Anti-Hepatocellular Carcinoma (HepG2) Activities of Monoterpene Hydroxy Lactones Isolated from the Marine Microalga Tisochrysis Lutea



 , and
, and
Abstract
1. Introduction
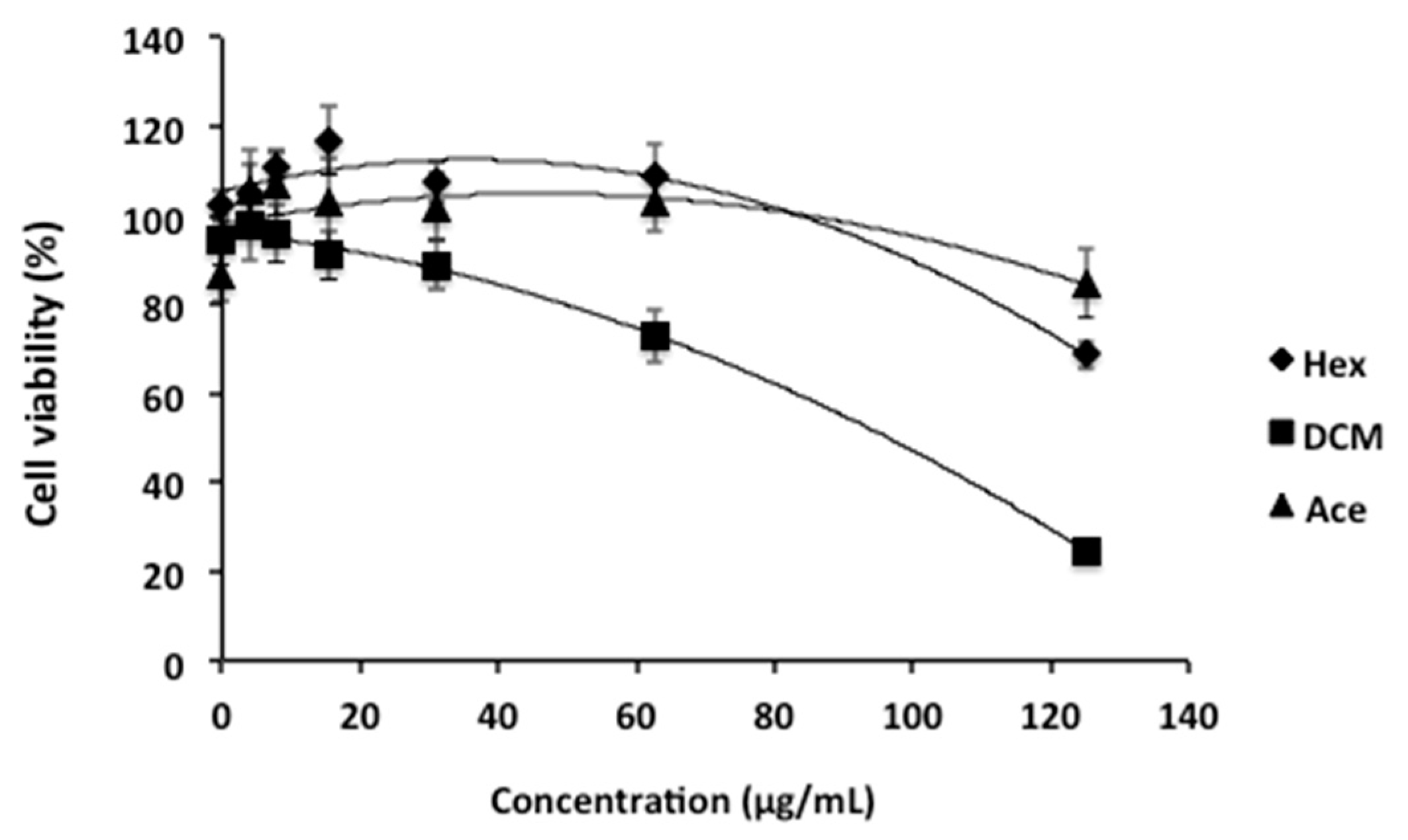
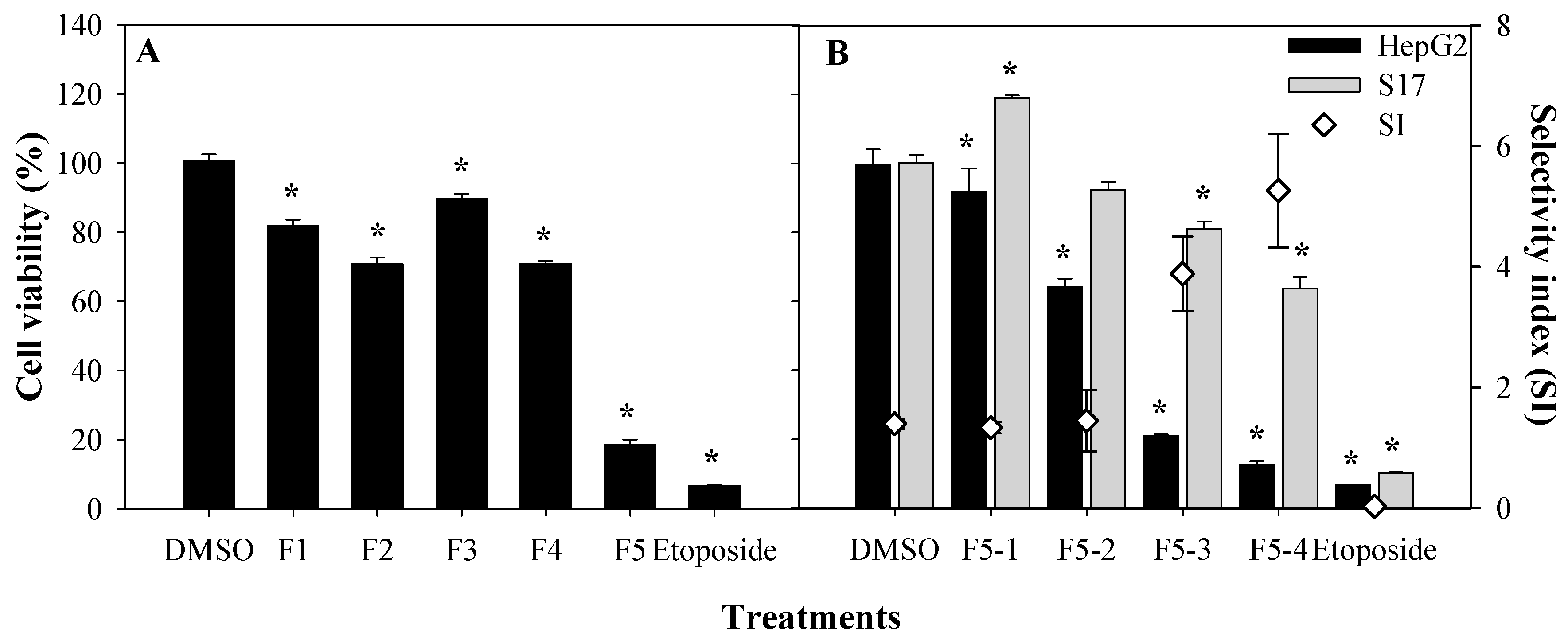
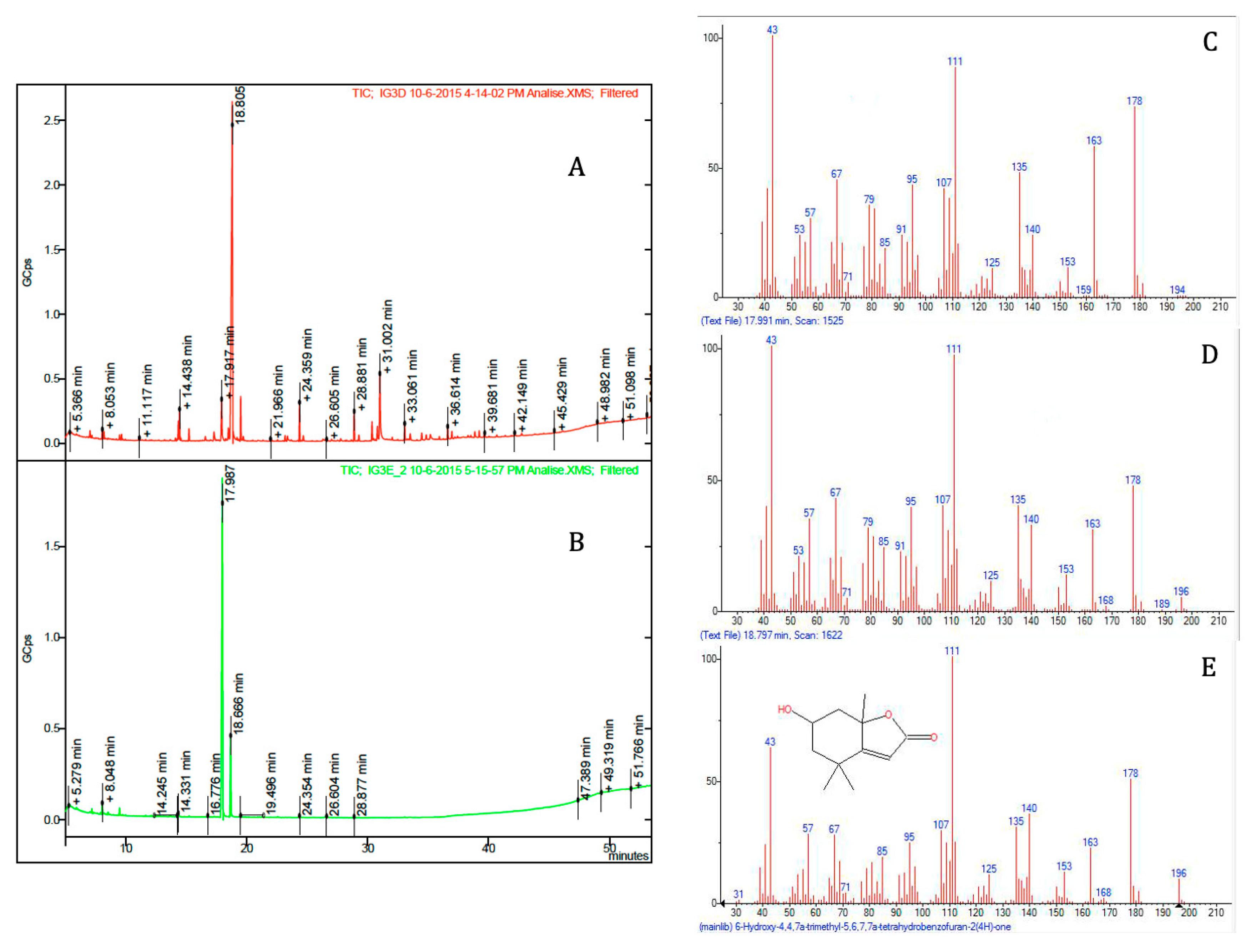
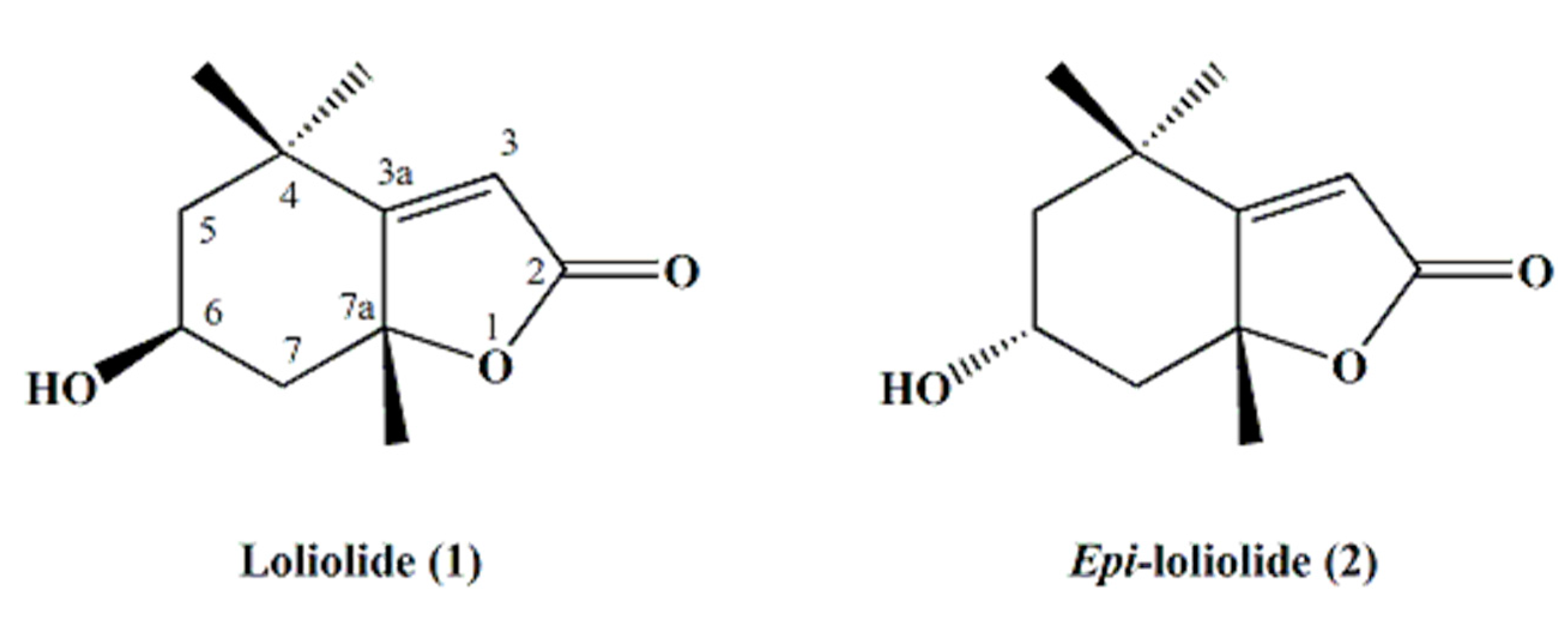
2. Results and Discussion
3. Materials and Methods
3.1. Materials
3.2. Extract Preparation from Microalgae
3.3. Cellular Viability
3.4. Bio-Guided Fractionation and Isolation of Anti-Tumoral Compounds
3.5. Spectral and Chromatographic Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilbert-López, B.; Mendiola, J.A.; Fontecha, J.; van den Broek, L.A.M.; Sijtsma, L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Downstream processing of Isochrysis galbana: A step towards microalgal biorefinery†. Green Chem. 2015, 17, 4599–4609. [Google Scholar] [CrossRef]
- De los Reyes, C.; Ortega, M.J.; Rodríguez-Luna, A.; Talero, E.; Motilva, V.; Zubía, E. Molecular characterization and anti-inflammatory activity of galactosylglycerides and galactosylceramides from the microalga Isochrysis galbana. J. Agric. Food Chem. 2016, 64, 8783–8794. [Google Scholar] [CrossRef] [PubMed]
- Bendif, E.M.; Probert, I.; Schroeder, D.C.; de Vargas, C. On the description of Tisochrysis lutea gen. nov. sp. nov. and Isochrysis nuda sp. nov. in the Isochrysidales, and the transfer of Dicrateria to the Prymnesiales (Haptophyta). J. Appl. Phycol. 2013, 25, 1763–1776. [Google Scholar] [CrossRef]
- Bigagli, E.; Cinci, L.; Niccolai, A.; Biondi, N.; Rodolfi, L.; D’Ottavio, M.; D’Ambrosio, M.; Lodovici, M.; Tredici, M.R.; Luceri, C. Preliminary data on the dietary safety, tolerability and effects on lipid metabolism of the marine microalga Tisochrysis lutea. Algal Res. 2018, 34, 244–249. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Delbrut, A.; Albina, P.; Lapierre, T.; Pradelles, R.; Dubreucq, E. Fucoxanthin and polyunsaturated fatty acids co-extraction by a green process. Molecules 2018, 23, 874. [Google Scholar] [CrossRef]
