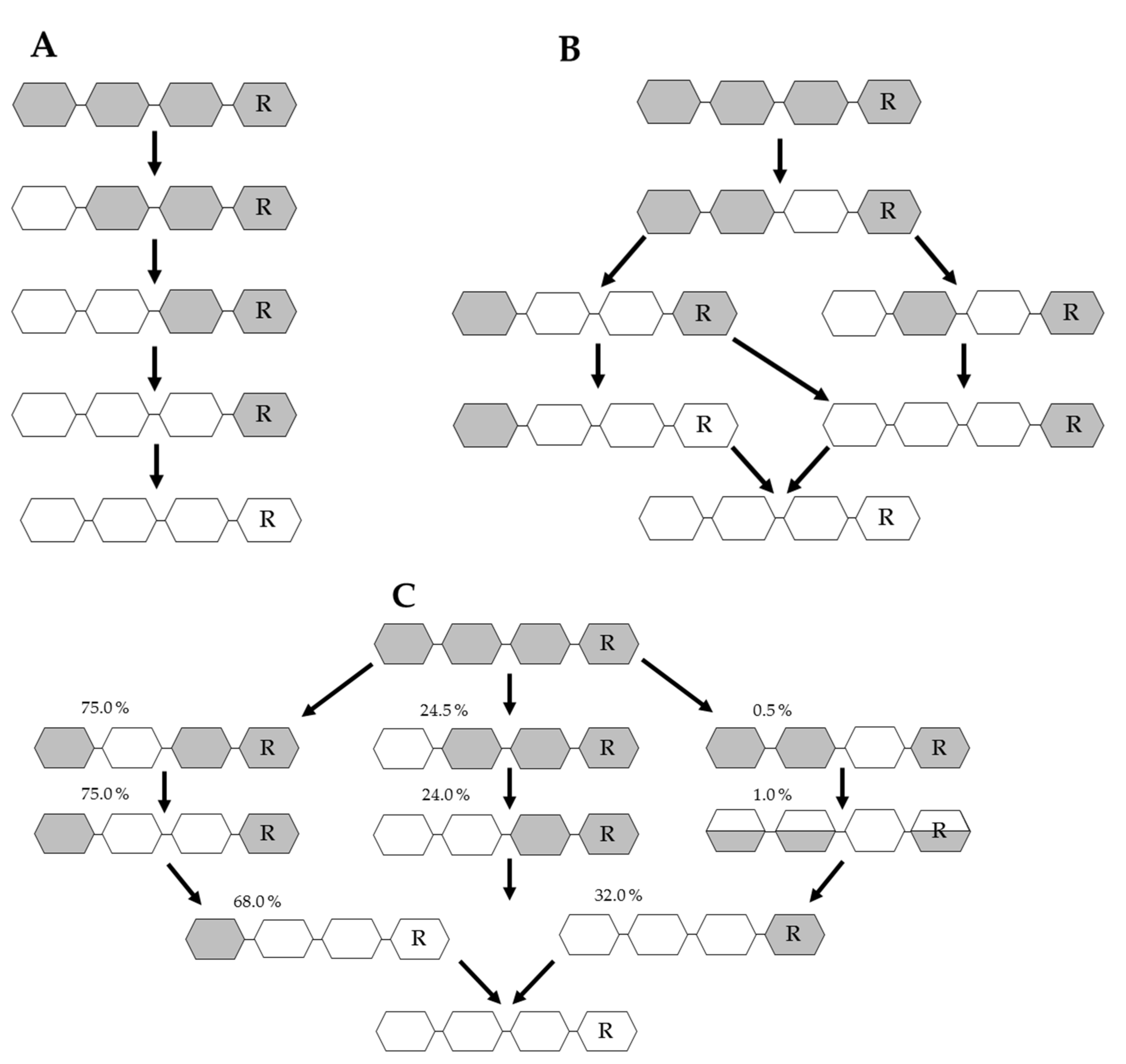
3.2. Chemical Depolymerization of Chitosan
4. Biological Conversion of Chitin to COS
4.1. Enzymatic Deacetylation of Chitin
4.2. Enzymatic Chitin/Chitosan Depolymerization
5. Fully Enzymatic COS Production
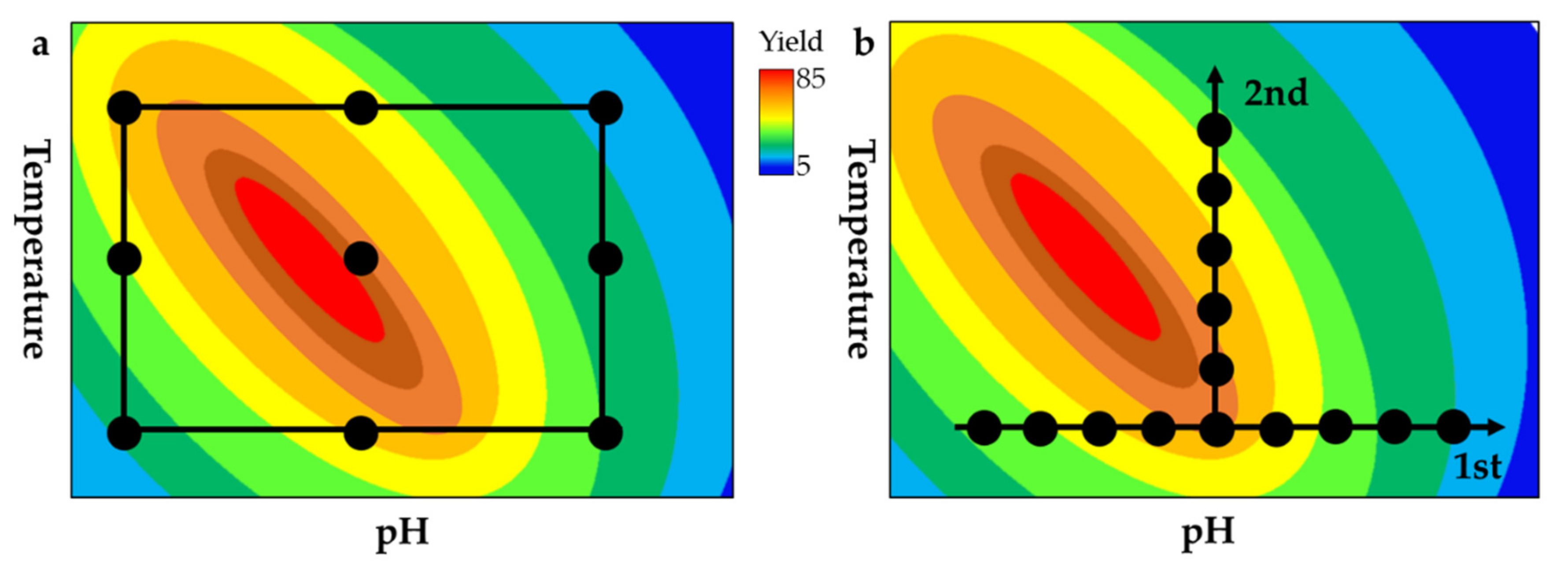
6. Design-of-experiments Approach for Multi-enzyme Process Optimization
7. Conclusion and Future Prospects
Funding
Acknowledgments
Conflicts of Interest
References
- Tharanathan, R.N.; Kittur, F.S. Chitin—The undisputed biomolecule of great potential. Crit. Rev. Food Sci. Nutr. 2003, 43, 61–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K. Chitin, Chitosan, Oligosaccarides and Their Derivatives; CRC Press: Boca Raton, FL, USA, 2011; Volume 1. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Minke, R.; Blackwell, J. The structure of α-chitin. J. Mol. Biol. 1978, 120, 167–181. [Google Scholar] [CrossRef]
- Dweltz, N.E. The structure of β-chitin. Biochim. Biophys. Acta 1961, 51, 283–294. [Google Scholar] [CrossRef]
- Bloch, R.; Burger, M.M. Purification of wheat germ agglutinin using affinity chromatography on chitin. Biochem. Biophys. Res. Commun. 1974, 58, 13–19. [Google Scholar] [CrossRef]
- Wolman, F.J.; Copello, G.J.; Mebert, A.M.; Targovnik, A.M.; Miranda, M.V.; Navarro del Cañizo, A.A.; Díaz, L.E.; Cascone, O. Egg white lysozyme purification with a chitin–silica-based affinity chromatographic matrix. Eur. Food Res. Technol. 2010, 231, 181–188. [Google Scholar] [CrossRef]
- Brzezinska, M.S.; Jankiewicz, U. Production of antifungal chitinase by Aspergillus niger LOCK 62 and its potential role in the biological control. Curr. Microbiol. 2012, 65, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Ruckenstein, E.; Zeng, X. Macroporous chitin affinity membranes for lysozyme separation. Biotechnol. Bioeng. 1997, 56, 610–617. [Google Scholar] [CrossRef]
- Krajewska, B. Application of chitin- and chitosan-based materials for enzyme immobilizations: A review. Enzym. Microb. Technol. 2004, 35, 126–139. [Google Scholar] [CrossRef]
- Winkler, A.J.; Dominguez-Nunez, J.A.; Aranaz, I.; Poza-Carrion, C.; Ramonell, K.; Somerville, S.; Berrocal-Lobo, M. Short-Chain Chitin Oligomers: Promoters of Plant Growth. Mar. Drugs 2017, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Tsakalidi, A.L.; Chalikiopoulos, D.; Papasavvas, A. Effect of chitin on growth and chlorophyll content of two medicinal plants. J. Med. Plant. Res. 2010, 4, 499–508. [Google Scholar]
- No, H.K.; Park, N.Y.; Lee, S.H.; Meyer, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trends Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Gutiérrez, T.J. Chitosan Applications for the Food Industry. In Chitosan: Derivatives, Composites and Applications, Chapter 8; Wiley Online Library: Hoboken, NJ, USA, 2017; pp. 183–232. [Google Scholar]
- Aranaz, I.; Acosta, N.; Civera, C.; Elorza, B.; Mingo, J.; Castro, C.; De los Llano Gandia, M.; Heras Caballer, A. Cosmetics and cosmeceutical applications of chitin, chitosan and their derivatives. Polymers 2018, 10, 213. [Google Scholar] [CrossRef] [PubMed]
- Gumilar, T.A.; Prihastanti, E.; Haryanti, S.; Subagio, A.; Ngadiwiyana, A. Utilization of waste silica and chitosan as fertilizer nano chisil to improve corn production in Indonesia. Adv. Sci. Lett. 2017, 23, 2447–2449. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Recent Advances of Chitosan Applications in Plants. Polymers 2018, 10, 118. [Google Scholar] [CrossRef]
- Chen, Y.-E.; Yuan, S.; Liu, H.-M.; Chen, Z.-Y.; Zhang, Y.-H.; Zhang, H.-Y. A combination of chitosan and chemical fertilizers improves growth and disease resistance in Begonia × hiemalis Fotsch. Hortic. Environ. Biotechnol. 2016, 57, 1–10. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Nechita, P. Applications of Chitosan in Wastewater Treatment. Biol. Act. Appl. Mar. Polysacch. 2017, 209–228. [Google Scholar]
- Badawi, M.A.; Negm, N.A.; Abou Kana, M.T.H.; Hefni, H.H.; Abdel Moneem, M.M. Adsorption of aluminum and lead from wastewater by chitosan-tannic acid modified biopolymers: Isotherms, kinetics, thermodynamics and process mechanism. Int. J. Biol Macromol. 2017, 99, 465–476. [Google Scholar] [CrossRef]
- Chi, F.H.; Cheng, W.P. Use of Chitosan as Coagulant to Treat Wastewater from Milk Processing Plant. J. Polym. Environ. 2006, 14, 411–417. [Google Scholar] [CrossRef]
