Proteo-Transcriptomic Analysis Identifies Potential Novel Toxins Secreted by the Predatory, Prey-Piercing Ribbon Worm Amphiporus lactifloreus

 ,
,  , , ,
, , ,
Abstract
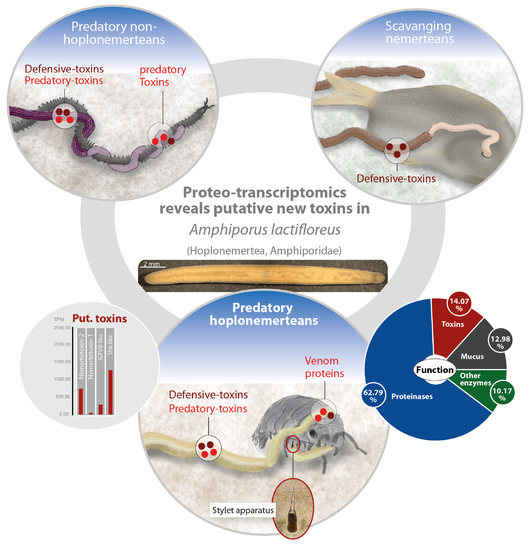
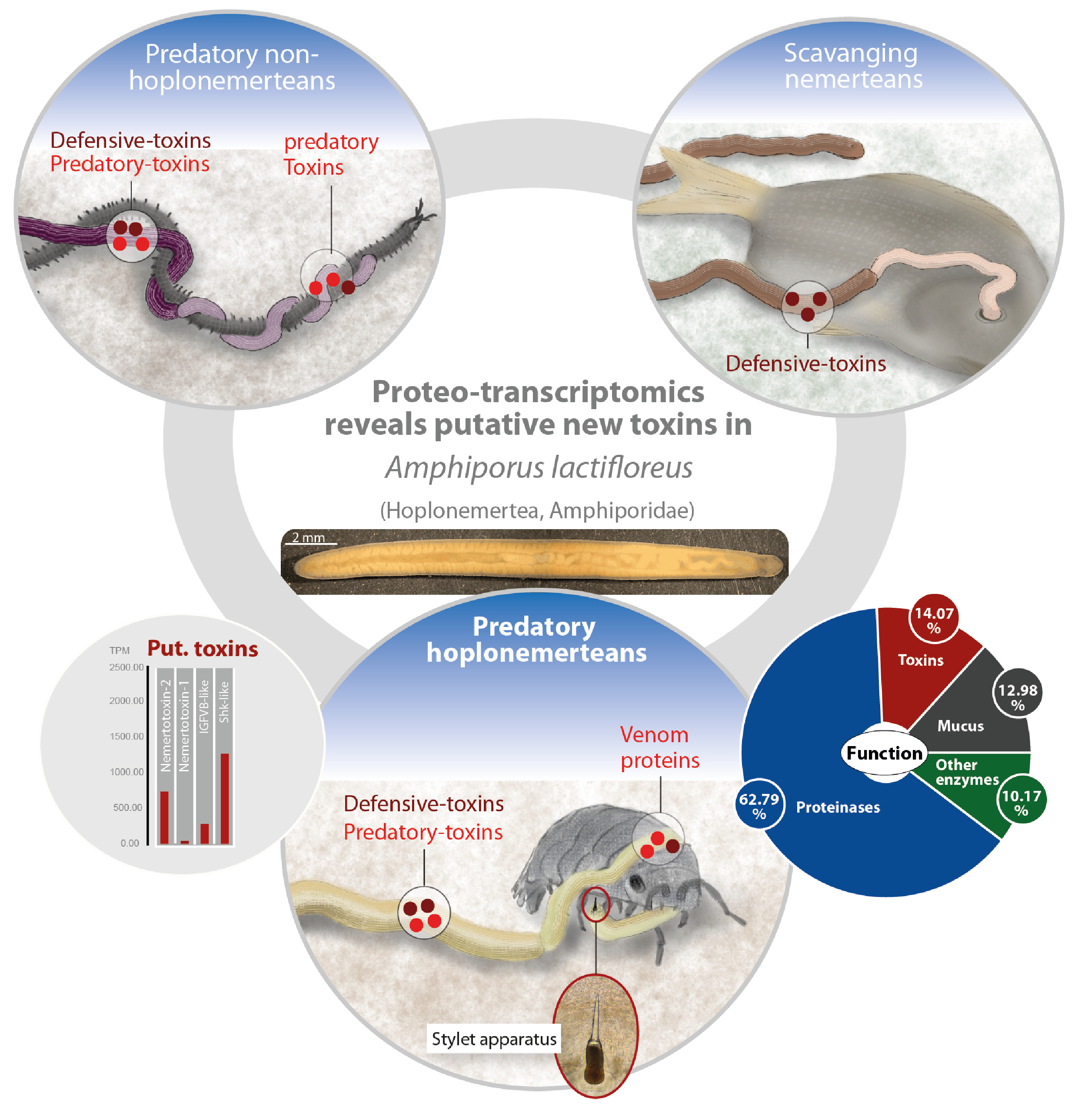
:
1. Introduction
2. Results
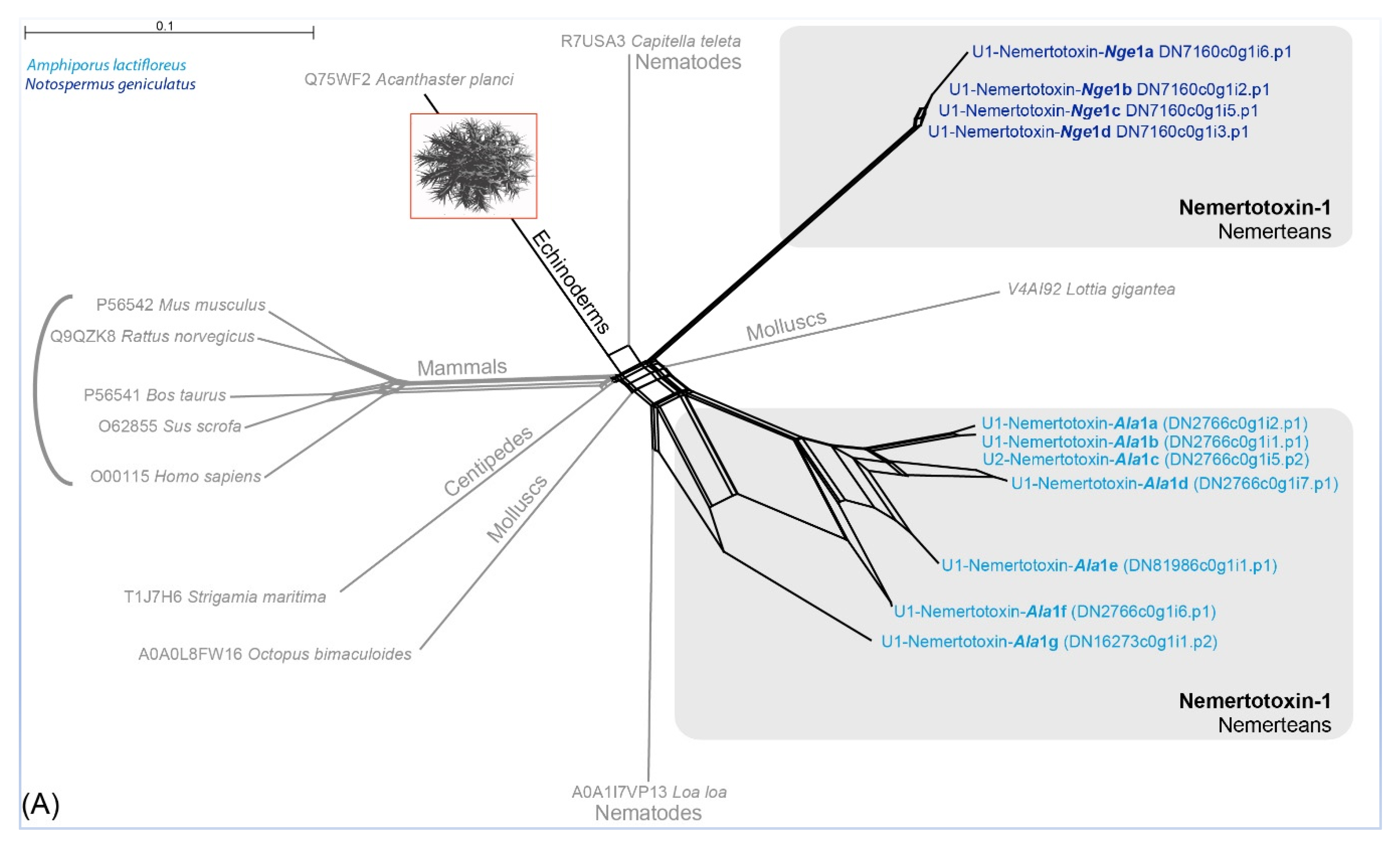