- Yu, C.C.; Chen, H.W.; Chen, M.J.; Chang, Y.C.; Chien, S.C.; Kuo, Y.H.; Chao, L.K.P. Chemical composition and bioactivities of the marine alga Isochrysis galbana CCMP 1324 from Taiwan. Nat. Prod. Commun. 2010, 5, 1941–1944. [Google Scholar] [CrossRef]
- Nuño, K.; Villarruel-Lopez, A.; Puebla-Perez, A.M.; Romero-Velarde, E.; Puebla-Mora, A.G.; Ascencio, F. Effects of the marine microalgae Isochrysis galbana and Nannochloropsis oculata in diabetic rats. J. Funct. Foods 2013, 5, 106–115. [Google Scholar] [CrossRef]
- Yingying, S.; Hui, W.; Ganlin, G.; Yinfang, P.; Binlun, Y. The isolation and antioxidant activity of polysaccharides from the marine microalgae Isochrysis galbana. Carbohydr. Polym. 2014, 113, 22–31. [Google Scholar] [CrossRef]
- Prakash, S.; Sasikala, S.L.; Aldous, V.H.J. Isolation and identification of MDR-Mycobacterium tuberculosis and screening of partially characterised antimycobacterial compounds from chosen marine micro algae. Asian Pac. J. Trop. Med. 2010, 3, 655–661. [Google Scholar] [CrossRef]
- Bonfanti, C.; Cardoso, C.; Afonso, C.; Matos, J.; Garcia, T.; Tanni, S.; Bandarra, N.M. Potential of microalga Isochrysis galbana: Bioactivity and bioaccessibility. Algal Res. 2018, 29, 242–248. [Google Scholar] [CrossRef]
- Sadovskaya, I.; Souissi, A.; Souissi, S.; Grad, T.; Lencel, P.; Greene, C.M.; Duin, S.; Dmitrenok, P.S.; Chizhov, A.O.; Shashkov, A.S.; et al. Chemical structure and biological activity of a highly branched (1→ 3, 1→ 6)-β-D-glucan from Isochrysis galbana. Carbohydr. Polym. 