- Jayasantha Kumari, H.; Krishnamoorthy, P.; Arumugam, T.K.; Radhakrishnan, S.; Vasudevan, D. An efficient removal of crystal violet dye from waste water by adsorption onto TLAC/Chitosan composite: A novel low cost adsorbent. Int. J. Biol. Macromol. 2017, 96, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Park, B.K.; Kim, M.M. Applications of chitin and its derivatives in biological medicine. Int. J. Mol. Sci. 2010, 11, 5152–5164. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Izumi, R.; Osaki, T.; Ifuku, S.; Morimoto, M.; Saimoto, H.; Minami, S.; Okamoto, Y. Chitin, chitosan, and its derivatives for wound healing: Old and new materials. J. Funct. Biomater. 2015, 6, 104–142. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.L. Chitin-based materials in tissue engineering: Applications in soft tissue and epithelial organ. Int. J. Mol. Sci. 2011, 12, 1936–1963. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, R.; Chennazhi, K.P.; Muzzarelli, R.A.A.; Tamura, H.; Nair, S.V.; Selvamurugan, N. Chitosan conjugated DNA nanoparticles in gene therapy. Carbohydr. Polym. 2010, 79, 1–8. [Google Scholar] [CrossRef]
- Hamedi, H.; Moradi, S.; Hudson, S.M.; Tonelli, A.E. Chitosan based hydrogels and their applications for drug delivery in wound dressings: A review. Carbohydr. Polym. 2018, 199, 445–460. [Google Scholar] [CrossRef]
- Abdel-Rahman, R.M.; Abdel-Mohsen, A.M.; Hrdina, R.; Burgert, L.; Fohlerova, Z.; Pavlinak, D.; Sayed, O.N.; Jancar, J. Wound dressing based on chitosan/hyaluronan/nonwoven fabrics: Preparation, characterization and medical applications. Int. J. Biol. Macromol. 2016, 89, 725–736. [Google Scholar] [CrossRef]
- Cheung, R.C.; Ng, T.B.; Wong, J.H.; Chan, W.Y. Chitosan: An Update on Potential Biomedical and Pharmaceutical Applications. Mar. Drugs 2015, 13, 5156–5186. [Google Scholar] [CrossRef]
- Wu, Y.; Lin, Q.L.; Chen, Z.X.; Wu, W.; Xiao, H.X. Preparation of chitosan oligomers COS and their effect on the retrogradation of intermediate amylose rice starch. J. Food Sci. Technol. 2012, 49, 695–703. [Google Scholar] [CrossRef]
- Park, J.K.; Chung, M.J.; Choi, H.N.; Park, Y.I. Effects of the molecular weight and the degree of deacetylation of chitosan oligosaccharides on antitumor activity. Int. J. Mol. Sci. 2011, 12, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Moerschbacher, B.M. The cell factory approach toward biotechnological production of high-value chitosan oligomers and their derivatives: An update. Crit. Rev. Biotechnol. 2017, 37, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.-J.; Shahidi, F.; Kim, S.-K. Preparation of Chitin and Chitosan Oligomers and Their Applications in Physiological Functional Foods. Food Rev. Int. 2000, 16, 159–176. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Eaton, P.; Nascimento, H.; Gião, M.S.; Ramos, O.S.; Belo, L.; Santos-Silva, A.; Pintado, M.E.; Malcata, F.X. Antioxidant activity of chitooligosaccharides upon two biological systems: Erythrocytes and bacteriophages. Carbohydr. Polym. 2010, 79, 1101–1106. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Sereno, J.; Garrido, P.; Parada, B.; Cunha, M.F.X.; Reis, F.; Pintado, M.E.; Santos-Silva, A. Inhibition of bladder tumor growth by chitooligosaccharides in an experimental carcinogenesis model. Mar. Drugs 2012, 10, 2661–2675. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.; Spindola, H.; De Sousa, V.; Santos-Silva, A.; Pintado, M.E.; Malcata, F.X.; Carvalho, J.E. Anti-inflammatory activity of chitooligosaccharides in vivo. Mar. Drugs 2010, 8, 1763–1768. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.C.; Tavaria, F.K.; Soares, J.C.; Ramos, O.S.; Joao Monteiro, M.; Pintado, M.E.; Xavier Malcata, F. Antimicrobial effects of chitosans and chitooligosaccharides, upon Staphylococcus aureus and Escherichia coli, in food model systems. Food Microbiol. 2008, 25, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Byun, H.-G.; Kim, Y.-T.; Park, P.-J.; Lin, X.; Kim, S.-K. Chitooligosaccharides as a novel b-secretase inhibitor. Carbohydr. Polym. 2005, 61, 198–202. [Google Scholar] [CrossRef]
- Du, Y.Z.; Lu, P.; Zhou, J.P.; Yuan, H.; Hu, F.Q. Stearic acid grafted chitosan oligosaccharide micelle as a promising vector for gene delivery system: Factors affecting the complexation. Int. J. Pharm. 2010, 391, 260–266. [Google Scholar] [CrossRef]
- Muanprasat, C.; Chatsudthipong, V. Chitosan oligosaccharide: Biological activities and potential therapeutic applications. Pharmacol. Ther. 2017, 170, 80–97. [Google Scholar] [CrossRef]
- Sun, T.; Zhou, D.; Xie, J.; Mao, F. Preparation of chitosan oligomers and their antioxidant activity. Eur. Food Res. Technol. 2007, 225, 451–456. [Google Scholar] [CrossRef]
- Sorlier, P.; Denuzière, A.; Viton, C.; Domard, A. Relation between the degree of acetylation and the electrostatic properties of chitin and chitosan. Biomacromolecules 2001, 2, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Tan, H.; Li, H.; You, C. Molecular weight dependence of structure and properties of chitosan oligomers. RSC Adv. 2015, 5, 69445–69452. [Google Scholar] [CrossRef]
