α-Conotoxin as Potential to α7-nAChR Recombinant Expressed in Escherichia coli

Abstract
:1. Introduction
2. Results
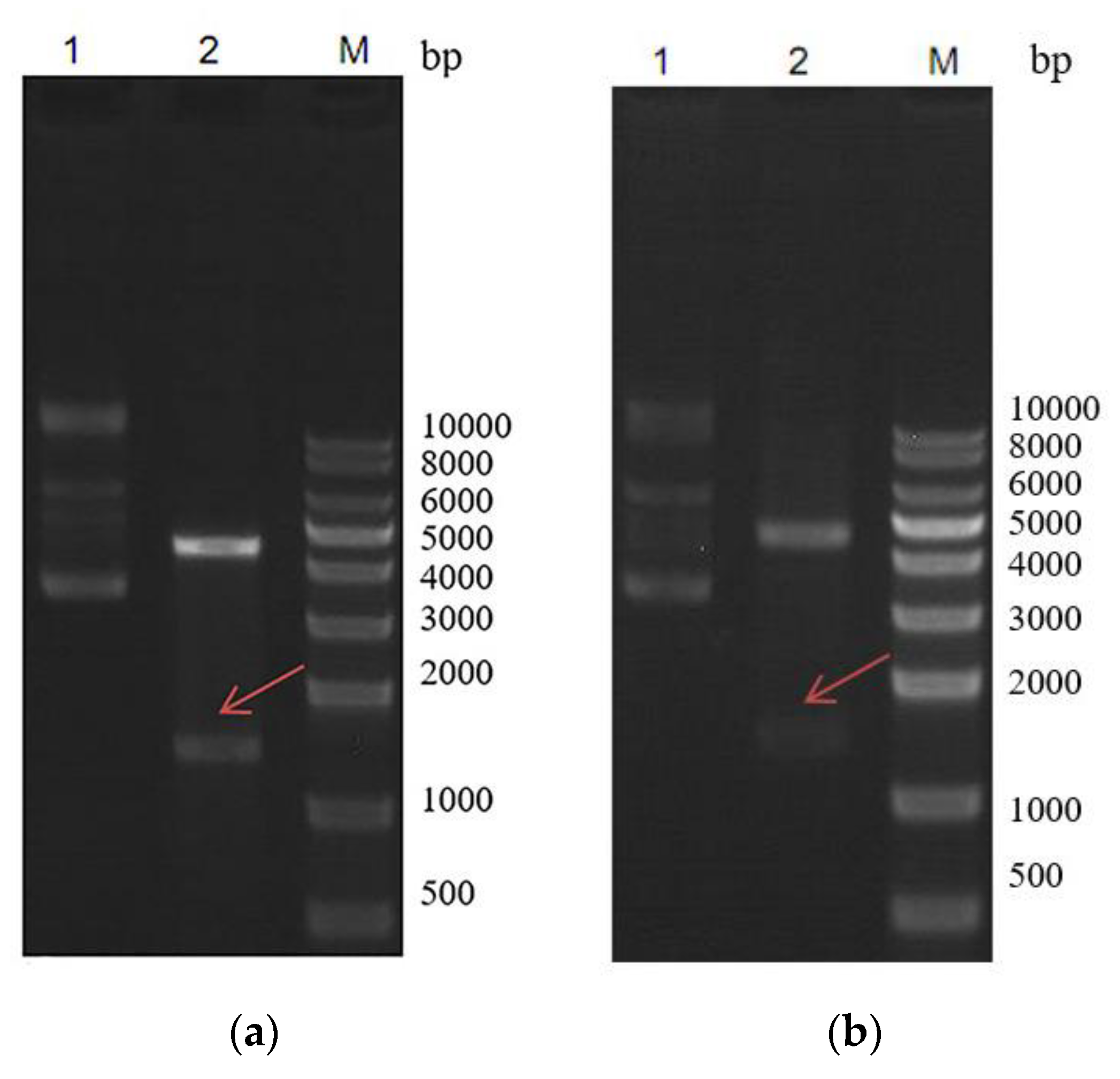
2.1. Construction of pET–ArIB and pET–ArIB (V11L, V16A) Expression Vector
2.2. Expression and Purification of Recombinant Trx-His6-ArIB and Trx-His6-ArIB (V11L, V16A) in Escherichia coli
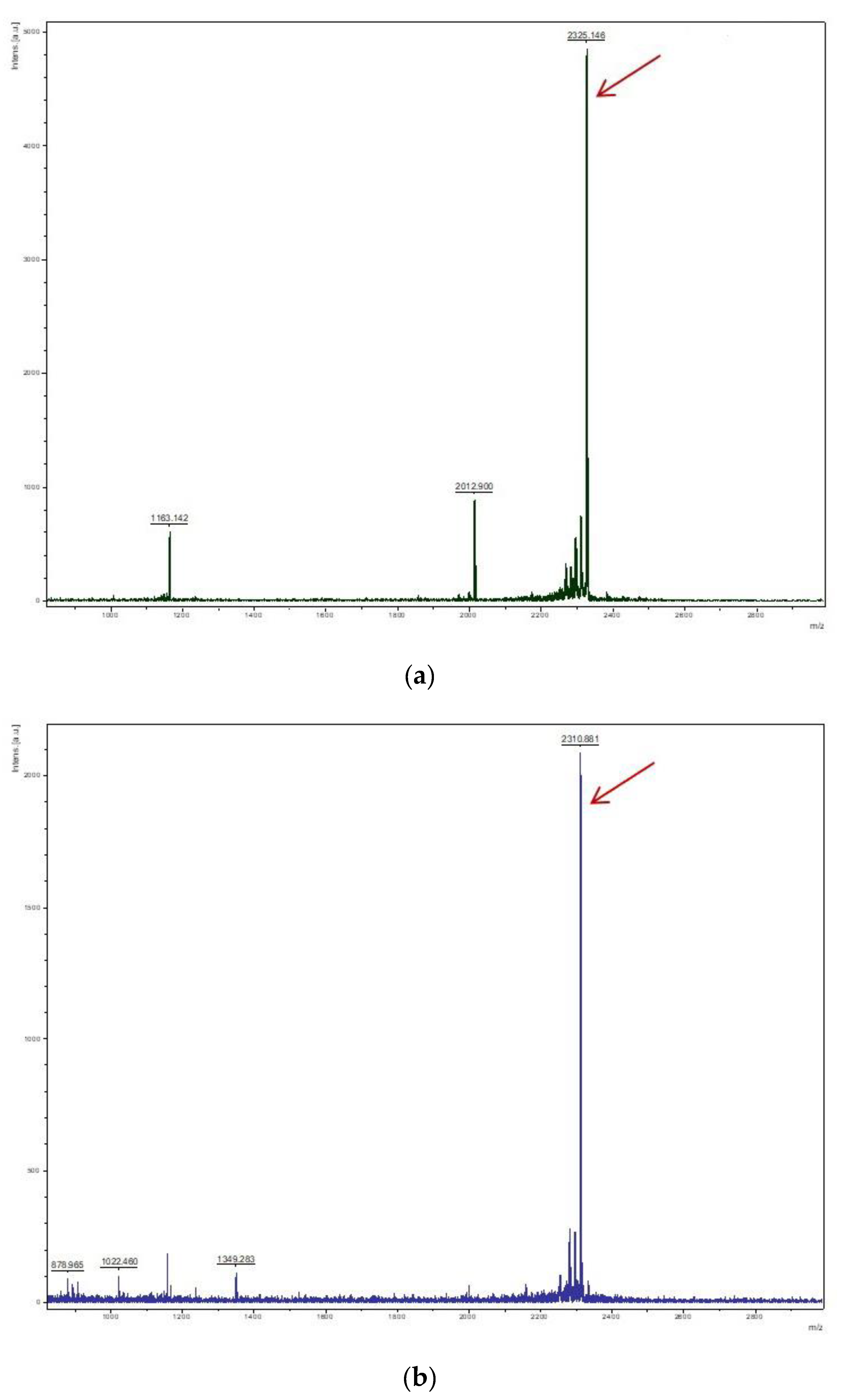
2.3. Cleavage and Purification of Trx-His-ArIB and Trx-His-ArIB (V11L, V16A)
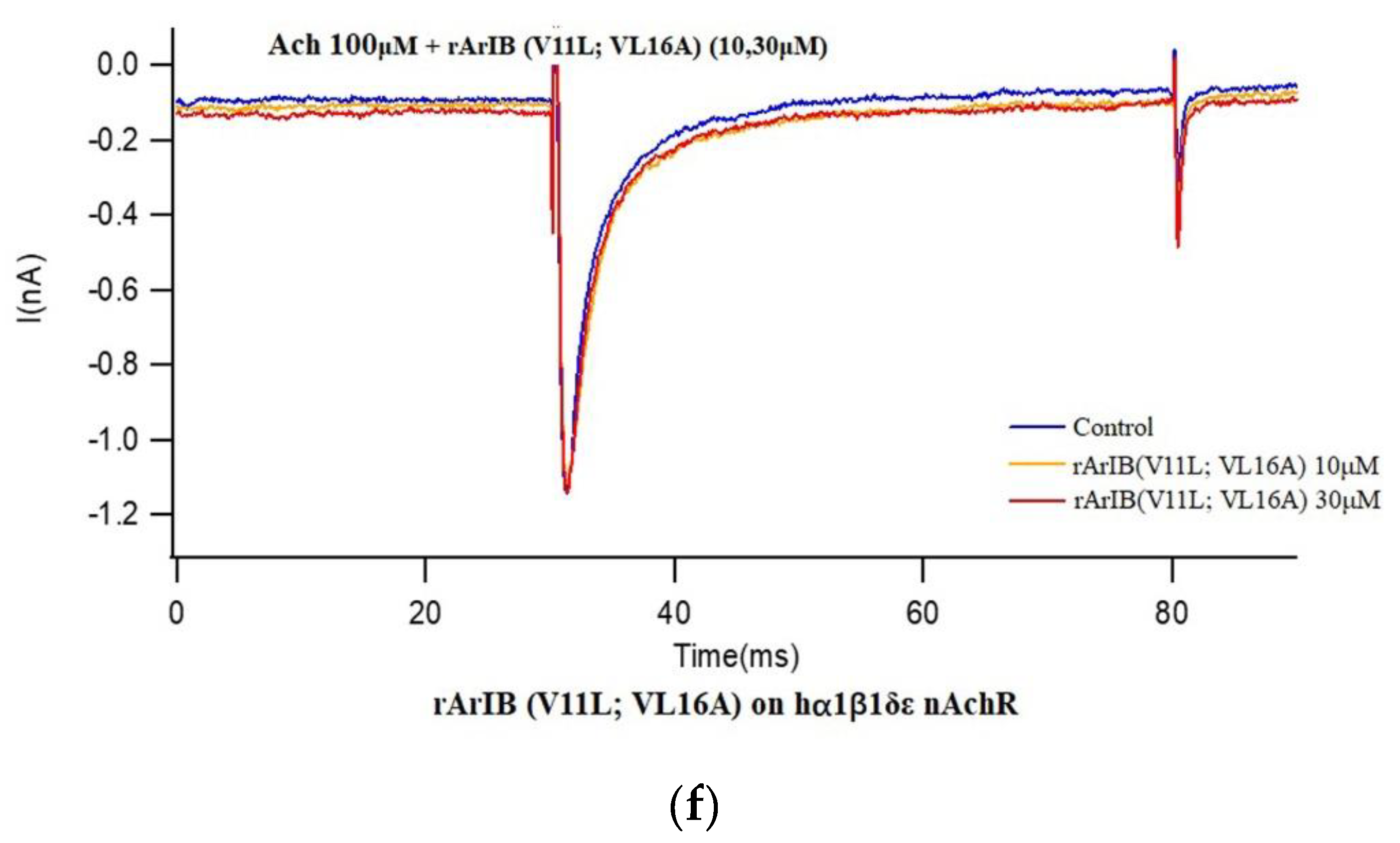
2.4. Effect of rArIB and rArIB (V11L, V16A) on ACh-Evoked Current of nAChRs
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmid Vectors and Reagents
4.2. Construction of pET–ArIB and pET–ArIB (V11L, V16A) Expression Vector
4.3. Recombinant Expression, Purification and Identification of Fusion Protein Trx-His6-ArIB and Trx-His6-ArIB (V11L, V16A)
4.4. Cleavage and Purification of Trx-His-ArIB and Trx-His-ArIB (V11L, V16A)
4.5. Mass Spectrometry
4.6. cRNA Preparation and Injection
4.7. Voltage Clamp Recording
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Changeux, J.P. The nicotinic acetylcholine receptor: The founding father of the pentameric ligand-gated ion channel superfamily. J. Biol. Chem. 2012, 287, 40207–40215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoli, M.; Pucci, S.; Vilella, A.; Gotti, C. Neuronal and extraneuronal nicotinic acetylcholine receptors. Curr. Neuropharmacol. 2018, 16, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Gotti, C.; Clementi, F.; Fornari, A.; Gaimarri, A.; Guiducci, S.; Manfredi, I.; Moretti, M.; Pedrazzi, P.; Pucci, L.; Zoli, M. Structural and functional diversity of native brain neuronal nicotinic receptors. Biochem. Pharmacol. 