2014, 111, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Custódio, L.; Soares, F.; Pereira, H.; Barreira, L.; Vizetto-Duarte, C.; Rodrigues, M.J.; Amélia Pilar Rauter, A.M.; Alberício, F.; Varela, J. Fatty acid composition and biological activities of Isochrysis galbana T-ISO, Tetraselmis sp. and Scenedesmus sp.: Possible application in the pharmaceutical and functional food industries. J. Appl. Phycol. 2014, 26, 151–161. [Google Scholar] [CrossRef]
- Vizetto-Duarte, C.; Custódio, L.; Gangadhar, K.N.; Lago, J.H.G.; Dias, C.; Matos, A.M.; Rauter, A.P. Isololiolide, a carotenoid metabolite isolated from the brown alga Cystoseira tamariscifolia, is cytotoxic and able to induce apoptosis in hepatocarcinoma cells through caspase-3 activation, decreased Bcl-2 levels, increased p53 expression and PARP cleavage. Phytomedicine 2016, 23, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, Y.; Zhou, T.; Zheng, J.; Li, S.; Li, H.-B. Dietary natural products for prevention and treatment of liver cancer. Nutrients 2016, 8, 156. [Google Scholar] [CrossRef]
- Siddiqui, S.; Ahmad, R.; Khan, M.A.; Upadhyay, S.; Husain, I.; Srivastava, A.N. Cytostatic and anti-tumor potential of ajwa date pulp against human hepatocellular carcinoma HepG2 cells. Sci. Rep. 2019, 9, 245. [Google Scholar] [CrossRef]
- Andrade, K.A.M.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef]
- Xie, B.S.; Zhao, H.C.; Yao, S.K.; Zhuo, D.X.; Jin, B.; Lv, D.C.; Wu, C.L.; Dai-Long Ma, D.L.; Ai, Z.L. Autophagy inhibition enhances etoposide-induced cell death in human hepatoma G2 cells. Int. J. Mol. Med. 2011, 27, 599–606. [Google Scholar] [CrossRef]
- Dostál, Z.; Kosina, P.; Mlejnek, P.; Kikalová, K.; Modrianský, M. Mifepristone potentiates etoposide toxicity in Hep G2 cells by modulating drug transport. Toxicol. In Vitro 2019, 54, 33–40. [Google Scholar] [CrossRef]