- Raafat, D.; Sahl, H.G. Chitosan and its antimicrobial potential--a critical literature survey. Microb. Biotechnol. 2009, 2, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.G.; Koo, J.C.; Park, J.K. Antifungal Effect of Chitosan as Ca(2+) Channel Blocker. Plant. Pathol. J. 2016, 32, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Hamer, S.N.; Cord-Landwehr, S.; Biarnes, X.; Planas, A.; Waegeman, H.; Moerschbacher, B.M.; Kolkenbrock, S. Enzymatic production of defined chitosan oligomers with a specific pattern of acetylation using a combination of chitin oligosaccharide deacetylases. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Erdogan, S.; Kaya, M.; Akata, I. Chitin extraction and chitosan production from cell wall of two mushroom species (Lactarius vellereus and Phyllophora ribis). AIP Conf. Proc. 2017, 1809, 020012:1–020012:11. [Google Scholar]
- Ospina Alvarez, S.P.; Ramirez Cadavid, D.A.; Escobar Sierra, D.M.; Ossa Orozco, C.P.; Rojas Vahos, D.F.; Zapata Ocampo, P.; Atehortua, L. Comparison of extraction methods of chitin from Ganoderma lucidum mushroom obtained in submerged culture. Biomed. Res. Int. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ibitoye, E.B.; Lokman, I.H.; Hezmee, M.N.M.; Goh, Y.M.; Zuki, A.B.Z.; Jimoh, A.A. Extraction and physicochemical characterization of chitin and chitosan isolated from house cricket. Biomed. Mater. 2018, 13, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-W.; Han, Y.S.; Jo, Y.H.; Choi, M.H.; Kang, S.H.; Kim, S.-A.; Jung, W.-J. Extraction of chitin and chitosan from housefly, Musca domestica, pupa shells. Entomol. Res. 2016, 46, 324–328. [Google Scholar] [CrossRef]
- Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Zhu, F.; Qu, M.; Jiang, C.; Yang, Q. Extraction and characterization of chitin from the beetle Holotrichia parallela Motschulsky. Molecules 2012, 17, 4604–4611. [Google Scholar] [CrossRef] [PubMed]
- Kannan, M.; Nesakumari, M.; Rajarathinam, K.; Ranjit Singh, A.J.A. Production and characterization of mushroom chitosan under solid-state fermentation conditions. Adv. Biol. Res. 2010, 4, 10–13. [Google Scholar]
- Nitschke, J.; Altenbach, H.J.; Malolepszy, T.; Molleken, H. A new method for the quantification of chitin and chitosan in edible mushrooms. Carbohydr. Res. 2011, 346, 1307–1310. [Google Scholar] [CrossRef] [PubMed]
- Teng, W.L.; Khor, E.; Tan, T.K.; Lim, L.Y.; Tana, S.C. Concurrent production of chitin from shrimp shells and fungi. Carbohydr. Res. 2001, 332, 305–316. [Google Scholar] [CrossRef]
- Park, J.K. Algal Polysaccharides: Properties and Applications. Biochem. Anal. Biochem. 2015, 4, 1–2. [Google Scholar] [CrossRef]
- Rahman, M.A.; Halfar, J. First evidence of chitin in calcified coralline algae: New insights into the calcification process of Clathromorphum compactum. Sci. Rep. 2014, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.Y.; Huang, X.; Cheong, K.L. Recent Advances in Marine Algae Polysaccharides: Isolation, Structure, and Activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef] [PubMed]
- Abdulkarim, A.; Tijani Isa, M.; Abdulsalam, S.; Jaju Muhammad, A.; Opuada Ameh, A. Extraction and characterisation of chitin and chitosan from mussel shell. Civ. Environ. Res. 2013, 3, 108–115. [Google Scholar]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef]
- Mohammed, M.H.; Williams, P.A.; Tverezovskaya, O. Extraction of chitin from prawn shells and conversion to low molecular mass chitosan. Food Hydrocoll. 2013, 31, 166–171. [Google Scholar] [CrossRef] [Green Version]
- No, H.K.; Hur, E.Y. Control of foam formation by antifoam during demineralization of crustacean shell in preparation of chitin. J. Agric. Food Chem. 1998, 46, 3844–3846. [Google Scholar] [CrossRef]
- Tolaimate, A.; Desbrieres, J.; Rhazi, M.; Alagui, A. Contribution to the preparation of chitins and chitosans with controlled physico-chemical properties. Polymer 2003, 44, 7939–7952. [Google Scholar] [CrossRef]
- Bajaj, M.; Winter, J.; Gallert, C. Effect of deproteination and deacetylation conditions on viscosity of chitin and chitosan extracted from Crangon crangon shrimp waste. Biochem. Eng. J. 2011, 56, 51–62. [Google Scholar] [CrossRef]
- Gaikwad, B.V.; Koli, J.M.; Desai, A.S. Isolation and characterization of chitosan from crab (Scylla serrata)) shell waste. Int. J. Sci. Appl. Res. 2015, 2, 78–84. [Google Scholar]
- Percot, A.; Viton, C.; Domard, A. Characterization of shrimp shell deproteinization. Biomacromolecules 2003, 4, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Okafor, N. Isolation of chitin from the shell of the cuttlefish, Sepia oficirralis L. Biochim. Biophys. Acta 1965, 101, 193–200. [Google Scholar] [PubMed]