2009, 78, 703–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, T.L.; Bertrand, D. Importance of the nicotinic acetylcholine receptor system in the prefrontal cortex. Biochem. Pharmacol. 2013, 85, 1713–1720. [Google Scholar] [CrossRef]
- Hurst, R.; Rollema, H.; Bertrand, D. Nicotinic acetylcholine receptors: From basic science to therapeutics. Pharmacol. Ther. 2013, 137, 22–54. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [Green Version]
- Taly, A.; Corringer, P.J.; Guedin, D.; Lestage, P.; Changeux, J.-P. Nicotinic receptors: Allosteric transitions and therapeutic targets in the nervous system. Nat. Rev. Drug Discov. 2009, 8, 733–750. [Google Scholar] [CrossRef]
- Gotti, C.; Clementi, F. Neuronal nicotinic receptors: From structure to pathology. Prog. Neurobiol. 2004, 74, 363–396. [Google Scholar] [CrossRef]
- Ramachandran Nair, L.; Liu, X. Targeting the alpha4beta2- and alpha7-Subtypes of nicotinic acetylcholine receptors for smoking cessation medication development. J. Addict. Res. Ther. 2019, 10, 381. [Google Scholar]
- Dineley, K.T.; Pandya, A.A.; Yakel, J.L. Nicotinic ACh receptors as therapeutic targets in CNS disorders. Trends Pharmacol. Sci. 2015, 36, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Kalkman, H.O.; Feuerbach, D. Modulatory effects of α7 nAChRs on the immune system and its relevance for CNS disorders. Cell. Mol. Life Sci. 2016, 73, 2511–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, H.K.; Christensen, S.B.; Di Cesare