2.1. No Nemertean-Specific Toxin Transcripts Are Present in the A. lactifloreus Proboscis Transcriptome
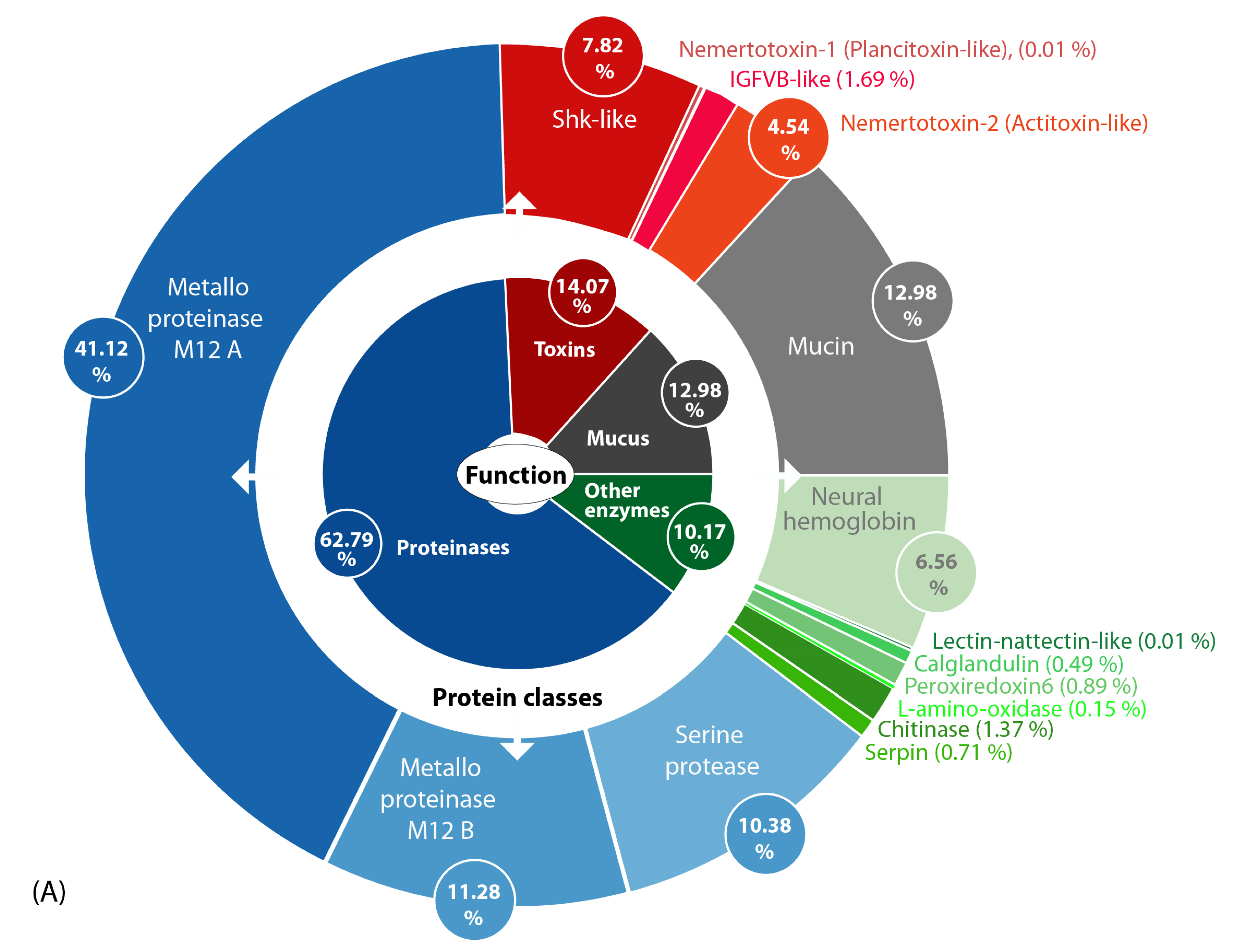
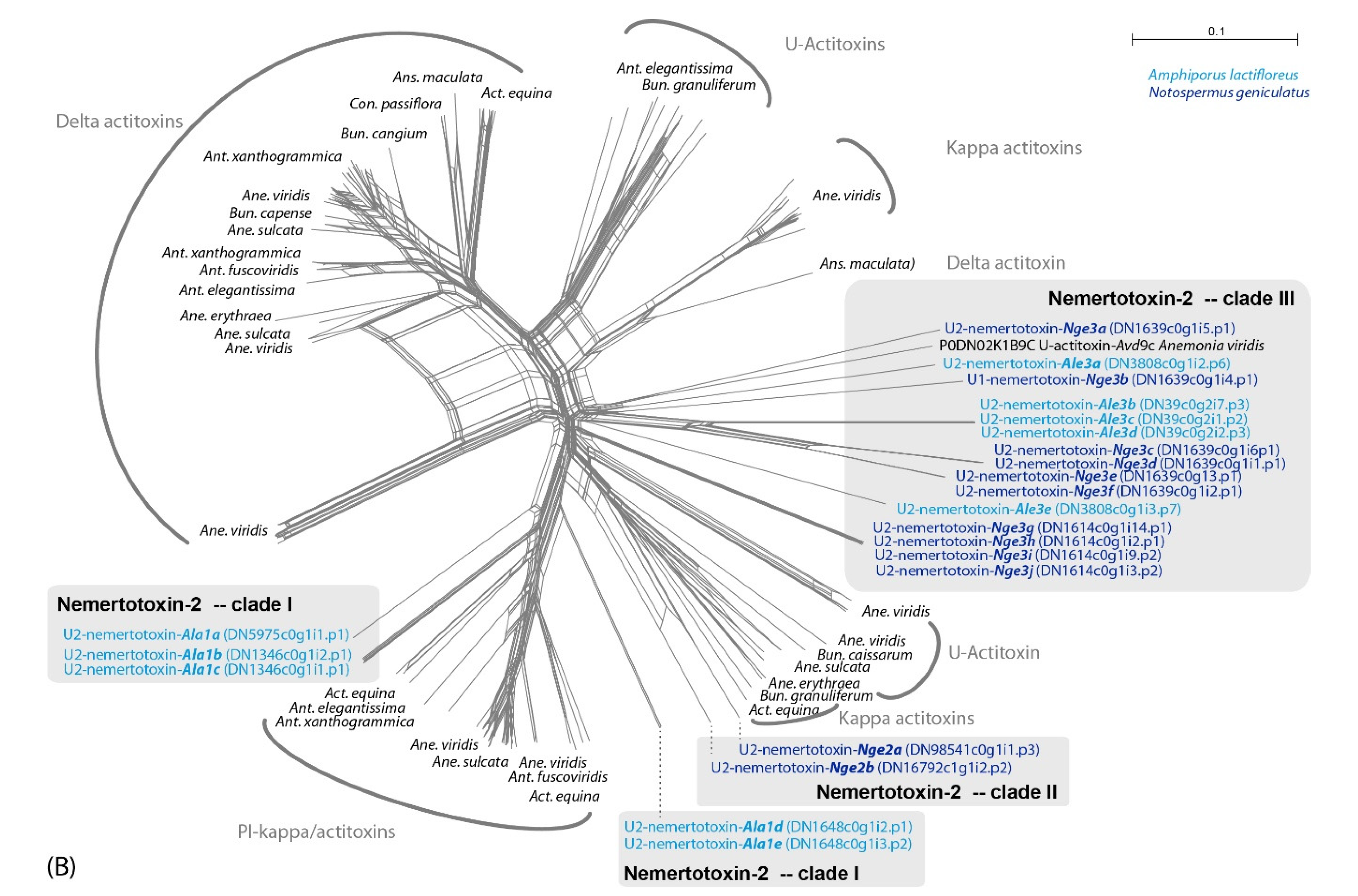
2.2. Metalloproteinase M12 and Actitoxin-Like Transcripts Are the Most Abundant Putative Toxin Transcripts in the Proboscis of A. lactifloreus