- Bajaj, M.; Freiberg, A.; Winter, J.; Xu, Y.; Gallert, C. Pilot-scale chitin extraction from shrimp shell waste by deproteination and decalcification with bacterial enrichment cultures. Appl. Microbiol. Biotechnol. 2015, 99, 9835–9846. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Dhillon, G.S. Recent trends in biological extraction of chitin from marine shell wastes: A review. Crit. Rev. Biotechnol. 2015, 35, 44–61. [Google Scholar] [CrossRef] [PubMed]
- Gortari, M.C.; Hours, R.A. Biotechnological processes for chitin recovery out of crustacean waste: A mini-review. Electron. J. Biotechnol. 2013, 16. [Google Scholar] [CrossRef]
- Younes, I.; Ghorbel-Bellaaj, O.; Nasri, R.; Chaabouni, M.; Rinaudo, M.; Nasri, M. Chitin and chitosan preparation from shrimp shells using optimized enzymatic deproteinization. Process. Biochem. 2012, 47, 2032–2039. [Google Scholar] [CrossRef]
- Lopes, C.; Antelo, L.T.; Franco-Uría, A.; Alonso, A.A.; Perez-Martín, R. Chitin production from crustacean biomass: Sustainability assessment of chemical and enzymatic processes. J. Clean. Prod. 2018, 172, 4140–4151. [Google Scholar] [CrossRef] [Green Version]
- Hajji, S.; Ghorbel-Bellaaj, O.; Younes, I.; Jellouli, K.; Nasri, M. Chitin extraction from crab shells by Bacillus bacteria. Biological activities of fermented crab supernatants. Int. J. Biol. Macromol. 2015, 79, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yun, S.; Song, L.; Zhang, Y.; Zhao, Y. The preparation and characterization of chitin and chitosan under large-scale submerged fermentation level using shrimp by-products as substrate. Int. J. Biol. Macromol. 2017, 96, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, J.A.; Rodriguez-Amado, I.; Montemayor, M.I.; Fraguas, J.; Gonzalez Mdel, P.; Murado, M.A. Chondroitin sulfate, hyaluronic acid and chitin/chitosan production using marine waste sources: Characteristics, applications and eco-friendly processes: A review. Mar. Drugs 2013, 11, 747–774. [Google Scholar] [CrossRef] [PubMed]
- Pires, C.T.G.V.M.T.; Vilela, J.A.P.; Airoldi, C. The Effect of Chitin Alkaline Deacetylation at Different Condition on Particle Properties. Procedia Chem. 2014, 9, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Tolaimatea, A.; Desbrieres, J.; Rhazi, M.; Alagui, A.; Vincendon, M.; Vottero, P. On the influence of deacetylation process on the physicochemical characteristics of chitosan from squid chitin. Polymer 2000, 41, 2463–2469. [Google Scholar] [CrossRef]
- Yuan, Y.; Chesnutt, B.M.; Haggard, W.O.; Bumgardner, J.D. Deacetylation of Chitosan: Material Characterization and in vitro Evaluation via Albumin Adsorption and Pre-Osteoblastic Cell Cultures. Materials 2011, 4, 1399–1416. [Google Scholar] [CrossRef] [Green Version]
- Hwang, K.T.; Jung, S.T.; Lee, G.D.; Chinnan, M.S.; Park, Y.S.; Park, H.J. Controlling molecular weight and degree of deacetylation of chitosan by rResponse surface methodology. J. Food Eng. 2007, 80, 749–753. [Google Scholar]
- Weska, R.F.; Moura, J.M.; Batista, L.M.; Rizzi, J.; Pinto, L.A.A. Optimization of deacetylation in the production of chitosan from shrimp wastes: Use of response surface methodology. J. Food Eng. 2007, 80, 749–753. [Google Scholar] [CrossRef]
- Trombotto, S.; Ladavière, C.; Delolme, F.; Domard, A. Chemical preparation and structural characterization of a homogeneous series of chitin/chitosan oligomers. Biomacromolecules 2008, 9, 1731–1738. [Google Scholar] [CrossRef]
- Chang, K.L.B.; Tai, M.C.; Cheng, F.H. Kinetics and products of the degradation of chitosan by hydrogen peroxide. J. Agric. Food Chem. 2001, 49, 4845–4851. [Google Scholar] [CrossRef] [PubMed]
- Aam, B.B.; Heggset, E.B.; Norberg, A.L.; Sørlie, M.; Vårum, K.M.; Eijsink, V.G.H. Production of chitooligosaccharides and their potential applications in medicine. Mar. Drugs 2010, 8, 1482–1517. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Rajapakse, N. Enzymatic production and biological activities of chitosan oligosaccharides (COS): A review. Carbohydr. Polym. 2005, 62, 357–368. [Google Scholar] [CrossRef]
- Halder, S.K.; Mondal, K.C. Microbial Valorization of Chitinous Bioresources for Chitin Extraction and Production of Chito-Oligomers and N-Acetylglucosamine: Trends, Perspectives and Prospects; Springer: Berlin, Germany, 2018; pp. 69–107. [Google Scholar]
- Cabrera, J.C.; Van Cutsem, P. Preparation of chitooligosaccharides with degree of polymerization higher than 6 by acid or enzymatic degradation of chitosan. Biochem. Eng. J. 2005, 25, 165–172. [Google Scholar] [CrossRef]
- Ibrahim, K.A.; El-Eswed, B.I.; Abu-Sbeih, K.A.; Arafat, T.A.; Al Omari, M.M.; Darras, F.H.; Badwan, A.A. Preparation of Chito-Oligomers by Hydrolysis of Chitosan in the Presence of Zeolite as Adsorbent. Mar. Drugs 2016, 14, 43. [Google Scholar] [CrossRef] [PubMed]