- Oh, S.H.; Ahn, J.; Kang, D.H.; Lee, H.Y. The effect of ultrasonificated extracts of Spirulina maxima on the anticancer activity. Mar. Biotechnol. 2011, 13, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.J.; Gangadhar, K.N.; Vizetto-Duarte, C.; Wubshet, S.G.; Nyberg, N.T.; Barreira, L.; Varela, J.; Custódio, L. Maritime halophyte species from Southern Portugal as sources of bioactive molecules. Mar. Drugs 2014, 12, 2228–2244. [Google Scholar] [CrossRef] [PubMed]
- Mencher, S.K.; Wang, L.G. Promiscuous drugs compared to selective drugs (promiscuity can be a virtue). BMC Clin. Pharmacol. 2005, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Segun, P.A.; Ogbole, O.O.; Ismail, F.M.D.; Nahar, L.; Evans, A.R.; Ajaiyeoba, E.O.; Sarker, S.D. Resveratrol derivatives from Commiphora africana (A. Rich.) Endl. display cytotoxicity and selectivity against several human cancer cell lines. Phytother. Res. 2019, 33, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Percot, A.; Yalcin, A.; Aysel, V.; Erdugan, H.; Dural, B.; Guven, K.C. Loliolide in marine algae. Nat. Prod. Res. 2009, 23, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Qin, J.J.; Cheng, X.R.; Yan, S.K.; Jin, H.Z.; Zhang, W.D. Five new sesquiterpene lactones from Inula hupehensis. Arch. Pharm. Res. 2013, 36, 1319–1325. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Vanderah, D.J.; Hollenbeak, K.H.; Enwall, C.E.L.; Gopichand, Y.; SenGupta, P.K.; Van der Helm, D. Metabolites from the marine sponge Tedania ignis. A new atisanediol and several known diketopiperazines. J. Org. Chem. 1983, 48, 3941–3945. [Google Scholar] [CrossRef]
- Peng, Y.; Huang, R.-M.; Lin, X.-P.; Liu, Y.-H. Norisoprenoids from the brown alga Sargassum naozhouense Tseng et Lu. Molecules 2018, 23, 348. [Google Scholar] [CrossRef]