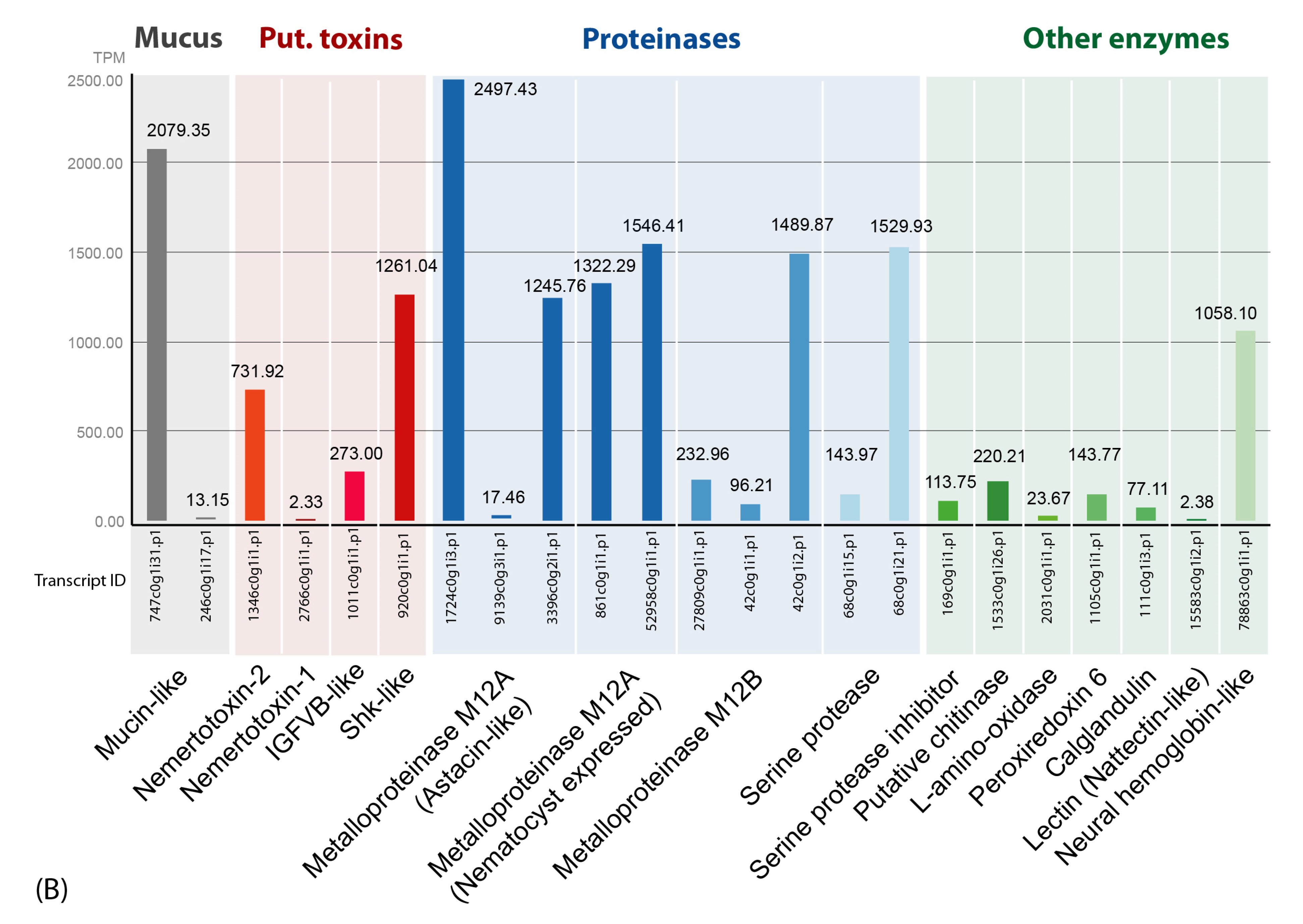
2.3. Proteinases Dominate the Skin and Mucus Secretions Accompanied by Mucins, Enzymes and Putative Toxins
2.4. Several Secreted Proteins Are Strongly Expressed but Remain Mostly Uncharacterized
2.5. No Putative Antimicrobial Peptides Were Identified in the A. lactifloreus Proteotranscriptome
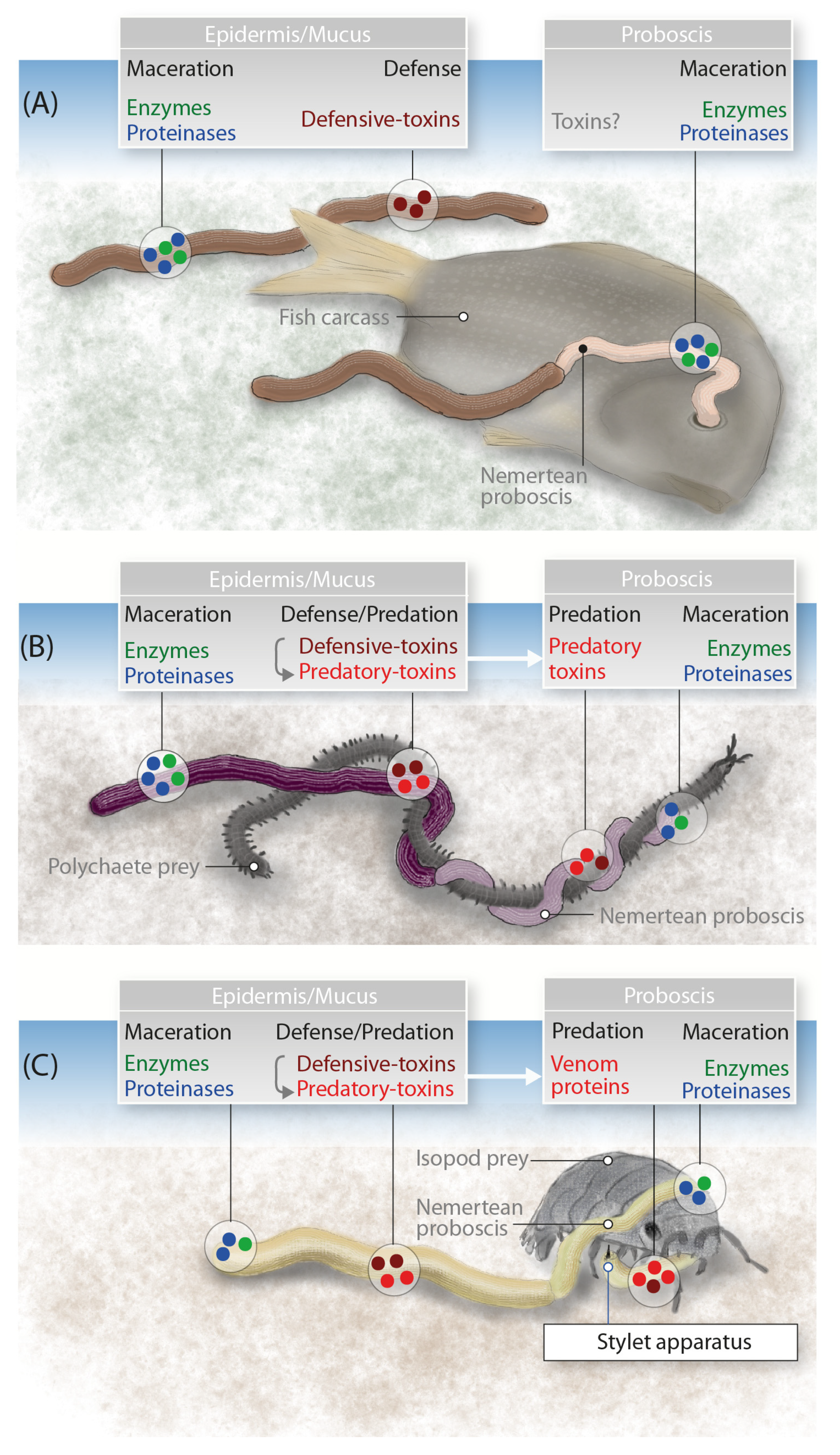
3. Discussion
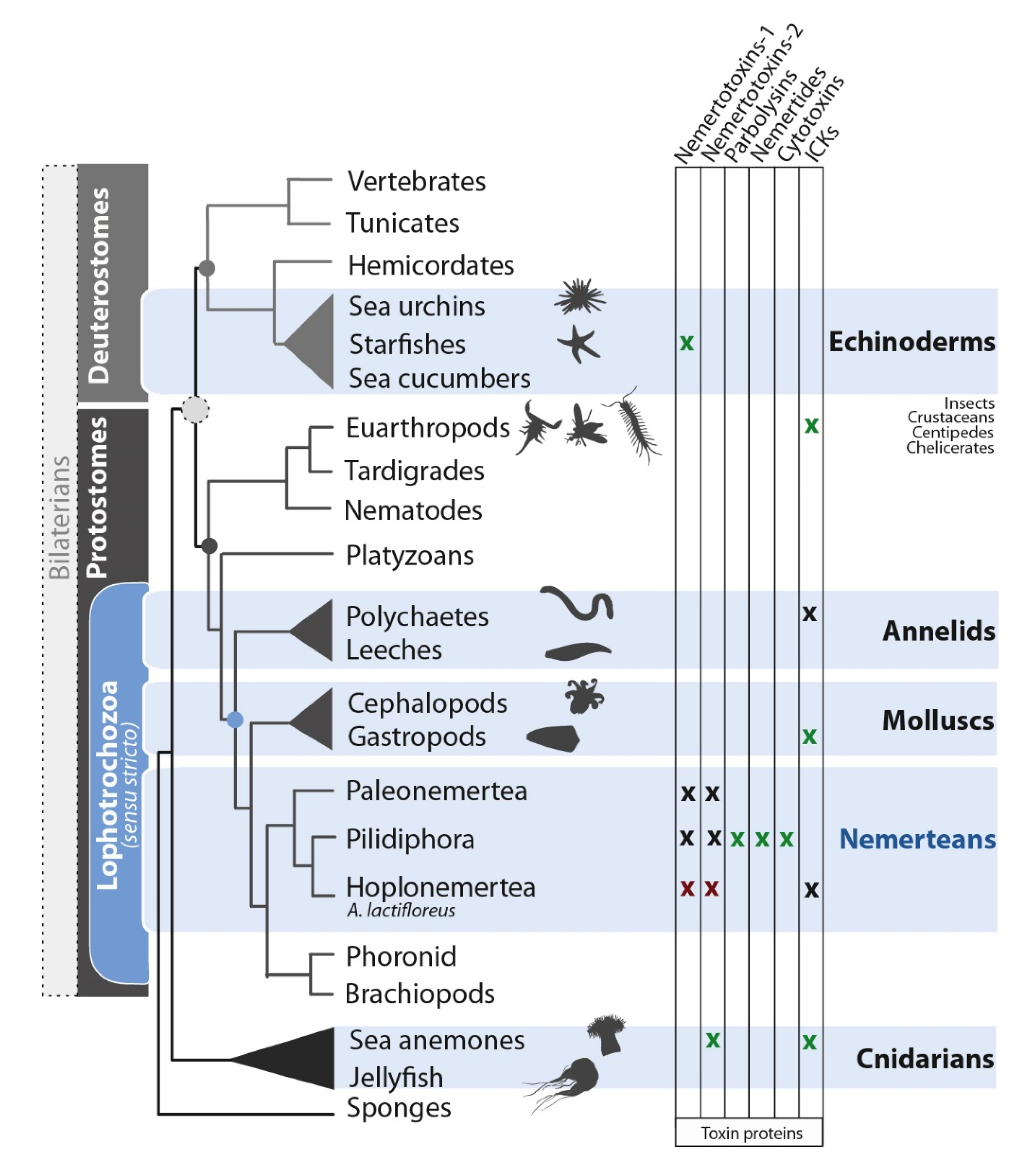
3.1. Are Known Nemertean Toxins Taxon-Specific?
3.2. The Putative Venom Cocktail of A. lactifloreus and Its Mode of Action
3.3. General Use of Toxin Proteins and Their Mode of Action in Nemerteans
4. Conclusions
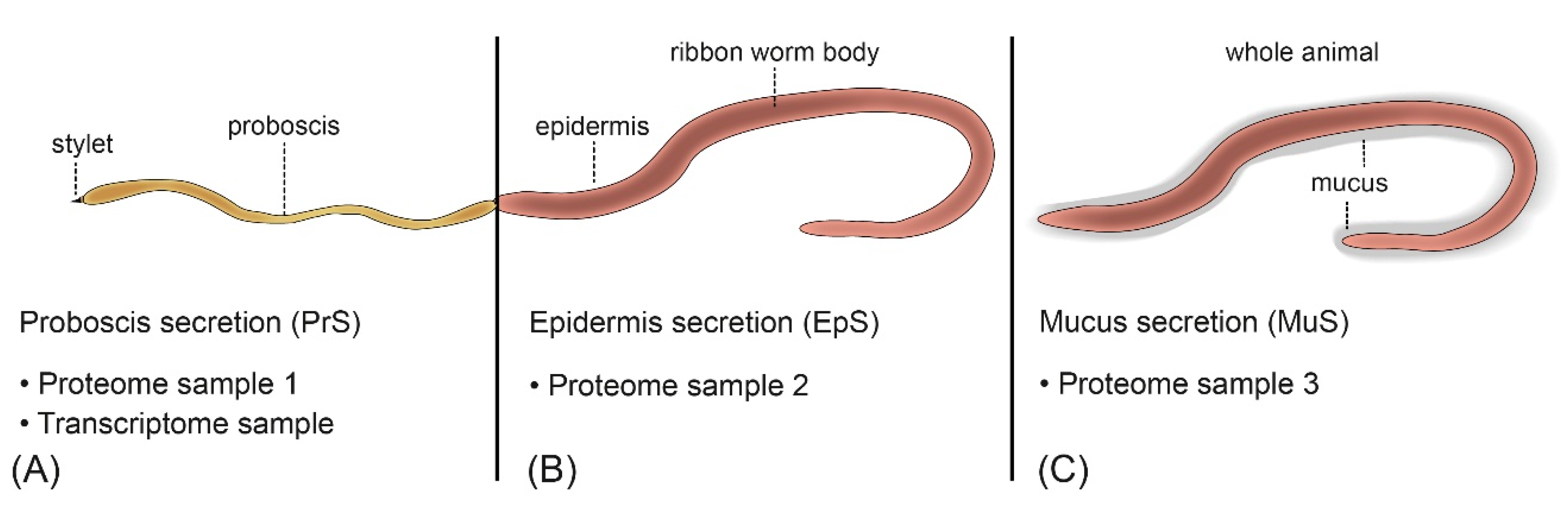
5. Materials and Methods
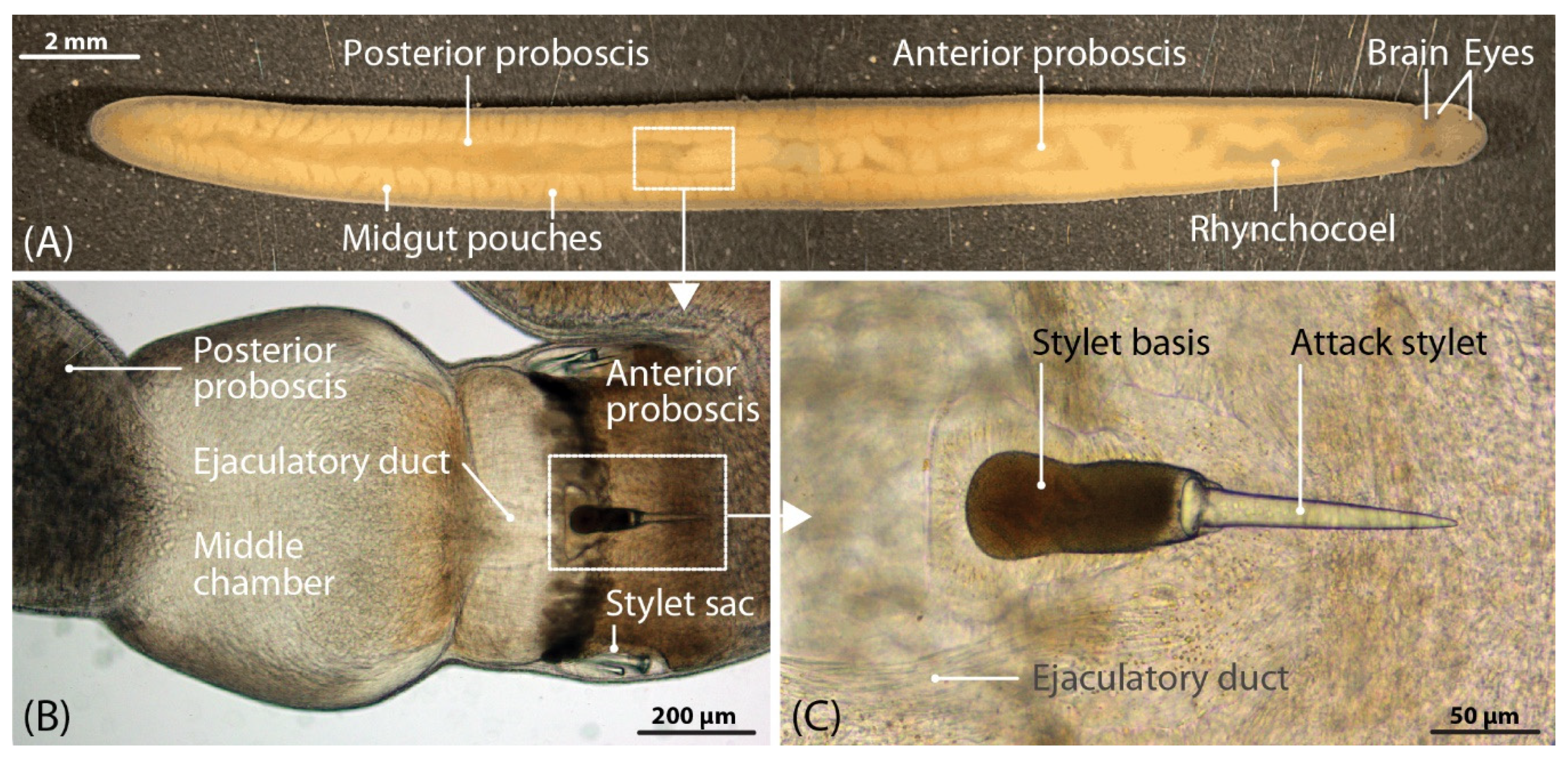
5.1. Collection and Preparation of A. lactifloreus Specimens
5.2. RNA Isolation, Library Preparation and Illumina Sequencing
5.3. Transcriptome Assembly, ORF Prediction and Identification of Venom Proteins
5.4. Peptide and Protein Identification
5.5. Matching Proteome and Transcriptome Data
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Appeltans, W.; Ahyong, S.T.; Anderson, G.; Angel, M.V.; Artois, T.; Bailly, N.; Bamber, R.; Barber, A.; Bartsch, I.; Berta, A.; et al. The Magnitude of Global Marine Species Diversity. Curr. Biol. 2012, 22, 2189–2202. [Google Scholar] [CrossRef]
- Göransson, U.; Jacobsson, E.; Strand, M.; Andersson, H. The Toxins of Nemertean Worms. Toxins 2019, 11, 120. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.-J.; Kanda, M.; Koyanagi, R.; Hisata, K.; Akiyama, T.; Sakamoto, H.; Sakamoto, T.; Satoh, N. Nemertean and phoronid genomes reveallophotrochozoan evolution and the origin of bilaterian heads. Nat. Ecol. Evol. 2018, 2, 1–14. [Google Scholar] [CrossRef]
- Strand, M.; Norenburg, J.; Alfaya, J.E.; Fernández-Álvarez, F.Á.; Andersson, H.S.; Andrade, S.C.S.; Bartolomaeus, T.; Beckers, P.; Bigatti, G.; Cherneva, I.; et al. Nemertean taxonomy—Implementing changes in the higher ranks, dismissing Anopla and Enopla. Zool. Scr. 2019, 48, 118–119. [Google Scholar] [CrossRef] [Green Version]
- McDermott, J.J. Observations on the food and feeding behavior of estuarine nemertean worms belonging to the order Hoplonemertea. Biol. Bull. 1976, 150, 57–68. [Google Scholar] [CrossRef]
- Thiel, M.; Kruse, I. Status of the Nemertea as predators in marine ecosystems. Hydrobiologia 2001, 456, 21–32. [Google Scholar] [CrossRef]
- Kruse, I.; Buhs, F. Preying at the edge of the sea: The nemertine Tetrastemma melanocephalum and its amphipod prey on high intertidal sandflats. In Life at Interfaces and under Extreme Conditions; Liebezeit, G., Dittmann, S., Kröncke, I., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 43–55. ISBN 978-94-010-5808-7. [Google Scholar]
- Mcdermott, J.J.; Roe, P. Food, Feeding Behavior and Feeding Ecology of Nemerteans. Am. Zool. 1985, 25, 113–125. [Google Scholar] [CrossRef]