- Kazami, N.; Sakaguchi, M.; Mizutani, D.; Masuda, T.; Wakita, S.; Oyama, F.; Kawakita, M.; Sugahara, Y. A simple procedure for preparing chitin oligomers through acetone precipitation after hydrolysis in concentrated hydrochloric acid. Carbohydr. Polym. 2015, 132, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42(Database issue), D490–D495. [Google Scholar] [CrossRef]
- John, M.; Roehrig, H.; Schmidt, J.; Wieneke, U.; Schell, J. Rhizobium NodB protein involved in nodulation signal synthesis is a chitooligosaccharide deacetylase. Proc. Natl. Acad. Sci. USA 1993, 90, 625–629. [Google Scholar] [CrossRef]
- Araki, Y.; Ito, E. A pathway of chitosan formationin Mucor rouxii: Enzymatic deacetylaytion of chitin. Biochem. Biophys. Res. Commun. 1974, 56, 669–675. [Google Scholar] [CrossRef]
- Tsigos, I.; Martinou, A.; Kafetzopoulos, D.; Bouriotis, V. Chitin deacetylases: New, versatile tools in biotechnology. Trends Biotechnol. 2000, 18, 305–312. [Google Scholar] [CrossRef]
- Zhao, Y.; Park, R.D.; Muzzarelli, R.A. Chitin deacetylases: Properties and applications. Mar. Drugs 2010, 8, 24–46. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Martinez, A.; Grifoll-Romero, L.; Aragunde, H.; Sancho-Vaello, E.; Biarnes, X.; Lopez-Llorca, L.V.; Planas, A. Expression and specificity of a chitin deacetylase from the nematophagous fungus Pochonia chlamydosporia potentially involved in pathogenicity. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tsigos, I.; Zydowicz, N.; Martinou, A.; Domard, A.; Bouriotis, V. Mode of action of chitin deacetylase from Mucor rouxii on N-acetylchitooligosaccharides. Eur. J. Biochem. 1999, 261, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ju, W.-T.; Jo, G.-H.; Jung, W.-J.; Park, R.-D. Perspectives of chitin deacetylase research. Biotechnol. Biopolym. 2011, 131–145. [Google Scholar]
- Tokuyasu, K.; Mitsutomi, M.; Yamaguchi, I.; Hayashi, K.; Mori, Y. Recognition of chitooligosaccharides and their N-acetyl groups by putative subsites of chitin deacetylase from a deuteromycete Colletotrichum Lindemuthianum. Biochemistry 2000, 39, 8837–8843. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Cord-Landwehr, S.; Singh, R.; Bernard, F.; Kolkenbrock, S.; El Gueddari, N.E.; Moerschbacher, B.M. A Recombinant Fungal Chitin Deacetylase Produces Fully Defined Chitosan Oligomers with Novel Patterns of Acetylation. Appl. Environ. Microbiol. 2016, 82, 6645–6655. [Google Scholar] [CrossRef] [Green Version]
- Grifoll-Romero, L.; Pascual, S.; Aragunde, H.; Biarnés, X.; Planas, A. Chitin Deacetylases: Structures, Specificities, and Biotech Applications. Polymers 2018, 10, 352. [Google Scholar] [CrossRef]
- Hirano, T.; Uehara, R.; Shiraishi, H.; Hakamata, W.; Nishio, T. Chitin oligosaccharide deacetylase from Shewanella woodyi ATCC51908. J. Appl. Glycosci. 2015, 62, 153–157. [Google Scholar] [CrossRef]
- Hirano, T.; Shiraishi, H.; Ikejima, M.; Uehara, R.; Hakamata, W.; Nishio, T. Chitin oligosaccharide deacetylase from Shewanella baltica ATCC BAA-1091. Biosci. Biotechnol. Biochem. 2017, 81, 547–550. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.X.; Wang, X.; Roseman, S. The chitin catabolic cascade in the marine bacterium Vibrio cholerae: Characterization of a unique chitin oligosaccharide deacetylase. Glycobiology 2007, 17, 1377–1387. [Google Scholar] [CrossRef]
- Tuveng, T.R.; Rothweiler, U.; Udatha, G.; Vaaje-Kolstad, G.; Smalas, A.; Eijsink, V.G.H. Structure and function of a CE4 deacetylase isolated from a marine environment. PLoS ONE 2017, 12, e0187544. [Google Scholar] [CrossRef] [PubMed]
- Mishra, C.; Seminko, C.E.; Mccreath, K.J.; De la Vega, H.; Jones, B.J.; Specht, C.A.; Robbins, P.W. Cloning and expression of two chitin deacetylase genes of Saccharomyces Cerevisiae. Yeast 1997, 13, 327–336. [Google Scholar] [CrossRef]
- Martinou, A.; Koutsioulis, D.; Bouriotis, V. Expression, purification, and characterization of a cobalt-activated chitin deacetylase (Cda2p) from Saccharomyces cerevisiae. Protein Expr Purif. 2002, 24, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Martinou, A.; Koutsioulis, D.; Bouriotis, V. Cloning and expression of a chitin deacetylase gene (CDA2) from Saccharomyces cerevisiae in Escherichia coli. Enzym. Microb. Technol. 2003, 32, 757–763. [Google Scholar] [CrossRef]
- Yamada, M.; Kurano, M.; Inatomi, S.; Taguchi, G.; Okazaki, M.; Shimosaka, M. Isolation and characterization of a gene coding for chitin deacetylase specifically expressed during fruiting body development in the basidiomycete Flammulina velutipes and its expression in the yeast Pichia pastoris. FEMS Microbiol. Lett. 2008, 289, 130–137. [Google Scholar] [CrossRef]
- Gauthier, C.; Clerisse, F.; Dommes, J.; Jaspar-Versali, M.F. Characterization and cloning of chitin deacetylases from Rhizopus circinans. Protein Expr. Purif. 2008, 59, 127–137. [Google Scholar] [CrossRef]