- Mori, K.; Khlebnikove, V. Carotenoids and Degraded Carotenoids, VIII – Synthesis of (+)-dihydroactinidiolide, (+)- and (−)-actinidiolide, (+)- and (−)-loliolide as well as (+)- and (−)-epiloliolide. Liebigs Ann. Chem. 1993, 1, 77–82. [Google Scholar] [CrossRef]
- Murata, M.; Nakai, Y.; Kawazu, K.; Ishizaka, M.; Kajiwara, H.; Abe, H.; Seo, S. Loliolide, a carotenoid metabolite, is a potential endogenous inducer of herbivore resistance. Plant Physiol. 2019, 179, 1822–1833. [Google Scholar] [CrossRef]
- Machida, K.; Kikuchi, M. Norisoprenoids from Viburnum dilatatum*. Phytochemistry 1996, 41, 1333–1336. [Google Scholar] [CrossRef]
- Ko, R.K.; Kang, M.C.; Kim, S.S.; Oh, T.H.; Kim, G.O.; Hyun, C.G.; Lee, N.H. Anti-melanogenesis constituents from the seaweed Dictyota coriacea. Nat. Prod. Commun. 2013, 8, 427–428. [Google Scholar] [CrossRef] [PubMed]
- Repeta, D.J. Carotenoid diagenesis in recent marine sediments: II. Degradation of fucoxanthin to loliolide*. Geochimica et Cosmochimica Acta 1989, 53, 669–707. [Google Scholar] [CrossRef]
- Taylor, H.F.; Burden, R.S. Identification of plant growth inhibitors produced by photolysis of violaxanthin. Phytochemistry 1970, 9, 2217–2223. [Google Scholar] [CrossRef]
- Schüler, L.M.; Gangadhar, K.N.; Duarte, P.; Placines, C.; Molina-Márquez, A.M.; Léon-Bañares, R.; Sousa, V.S.; Varela, J.; Barreira, L. Improvement of carotenoid extraction from a recently isolated, robust microalga, Tetraselmis sp. CTP4 (chlorophyta). Bioproc. Biosyst. Eng. 2020, 43, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Beuzenberg, V.; Goodwin, E.O.; Puddick, J.; Romanazzi, D.; Adams, S.L.; Packer, M.A. Optimising conditions for growth and xanthophyll production in continuous culture of Tisochrysis lutea using photobioreactor arrays and central composite design experiments. N. Z. J. Bot. 2017, 55, 64–78. [Google Scholar] [CrossRef]