Mannelli, L.; Gajewiak, J.; Ramachandra, R.; Elmslie, K.S.; Vetter, D.E.; Ghelardini, C.; Iadonato, S.P.; Mercado, J.L.; et al. Inhibition of α9α10 nicotinic acetylcholine receptors prevents chemotherapy-induced neuropathic pain. Proc. Natl. Acad. Sci. USA 2017, 114, E1825–E1832. [Google Scholar] [CrossRef] [Green Version]
- Verbitsky, M.; Rothlin, C.V.; Katz, E.; Belén Elgoyhen, A. Mixed nicotinic–muscarinic properties of the α9 nicotinic cholinergic receptor. Neuropharmacology 2000, 39, 2515–2524. [Google Scholar] [CrossRef]
- Sgard, F.; Charpantier, E.; Bertrand, S.; Walker, N.; Caput, D.; Graham, D.; Bertrand, D.; Besnard, F. A novel human nicotinic receptor subunit, α10, that confers functionality to the α9-Subunit. Mol. Pharmacol. 2002, 61, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, N.; Lewis, R.J. Neuronal nicotinic acetylcholine receptor modulators from cone snails. Mar. Drugs 2018, 16, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellison, M.; Haberlandt, C.; Gomez-Casati, M.E.; Watkins, M.; Elgoyhen, A.B.; McIntosh, J.M.; Olivera, B.M. α-RgIA: A novel conotoxin that specifically and potently blocks the α9α10 nAChR. Biochemistry 2006, 45, 1511–1517. [Google Scholar]
- Di Cesare Mannelli, L.; Cinci, L.; Micheli, L.; Zanardelli, M.; Pacini, A.; McIntosh, J.M.; Ghelardini, C. α-conotoxin RgIA protects against the development of nerve injury-induced chronic pain and prevents both neuronal and glial derangement. Pain 2014, 155, 1986–1995. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Kompella, S.N.; Adams, D.J.; Craik, D.J.; Kaas, Q. Determination of the α-Conotoxin Vc1.1 binding site on the α9α10 nicotinic acetylcholine receptor. J. Med. Chem. 2013, 56, 3557–3567. [Google Scholar]