- Hossbach, J.; Busswinkel, F.; Kranz, A.; Wattjes, J.; Cord-Landwehr, S.; Moerschbacher, B.M. A chitin deacetylase of Podospora anserina has two functional chitin binding domains and a unique mode of action. Carbohydr. Polym. 2018, 183, 1–10. [Google Scholar]
- Wang, Y.; Song, J.Z.; Yang, Q.; Liu, Z.H.; Huang, X.M.; Chen, Y. Cloning of a heat-stable chitin deacetylase gene from Aspergillus nidulans and its functional expression in Escherichia coli. Appl. Biochem. Biotechnol. 2010, 162, 843–854. [Google Scholar] [CrossRef]
- Cord-Landwehr, S.; Melcher, R.L.; Kolkenbrock, S.; Moerschbacher, B.M. A chitin deacetylase from the endophytic fungus Pestalotiopsis sp. efficiently inactivates the elicitor activity of chitin oligomers in rice cells. Sci. Rep. 2016, 6, 1–11. [Google Scholar]
- Hembach, L.; Cord-Landwehr, S.; Moerschbacher, B.M. Enzymatic production of all fourteen partially acetylated chitosan tetramers using different chitin deacetylases acting in forward or reverse mode. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Qin, Z.; Chen, Q.; Lin, S.; Luo, S.; Qiu, Y.; Zhao, L. Expression and characterization of a novel cold-adapted chitosanase suitable for chitooligosaccharides controllable preparation. Food Chem. 2018, 253, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Andres, E.; Albesa-Jove, D.; Biarnes, X.; Moerschbacher, B.M.; Guerin, M.E.; Planas, A. Structural basis of chitin oligosaccharide deacetylation. Angew. Chem. Int. Ed. Engl. 2014, 53, 6882–6887. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.S.; Ghormade, V.; Deshpande, M.V. Chitinolytic enzymes: An exploration. Enzym. Microb. Technol. 2000, 26, 473–483. [Google Scholar] [CrossRef]
- Liaqat, F.; Sozer Bahadir, P.; Elibol, M.; Eltem, R. Optimization of chitosanase production by Bacillus mojavensis EGE-B-5.2i. J. Basic Microbiol. 2018, 58, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Tews, I.; Perrakis, A.; Oppenheim, A.; Dauter, Z.; Wilson, K.S.; Vorgias, C.E. Bacterial chitobiase structure provides insight into catalytic mechanism and the basis of Tay-Sachs disease. Nat. Struct. Biol. 1996, 3, 638–648. [Google Scholar] [CrossRef]
- Aiba, S. Preparation of N-acetylchitooligosaccharides by lysozymic hydrolysates of partially N-acetylated chitosans. Carbohydr. Res. 1994, 261, 297–306. [Google Scholar] [CrossRef]
- Madhuprakash, J.; El Gueddari, N.E.; Moerschbacher, B.M.; Podile, A.R. Production of bioactive chitosan oligosaccharides using the hypertransglycosylating chitinase-D from Serratia proteamaculans. Bioresour. Technol. 2015, 198, 503–509. [Google Scholar] [CrossRef]
- Kuddus, S.M.; Ahmad, R.I.Z. Isolation of novel chitinolytic bacteria and production optimization of extracellular chitinase. J. Genet. Eng. Biotechnol. 2013, 11, 39–46. [Google Scholar]
- Nidheesh, T.; Pal, G.K.; Suresh, P.V. Chitooligomers preparation by chitosanase produced under solid state fermentation using shrimp by-products as substrate. Carbohydr. Polym 2015, 121, 1–9. [Google Scholar] [CrossRef]
- Krolicka, M.; Hinz, S.W.A.; Koetsier, M.J.; Joosten, R.; Eggink, G.; Van den Broek, L.A.M.; Boeriu, C.G. Chitinase Chi1 from Myceliophthora thermophila C1, a Thermostable Enzyme for Chitin and Chitosan Depolymerization. J. Agric. Food Chem. 2018, 66, 1658–1669. [Google Scholar] [CrossRef]
- Pechsrichuang, P.; Lorentzen, S.B.; Aam, B.B.; Tuveng, T.R.; Hamre, A.G.; Eijsink, V.G.H.; Yamabhai, M. Bioconversion of chitosan into chito-oligosaccharides (CHOS) using family 46 chitosanase from Bacillus subtilis (BsCsn46A). Carbohydr. Polym. 2018, 186, 420. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Jiang, S.; Ke, Z.M.; Wu, H.S.; Chi, C.W.; Guo, Z.Y. Recombinant expression of a chitosanase and its application in chitosan oligosaccharide production. Carbohydr. Res. 2009, 344, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Pechsrichuang, P.; Yoohat, K.; Yamabhai, M. Production of recombinant Bacillus subtilis chitosanase, suitable for biosynthesis of chitosan-oligosaccharides. Bioresour. Technol. 2013, 127, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Nampally, M.; Rajulu, M.B.; Gillet, D.; Suryanarayanan, T.S.; Moerschbacher, B.B. A High Diversity in Chitinolytic and Chitosanolytic Species and Enzymes and Their Oligomeric Products Exist in Soil with a History of Chitin and Chitosan Exposure. Biomed. Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohlhoff, M.; Niehues, A.; Wattjes, J.; Beneteau, J.; Cord-Landwehr, S.; El Gueddari, N.E.; Bernard, F.; Rivera-Rodriguez, G.R.; Moerschbacher, B.M. Chitinosanase: A fungal chitosan hydrolyzing enzyme with a new and unusually specific cleavage pattern. Carbohydr. Polym. 