- Beckers, P.; Bartolomaeus, T.; von Döhren, J. Observations and Experiments on the Biology and Life History of Riseriellus occultus (Heteronemertea: Lineidae). Zool. Sci. 2015, 32, 531–546. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Yanagihara, A.; Madio, B.; Nevalainen, T.; Alewood, P.; Fry, B. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [Green Version]
- Jouiaei, M.; Sunagar, K.; Federman Gross, A.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an ancient venom: Recognition of a novel family of cnidarian toxins and the common evolutionary origin of sodium and potassium neurotoxins in sea anemone. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- von Reumont, B.M.; Campbell, L.I.; Richter, S.; Hering, L.; Sykes, D.; Hetmank, J.; Jenner, R.A.; Bleidorn, C. A Polychaete’s powerful punch: Venom gland transcriptomics of Glycera reveals a complex cocktail of toxin homologs. Genome Biol. Evol. 2014, 6, 2406–2423. [Google Scholar] [CrossRef] [PubMed]
- Bon, C.; Saliou, B.; Thieffry, M.; Manaranche, R. Partial-Purification of Alpha-Glycerotoxin, a Presynaptic Neurotoxin From the Venom Glands of the Polychaete Annelid Glycera-Convoluta. Neurochem. Int. 1985, 7, 63–75. [Google Scholar] [CrossRef]
- Richter, S.; Helm, C.; Meunier, F.A.; Hering, L.; Campbell, L.I.; Drukewitz, S.H.; Undheim, E.A.B.; Jenner, R.A.; Schiavo, G.; Bleidorn, C. Comparative analyses of glycerotoxin expression unveil a novel structural organization of the bloodworm venom system. BMC Evol. Biol. 2017, 17, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stricker, S.A.; Cloney, R.A. The ultrastructure of venom-producing cells in Paranemertes peregrina (Nemertea, Hoplonemertea). J. Morphol. 1983, 177, 89–107. [Google Scholar] [CrossRef]
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.K.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. 2014, 89, 450–465. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Whelan, N.V.; Kocot, K.M.; Santos, S.R.; Halanych, K.M. Nemertean Toxin Genes Revealed through Transcriptome Sequencing. Genome Biol. Evol. 2014, 6, 3314–3325. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.J.; Undheim, E.A.B. True Lies: Using Proteomics to Assess the Accuracy of Transcriptome-Based Venomics in Centipedes Uncovers False Positives and Reveals Startling Intraspecific Variation in Scolopendra Subspinipes. Toxins 2018, 10, 96. [Google Scholar] [CrossRef] [Green Version]
- von Reumont, B.M. Studying Smaller and Neglected Organisms in Modern Evolutionary Venomics Implementing RNASeq (Transcriptomics)—A Critical Guide. Toxins 2018, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, C.; Dovell, S.; Melaun, C.; Marí, F. Definition of the R-superfamily of conotoxins: Structural convergence of helix-loop-helix peptidic scaffolds. Peptides 2018, 107, 75–82. [Google Scholar] [CrossRef] [PubMed]