- Boon, C.S.; McClements, D.J.; Weiss, J.; Decker, E.A. Factors influencing the chemical stability of carotenoids in foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef]
- Nonier, M.-F.; De Gaulejac, N.V.; Vivas, N.; Vitry, C. Characterization of carotenoids and their degradation products in oak wood. Incidence on the flavour of wood. Comptes Rendus Chim. 2004, 7, 689–698. [Google Scholar] [CrossRef]
- Mallik, A.K.; Qiu, H.; Sawada, T.; Takafuji, M.; Ihara, H. Molecular-shape selectivity by molecular gel-forming compounds: Bioactive and shape-constrained isomers through the integration and orientation of weak interaction sites†. Chem. Commun. 2011, 47, 10341–10343. [Google Scholar] [CrossRef]
- Okada, N.; Shirata, K.; Niwano, M.; Koshino, H.; Uramoto, M. Immunosuppressive activity of a monoterpene from Eucommia ulmoides. Int. J. Plant Biochem. 1994, 37, 281–282. [Google Scholar] [CrossRef]
- Kumar, J.; Dhar, P.; Tayade, A.B.; Gupta, D.; Chaurasia, O.P.; Upreti, D.K.; Srivastava, R.B. Chemical composition and biological activities of Trans-Himalayan alga Spirogyra porticalis (Muell.) cleve. PLOS ONE 2015, 10, e0118255. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Jeong, S.Y.; Jung, H.A.; Choi, J.S.; Min, B.S.; Woo, M.H. Anticholinesterase and antioxidant constituents from Gloiopeltis furcate. Chem. Pharm. Bull. 2010, 58, 1236–1239. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, D.S.; Kim, S.; Lorz, L.R.; Choi, E.; Lim, H.Y.; Yoon, K. Loliolide Presents Antiapoptosis and Antiscratching Effects in Human Keratinocytes. Int. J. Mol. Sci. 2019, 20, 651. [Google Scholar] [CrossRef]
- Islam, M.S.; Iwasaki, A.; Suenaga, K.; Kato-Noguchi, H. Isolation and identification of two potential phytotoxic substances from the aquatic fern Marsilea crenata. J. Plant Biol. 2017, 60, 75–81. [Google Scholar] [CrossRef]