2017, 174, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Wattjes, J.; Niehues, A.; David, L.; Delair, T.; Moerschbacher, B.M. Enzymatic Production and Enzymatic-Mass Spectrometric Fingerprinting Analysis of Chitosan Polymers with Different Non-Random Patterns of Acetylation. J. Am. Chem. Soc. 2019, 141, 3137–3145. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fraga, B.; Da Silva, A.F.; Lopez-Seijas, J.; Sieiro, C. Optimized expression conditions for enhancing production of two recombinant chitinolytic enzymes from different prokaryote domains. Bioprocess. Biosyst. Eng. 2015, 38, 2477–2486. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, M.; Kitouni, M.; Carro, L.; Pujic, P.; Normand, P.; Boubakri, H. Chitinolytic actinobacteria isolated from an Algerian semi-arid soil: Development of an antifungal chitinase-dependent assay and GH18 chitinase gene identification. Ann. Microbiol. 2019, 69, 395–405. [Google Scholar] [CrossRef]
- Su, P.-C.; Hsueh, W.-C.; Chang, W.-S.; Chen, P.T. Enhancement of chitosanase secretion by Bacillus subtilis for production of chitosan oligosaccharides. J. Taiwan Inst. Chem. Eng. 2017, 79, 49–54. [Google Scholar] [CrossRef]
- Vaikuntapu, P.R.; Rambabu, S.; Madhuprakash, J.; Podile, A.R. A new chitinase-D from a plant growth promoting Serratia marcescens GPS5 for enzymatic conversion of chitin. Bioresour. Technol. 2016, 220, 200–207. [Google Scholar] [CrossRef]
- Gray, T.R.G.; Baxby, P. Chitin decomposition in soil: II. The ecology of chitinoclastic micro-organisms in forest soil. Trans. Br. Mycol. Soc. 1968, 51, 293–309. [Google Scholar] [CrossRef]
- Beier, S.; Bertilsson, S. Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 2013, 4, 149. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Aziz, N.; Choudhary, D.; Shrivastava, N.; Varma, A.; Paul, B. Identification of Chitin Degrading Bacterial Strains Isolated from Bulk and Rhizospheric Soil. J. Pure Appl. Microbiol. 2018, 12, 133–141. [Google Scholar] [CrossRef]
- Chen, J.K.; Shen, C.R.; Liu, C.L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef] [PubMed]
- Rasche, S.; Herwartz, D.; Schuster, F.; Jablonka, N.; Weber, A.; Fischer, R.; Schillberg, S. More for less: Improving the biomass yield of a pear cell suspension culture by design of experiments. Sci. Rep. 2016, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, N.; Schmitz, C.; Gromping, U.; Fischer, R.; Schillberg, S. Assessment of cultivation factors that affect biomass and geraniol production in transgenic tobacco cell suspension cultures. PLoS ONE 2014, 9, e104620. [Google Scholar] [CrossRef] [PubMed]
- Mandenius, C.-F.; Brundin, A. Review: Biocatalysts and bioreactor design. Biotechnol. Prog. 2008, 24, 1191–1203. [Google Scholar] [CrossRef]
- Kumar, S.; Mishra, A. Optimization of laccase production from WRF-1 on groundnut shell and cyanobacterial biomass: By application of Box-Behnken experimental design. J. Microbiol. Biotechnol. Res. 2011, 1, 33–53. [Google Scholar]
- Kumar, M.; Brar, A.; Vivekanand, V.; Pareek, N. Process optimization, purification and characterization of a novel acidic, thermostable chitinase from Humicola grisea. Int. J. Biol. Macromol. 2018, 116, 931–938. [Google Scholar] [CrossRef]
- Schmitz, C.; Fritsch, L.; Fischer, R.; Schillberg, S.; Rasche, S. Statistical experimental designs for the production of secondary metabolites in plant cell suspension cultures. Biotechnol. Lett. 2016, 38, 2007–2014. [Google Scholar] [CrossRef]
- Weissman, S.A.; Anderson, N.G. Design of Experiments (DoE) and Process Optimization. A Review of Recent Publications. Org. Process. Res. Dev. 2014, 19, 1605–1633. [Google Scholar]
- Buyel, J.F.; Fischer, R. Characterization of complex systems using the design of experiments approach: Transient protein expression in tobacco as a case study. J. Vis. Exp. 2014, 83, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, D.C. Design and Analysis of Experiments, 8th ed.; Wiley: Hoboken, NJ, USA, 2012; p. 752. [Google Scholar]
- Chylenski, P.; Forsberg, Z.; Stahlberg, J.; Varnai, A.; Lersch, M.; Bengtsson, O.; Saebo, S.; Horn, S.J.; Eijsink, V.G.H. Development of minimal enzyme cocktails for hydrolysis of sulfite-pulped lignocellulosic biomass. J. Biotechnol. 2017, 246, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, F.S.; Klopsch, R.; Oliviero, T.; Schreiner, M.; Verkerk, R.; Dekker, M. Optimizing isothiocyanate formation during enzymatic glucosinolate breakdown by adjusting pH value, temperature and dilution in Brassica vegetables and Arab. Thaliana. Sci. Rep. 2017, 7, 1–15. [Google Scholar]
- Babaki, M.; Yousefi, M.; Habibi, Z.; Mohammadi, M. Process optimization for biodiesel production from waste cooking oil using multi-enzyme systems through response surface methodology. Renew. Energy 2017, 105, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Mekasha, S.; Roksvåg Byman, I.; Lynch, C.; Toupalová, H.; Andera, L.; Næs, T.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Development of enzyme cocktails for complete saccharification ofchitin using mono-component enzymes from Serratia marcescens. Process. Biochem. 2017, 56, 132–138. [Google Scholar] [CrossRef]