- von Reumont, B.M.; Undheim, E.A.B.; Jauss, R.-T.; Jenner, R.A. Venomics of Remipede Crustaceans Reveals Novel Peptide Diversity and Illuminates the Venom’s Biological Role. Toxins 2017, 9, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Reumont, B.M.; Blanke, A.; Richter, S.; Alvarez, F.; Bleidorn, C.; Jenner, R.A. The first venomous crustacean revealed by transcriptomics and functional morphology: Remipede venom glands express a unique toxin cocktail dominated by enzymes and a neurotoxin. Mol. Biol. Evol. 2014, 31, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drukewitz, S.H.; von Reumont, B.M. The Significance of Comparative Genomics in Modern Evolutionary Venomics. Front. Ecol. Evol. 2019, 7, 163. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [Green Version]
- Dubaissi, E.; Rousseau, K.; Hughes, G.W.; Ridley, C.; Grencis, R.K.; Roberts, I.S.; Thornton, D.J. Functional characterization of the mucus barrier on the Xenopus tropicalis skin surface. Proc. Natl. Acad. Sci. USA 2018, 115, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Romero-Gutiérrez, M.T.; Santibáñez-López, C.E.; Jiménez-Vargas, J.M.; Batista, C.V.F.; Ortiz, E.; Possani, L.D. Transcriptomic and Proteomic Analyses Reveal the Diversity of Venom Components from the Vaejovid Scorpion Serradigitus gertschi. Toxins 2018, 10, 359. [Google Scholar] [CrossRef] [Green Version]
- Shiomi, K.; Midorikawa, S.; Ishida, M.; Nagashima, Y.; Nagai, H. Plancitoxins, lethal factors from the crown-of-thorns starfish Acanthaster planci, are deoxyribonucleases II. Toxicon 2004, 44, 499–506. [Google Scholar] [CrossRef]
- Shiomi, K.; Itoh, K.; Yamanaka, H.; Kikuchi, T. Biological activity of crude venom from the crown-of-thorns starfish Acanthaster planci. NIPPON SUISAN GAKKAISHI 1985, 51, 1151–1154. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.S.; Fuentes-Silva, D.; King, G.F. Development of a rational nomenclature for naming peptide and protein toxins from sea anemones. Toxicon 2012, 60, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; King, G.F.; Undheim, E.A.B. Sea Anemone Toxins: A Structural Overview. Mar. Drugs 2019, 17, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Undheim, E.A.B.; Fry, B.G.; King, G.F. Centipede venom: Recent discoveries and current state of knowledge. Toxins 2015, 7, 679–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Undheim, E.A.B.; Jones, A.; Clauser, K.R.; Holland, J.W.; Pineda, S.S.; King, G.F.; Fry, B.G. Clawing through evolution: Toxin diversification and convergence in the ancient lineage Chilopoda (centipedes). Mol. Biol. Evol. 2014, 31, 2124–2148. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Li, C.; Haug, T.; Styrvold, O.B.; Jørgensen, T.Ø.; Stensvåg, K. Strongylocins, novel antimicrobial peptides from the green sea urchin, Strongylocentrotus droebachiensis. Dev. Comp. Immunol. 2008, 32, 1430–1440. [Google Scholar] [CrossRef]