- Takasaki, M.; Tokuda, H.; Nishino, H.; Konoshima, T. Cancer chemopreventive agents (antitumor-promoters) from Ajuga decumbens. J. Nat. Prod. 1999, 62, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Neergaard, J.S.; Rasmussen, H.B.; Stafford, G.I.; Van Staden, J.V.; Jäger, A.K. Serotonin transporter affinity of (−)-loliolide, a monoterpene lactone from Mondia whitei. S. Afr. J. Bot. 2010, 76, 593–596. [Google Scholar] [CrossRef][Green Version]
- Yang, H.H.; Hwangbo, K.; Zheng, M.S.; Cho, J.H.; Son, J.K.; Kim, H.Y.; Kim, J.R. Inhibitory effects of (2)-loliolide on cellular senescence in human dermal fibroblasts. Arch. Pharm. Res. 2015, 38, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.Y.; Liu, C.H.; Burnouf, T.; Wang, G.H.; Chang, S.P.; Jassey, A.; Yen, M.H. Activity-based and fraction-guided analysis of Phyllanthus urinaria identifies loliolide as a potent inhibitor of hepatitis C virus entry. Antivir. Res. 2016, 130, 58–68. [Google Scholar] [CrossRef]
- Lima, M.L.; Romanelli, M.M.; Borborema, S.E.T.; Johns, D.M.; Migotto, A.E.; Lago, J.H.G.; Tempone, A.G. Antitrypanosomal activity of isololiolide isolated from the marine hydroid Macrorhynchia philippina (Cnidaria, Hydrozoa). Bioorg. Chem. 2019, 89, 103002. [Google Scholar] [CrossRef]
- Machado, F.B.; Yamamoto, R.E.; Zanoli, K.; Nocchi, S.R.; Novello, C.R.; Schuquel, I.T.; Sakuragui, C.M.; Luftmann, H.; Nakamura, T.U.; Nakamura, C.V.; et al. Evaluation of the antiproliferative activity of the leaves from Arctium lappa by a bioassay-guided fractionation. Molecules 2012, 17, 1852–1859. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Loliolide (1) | Epi-loliolide (2) | ||
|---|---|---|---|---|
| δC | δH, m, J (Hz) | δC | δH, m, J (Hz) | |
| 2 | 171.49 | - | 171.55 | - |
| 3 | 112.89 | 5.69 s, 1H | 113.24 | 5.71 s, 1H |
| 3a | 182.68 | - | 180.75 | - |
| 4 | 35.91 | - | 35.03 | - |