- Kim, I.J.; Lee, H.J.; Kim, K.H. Pure enzyme cocktails tailored for the saccharification of sugarcane bagasse pretreated by using different methods. Process. Biochem. 2017, 57, 167–174. [Google Scholar] [CrossRef]
- Chen, J.-P.; Chang, K.-C. Immobilization of chitinase on a reversibly soluble-insoluble polymer for chitin hydrolysis. J.Chem. Tech. Biotechnol. 1994, 60, 133–140. [Google Scholar] [CrossRef]
- Datta, S.; Christena, L.R.; Rajaram, Y.R. Enzyme immobilization: An overview on techniques and support materials. 3 Biotech 2013, 3, 1–9. [Google Scholar] [CrossRef]
- Bloom, J.D.; Meyer, M.M.; Meinhold, P.; Otey, C.R.; MacMillan, D.; Arnold, F.H. Evolving strategies for enzyme engineering. Curr. Opin. Struct. Biol. 2005, 15, 447–452. [Google Scholar] [CrossRef]
- Chen, R. Enzyme engineering: Rational redesign versus directed evolution. Trends Biotechnol. 2001, 19, 13–14. [Google Scholar] [CrossRef]
- Newton, M.S.; Arcus, V.L.; Gerth, M.L.; Patrick, W.M. Enzyme evolution: Innovation is easy, optimization is complicated. Curr. Opin. Struct. Biol. 2018, 48, 110–116. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Source Organism | Expression Host | Substrate | COS Product | Literature |
|---|---|---|---|---|---|
| ClCDA | Colletotrichum lindemuthianum | Natural isolate | DP2–DP6 | Dn | [97] |
| MrCDA | Mucor rouxii | Natural isolate | DP3 | Dn | [95] |
| ScCDA1 | Saccharomyces cerevisiae | S. cerevisiae, Pichia pastoris | DP2–DP6 | n.r. | [104] |
| ScCDA2 | S. cerevisiae | S. cerevisiae, Pichia pastoris,E.coli | DP2–DP7 | n.r. | [105,106] |
| FvCDA | Flammulina velutipes | Pichia pastoris | DP2–DP6 | n.r. | [107] |
| RcCDA | Rhizopus circinans | Pichia pastoris | DP6 | n.r. | [108] |
| PaCDA | Podospora anserinas | Hansenula polymorpha | ≥DP2 | Dn | [109] |
| AnCDA | Aspergillus nidulans | E.coli | DP2–DP5 | Dn | [110] [114] |
| PgtCDA | Puccinia graminis | E.coli | DP4-DP6 | AADn-2 | [103] |
| PesCDA | Pestolotiopsis sp. | E.coli | DP4-DP6 | AADn-3A | [111] |
| PcCDA | Pochonia chlamydosporia | E.coli | DP4, DP5 | ADDAn-3 | [94] |
| SwCOD | Shewanella woodyi | E.coli | DP2–DP4 | AD; [ADAn−2] | [100] |
| SbCOD | Shewanella baltica | E.coli | DP2–DP4 | AD; [ADAn−2] | [99] |
| ArCE4A | Arthrobacter sp. | E.coli | DP2–DP6 | Dn−1 A | [102] |
| NodB | Rhizobium spp. | E.coli | DP1–DP6 | ADAn-2 | [48] |
| VcCOD | Vibrio chloreae | E.coli | DP2–DP6 | ADAn-2 | [110] |
| Enzyme | Source Organism | Expression Host | Substrate | COS Product | Literature |
|---|---|---|---|---|---|
| Chitinosanase | Alternaria alternata | Natural isolate | Chitosan DA 40–70% | Cleavage after GlcN-GlcNAc | [119] |
| Chitosanase | Purpureocillium lilacinum CFRNT12 | Natural isolate | Colloidal and crystalline chitosan | DP 2–6 | [120] |
| Chitinase Chi1 | Myceliophthora thermophila C1 | Myceliophthora thermophila C1 | Chitosan Mw (100, 600, and 3000 kDa); DA (77, 78, 88, 90%) | DP 2–12 | [121] |
| Chitosanase | B. subtilis | B. subtilis PT5 | α and β type chitosan | DP 2–4 | [122] |
| Chitinase-D | Serratia proteamaculans | Escherichia coli | Chitosan DA 35% and 61% | DP 2–12 | [123] |
| Chitinase-D | Serratia marcescens GPS5 | E. coli | Colloidal chitin, chitosan DA 10% | DP 1–8 | [124] |
| GH46 family chitosanase | Bacillus subtilis (BsCsn46A) | E. coli | Chitosan DA 15, 30 and 60% | DP 2–15 | [125,126] |
| GH46 family chitosanase | Gynuella sunshinyii | E. coli | Chitosan DA 95% COS DP 2–6 | DP 2–7 | [127] |
| GH8 family chitosanase | Bacillus strain | E. coli | Chitosan DA > 90% | DP 5.5 (mean) | [114] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmitz, C.; González Auza, L.; Koberidze, D.; Rasche, S.; Fischer, R.; Bortesi, L. Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Mar. Drugs 2019, 17, 452. https://doi.org/10.3390/md17080452
Schmitz C, González Auza L, Koberidze D, Rasche S, Fischer R, Bortesi L. Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Marine Drugs. 2019; 17(8):452. https://doi.org/10.3390/md17080452
Chicago/Turabian StyleSchmitz, Christian, Lilian González Auza, David Koberidze, Stefan Rasche, Rainer Fischer, and Luisa Bortesi. 2019. "Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects" Marine Drugs 17, no. 8: 452. https://doi.org/10.3390/md17080452
APA StyleSchmitz, C., González Auza, L., Koberidze, D., Rasche, S., Fischer, R., & Bortesi, L. (2019). Conversion of Chitin to Defined Chitosan Oligomers: Current Status and Future Prospects. Marine Drugs, 17(8), 452. https://doi.org/10.3390/md17080452