- Grafskaia, E.N.; Polina, N.F.; Babenko, V.V.; Kharlampieva, D.D.; Bobrovsky, P.A.; Manuvera, V.A.; Farafonova, T.E.; Anikanov, N.A.; Lazarev, V.N. Discovery of novel antimicrobial peptides: A transcriptomic study of the sea anemone Cnidopus japonicus. J. Bioinform. Comput. Biol. 2018, 16, 1840006. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.A.; Madio, B.; Jin, J.; Undheim, E.A.B.; Fry, B.G.; King, G.F. Melt with This Kiss: Paralyzing and Liquefying Venom of the Assassin Bug Pristhesancus plagipennis (Hemiptera: Reduviidae). Mol. Cell. Proteom. MCP 2017, 16, 552–566. [Google Scholar] [CrossRef] [Green Version]
- Drukewitz, S.H.; Fuhrmann, N.; Undheim, E.A.B.; Blanke, A.; Giribaldi, J.; Mary, R.; Laconde, G.; Dutertre, S.; von Reumont, B.M. A Dipteran’s Novel Sucker Punch: Evolution of Arthropod Atypical Venom with a Neurotoxic Component in Robber Flies (Asilidae, Diptera). Toxins 2018, 10, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raaymakers, C.; Verbrugghe, E.; Hernot, S.; Hellebuyck, T.; Betti, C.; Peleman, C.; Claeys, M.; Bert, W.; Caveliers, V.; Ballet, S.; et al. Antimicrobial peptides in frog poisons constitute a molecular toxin delivery system against predators. Nat. Commun. 2017, 8, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 11 June 2020).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ToxProt Annotation | Protein Family, “Actual” Scaffold | Original Taxon Source | Presumed Activity | Transcripts Included/(All) | Expression TPMs (Sum) |
|---|---|---|---|---|---|
| Toxin Candidates | |||||
| Actitoxin-like* (Nemertotoxin 2) | Actitoxins, Kunitz-BPTI | Sea anemones | Possible neurotoxicity | 10/(32) | 1052.76 |
| Delta-actitoxin-like | Actitoxins, Kunitz-BPTI | Sea anemones | Possible neurotoxicity | 2/(2) | 134.56 |
| Conotoxin-like | Conotoxin-like ICK 4-C scaffold | Mollusks | Possible neurotoxicity | 1/(3) | 19.74 |
| Conotoxin-like | Xibalbin1-like ICK 8-C scaffold | Mollusks | Possible neurotoxicity | 2/(3) | 19.74 |
| Plancitoxin-like* (Nemertotoxin 1) | DNase II | Starfishes | Possible hepatotoxicity | 7/(7) | 29.17 |
| Putative calcium channel toxin Tx758-like | ICK, knottin cysteine rich | Scorpions | Possible neurotoxicity | 1/(1) | 85.10 |
| Kunitz-type U1 aranetoxin-like | Kunitz-BPTI, cysteine-rich | Spiders | Possible neurotoxicity | 1/(1) | 9.42 |
| Proteinase Candidates | |||||
| Astacin-like metalloproteinase* | Metalloproteinase M12A | Diverse species | Proteinase activity | 8/(52) | 2597.43 |
| Nematocyst expressed protein 6 | Metalloproteinase M12A | Cnidarians | Proteinase activity | 1/(36) | 29.92 |
| Metalloproteinase zinc-disintegrin | Metalloproteinase ADAM | Diverse species | Proteinase activity | 1/(23) | 0.07 |
| Enzyme Candidates | |||||
| Venom allergen 5 | CAP, Cysteine-rich | Diverse species | Enzymatic | 1/(5) | 1.76 |