| 5 | 47.26 | 1.53 dd, 14.5, 3.3, 1H, α-Hax | 49.7 | 1.33 t, 12.8, 1H, β-Hax |
| 1.97 brd, 14.6, 1H, β-Heq | 2.04 brd, 12.8, 1H, α-Heq | |||
| 6 | 66.81 | 4.33 m, 1H, α-Heq | 65.03 | 4.13 tt, 11.5, 4.1, 1H, α-Hax |
| 7 | 45.56 | 1.79 m, 1H, α-Hax | 47.25 | 1.51 t, 11.9, 1H, β-Hax |
| 2.45 brd, 14.1, 1H, β-Heq | 2.54 brd, 11.8, 1H, α-Heq | |||
| 7a | 86.67 | - | 86.45 | - |
| 4α-Me | 30.63 | 1.27 s, 3H, Meeq | 25.04 | 1.26 s, 3H, Meax |
| 4β-Me | 26.45 | 1.46 s, 3H, Meax | 29.89 | 1.31 s, 3H, Meeq |
| 7a-Me | 26.96 | 1.78 s, 3H, β-Meax | 25.55 | 1.58 s, 3H, α-Meax |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gangadhar, K.N.; Rodrigues, M.J.; Pereira, H.; Gaspar, H.; Malcata, F.X.; Barreira, L.; Varela, J. Anti-Hepatocellular Carcinoma (HepG2) Activities of Monoterpene Hydroxy Lactones Isolated from the Marine Microalga Tisochrysis Lutea. Mar. Drugs 2020, 18, 567. https://doi.org/10.3390/md18110567
Gangadhar KN, Rodrigues MJ, Pereira H, Gaspar H, Malcata FX, Barreira L, Varela J. Anti-Hepatocellular Carcinoma (HepG2) Activities of Monoterpene Hydroxy Lactones Isolated from the Marine Microalga Tisochrysis Lutea. Marine Drugs. 2020; 18(11):567. https://doi.org/10.3390/md18110567
Chicago/Turabian StyleGangadhar, Katkam N., Maria João Rodrigues, Hugo Pereira, Helena Gaspar, F. Xavier Malcata, Luísa Barreira, and João Varela. 2020. "Anti-Hepatocellular Carcinoma (HepG2) Activities of Monoterpene Hydroxy Lactones Isolated from the Marine Microalga Tisochrysis Lutea" Marine Drugs 18, no. 11: 567. https://doi.org/10.3390/md18110567
APA StyleGangadhar, K. N., Rodrigues, M. J., Pereira, H., Gaspar, H., Malcata, F. X., Barreira, L., & Varela, J. (2020). Anti-Hepatocellular Carcinoma (HepG2) Activities of Monoterpene Hydroxy Lactones Isolated from the Marine Microalga Tisochrysis Lutea. Marine Drugs, 18(11), 567. https://doi.org/10.3390/md18110567