| C-type lectin (lectotoxin) | Lectins, C-type lectin | Diverse species | Enzymatic | 3/(14) | 5.5 |
| Galactose-specific lectin* (nattectin) | Lectins, C-type lectin | Diverse species | Enzymatic | 2/(11) | 3.20 |
| Calglandulin* | Calglandulins, EF-hand motif | Diverse species | Enzymatic | 3/(75) | 8.58 |
| Cysteine-rich protein | Diverse, cysteine-rich | Diverse species | Enzymatic | 2/(25) | 16.89 |
| Hyaluronidase (Conhyal-Cn1) | Hyaluronidase | Diverse species | Enzymatic | 6/(9) | 10.68 |
| Kunitz-type serine proteinase inhibitor | Serpin | Diverse species | Enzymatic | 3/(3) | 20.81 |
| Phospholipase A2 | Phospholipase A2 | Diverse species | Enzymatic | 4/(6) | 713.59 |
| Snaclec bitiscetin | Lectins | Diverse species | Enzymatic | 4/(5) | 4.38 |
| Snaclec coagulation factor | Lectins | Diverse species | Enzymatic | 3/(4) | 3.05 |
| Snake venom metalloproteinase inhibitor | SVMP | Diverse species | Proteinase activity | 3/(7) | 8.15 |
| Snake venom 5 nucleotidase | 5-Nuclease | Snakes | Proteinase activity | 3/(4) | 15.43 |
| Other Protein Candidates | |||||
| Vascular endothelial growth factor | Growth factors | Diverse species | Possible spreading factor | 1/(2) | 4.42 |
| Venom nerve growth factor | Growth factors | Diverse species | Possible spreading factor | 1/(2) | 10.16 |
| Insulin-like growth factor (IGFVB) | Growth factors | Diverse species | Possible spreading factor | 3/(49) | 3 |
| Neuropeptide prohormone-4 | Hormone precursor | Diverse species | Possible spreading factor | 2/(7) | 155.35 |
| Transcript | TPM Value | Mascot Score | Length (aa) | Signal Peptide | Scaffold/Domain Prediction (Sequence Residue) |
|---|---|---|---|---|---|
| DN187_c0_g1_i14.p1 | 8524.05 | 195 | 161 | yes (1–17) | Non-cytoplasmic domain (18–160), disorder prediction (137–160) |
| DN66444_c0_g1_i1.p1 | 2276.01 | 117 | 144 | yes (1–19) | Non-cytoplasmic domain (20–144) |
| DN2243_c0_g1_i6.p1 | 2243.13 | 104 | 158 | yes (1–22) | Non-cytoplasmic domain (23–158) |
| DN904_c0_g1_i1.p1 | 1328.59 | 177 | 100 | yes (1–18) | Non-cytoplasmic domain (19–100) |
| DN416_c0_g1_i1.p1 | 1173.91 | 70 | 198 | yes (1–19) | Non-cytoplasmic domain (20–198) |
| DN2192_c0_g1_i1.p1 | 709.61 | 114 | 117 | yes (1–20) | Non-cytoplasmic domain (21–116) |
| DN4062_c0_g1_i1.p1 | 558.79 | 60 | 210 | yes (1–23) | Non-cytoplasmic domain (24–210) |
| DN629_c0_g1_i1.p1 | 513.04 | 48 | 117 | yes (1–43) | Non-cytoplasmic domain (44–117) |
| DN4200_c0_g1_i1.p1 | 394.49 | 101 | 122 | yes (1–22) | Non-cytoplasmic domain (23–122), disorder prediction (20–108) |
| DN1209_c0_g2_i1.p1 | 143.95 | 62 | 309 | yes (1–16) | Non cytoplasmic domain (17–308), disorder prediction (26–135), proline rich |
| DN5_c1_g1_i1.p1 | 140.00 | 78 | 117 | no | Disorder prediction, coil (54–74) |
| DN355_c0_g1_i1.p1 | 36.43 | 30 | 107 | yes (1–34) | Non-cytoplasmic domain (15–107), disorder prediction (38–65) |
| DN21169_c0_g1_i1.p1 | 6.28 | 95 | 120 | yes (1–20) | Non-cytoplasmic domain (21–119) |
| DN5485_c1_g1_i2.p1 | 6.13 | 150 | 121 | no | Galactose-like binding sf, unknown (11–120) |
| DN646_c1_g1_i1.p1 | 3.93 | 25 | 330 | yes (1–20) | Non-cytoplasmic domain (21–330), disorder prediction (121–143), EGGSHELL |
| DN9918_c0_g1_i1.p1 | 3.40 | 239 | 204 | yes (1–18) | Non-cytoplasmic domain (19203–), Unknown (25–177) |
| DN68091_c0_g1_i1.p1 | 3.28 | 137 | 121 | no | Unknown unintegrated (41–113) |
| DN11320_c0_g1_i1.p1 | 2.83 | 280 | 113 | yes (1–17) | Non-cytoplasmic domain (18–113) |
| DN70554_c0_g1_i1.p1 | 2.80 | 305 | 112 | no | Disorder prediction (1–25) |
| DN39869_c0_g1_i1.p1 | 2.30 | 146 | 157 | yes (1–17) | Non-cytoplasmic domain (18–157) |
| DN8497_c0_g1_i1.p2 | 2.16 | 60 | 131 | yes (1–23) | Non-cytoplasmic domain (24–131) |
| DN16202_c0_g2_i1.p1 | 2.07 | 223 | 279 | no | NA, proline and cysteine rich, disorder prediction (101–139) |
| DN40599_c0_g1_i1.p1 | 1.79 | 65 | 152 | NA | NA |
| DN7825_c0_g1_i1.p1 | 1.66 | 176 | 223 | no | Non-cytoplasmic domain (37–222) |
| DN16202_c0_g1_i1.p1 | 1.52 | 461 | 282 | no | NA, proline and cysteine rich |
| DN40858_c0_g1_i1.p1 | 1.36 | 47 | 167 | NA | NA |
| DN78568_c0_g1_i1.p1 | 1.28 | 99 | 146 | no | Non-cytoplasmic domain (22–97) |
| DN60477_c0_g1_i1.p1 | 1.11 | 44 | 169 | no | Unknown integrated (1-121), cysteine rich |
| DN7590_c0_g1_i1.p1 | 0.99 | 278 | 168 | no | Unknown unintegrated (1–121) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
von Reumont, B.M.; Lüddecke, T.; Timm, T.; Lochnit, G.; Vilcinskas, A.; von Döhren, J.; Nilsson, M.A. Proteo-Transcriptomic Analysis Identifies Potential Novel Toxins Secreted by the Predatory, Prey-Piercing Ribbon Worm Amphiporus lactifloreus. Mar. Drugs 2020, 18, 407. https://doi.org/10.3390/md18080407
von Reumont BM, Lüddecke T, Timm T, Lochnit G, Vilcinskas A, von Döhren J, Nilsson MA. Proteo-Transcriptomic Analysis Identifies Potential Novel Toxins Secreted by the Predatory, Prey-Piercing Ribbon Worm Amphiporus lactifloreus. Marine Drugs. 2020; 18(8):407. https://doi.org/10.3390/md18080407
Chicago/Turabian Stylevon Reumont, Björn Marcus, Tim Lüddecke, Thomas Timm, Günter Lochnit, Andreas Vilcinskas, Jörn von Döhren, and Maria A. Nilsson. 2020. "Proteo-Transcriptomic Analysis Identifies Potential Novel Toxins Secreted by the Predatory, Prey-Piercing Ribbon Worm Amphiporus lactifloreus" Marine Drugs 18, no. 8: 407. https://doi.org/10.3390/md18080407
APA Stylevon Reumont, B. M., Lüddecke, T., Timm, T., Lochnit, G., Vilcinskas, A., von Döhren, J., & Nilsson, M. A. (2020). Proteo-Transcriptomic Analysis Identifies Potential Novel Toxins Secreted by the Predatory, Prey-Piercing Ribbon Worm Amphiporus lactifloreus. Marine Drugs, 18(8), 407. https://doi.org/10.3390/md18080407