- Indurthi, D.C.; Pera, E.; Kim, H.-L.; Chu, C.; McLeod, M.D.; McIntosh, J.M.; Absalom, N.L.; Chebib, M. Presence of multiple binding sites on α9α10 nAChR receptors alludes to stoichiometric-dependent action of the α-conotoxin, Vc1.1. Biochem. Pharmacol. 2014, 89, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.J.; Berecki, G. Mechanisms of conotoxin inhibition of N-type (Cav2.2) calcium channels. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Huynh, T.G.; Cuny, H.; Slesinger, P.A.; Adams, D.J. Novel mechanism of Voltage-Gated N-type (Cav2.2) calcium channel inhibition revealed through α-Conotoxin Vc1.1 Activation of the GABAB Receptor. Mol. Pharmacol. 2015, 87, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.J.; Callaghan, B.; Berecki, G. Analgesic conotoxins: Block and G protein-coupled receptor modulation of N-type (Ca(V) 2.2) calcium channels. Br. J. Pharmacol. 2012, 166, 486–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, A.; Kompella, S.N.; Akondi, K.B.; Melaun, C.; Daly, N.L.; Luetje, C.W.; Alewood, P.F.; Craik, D.J.; Adams, D.J.; Marí, F. RegIIA: An α4/7-conotoxin from the venom of Conus regius that potently blocks α3β4 nAChRs. Biochem. Pharmacol. 2012, 83, 419–426. [Google Scholar] [CrossRef]
- Ning, J.; Ren, J.; Xiong, Y.; Wu, Y.; Zhangsun, M.; Zhangsun, D.; Zhu, X.; Luo, S. Identification of crucial residues in α-Conotoxin EI inhibiting muscle nicotinic acetylcholine receptor. Toxins 2019, 11, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Zhangsun, D.; Zhu, X.; Wu, Y.; Hu, Y.; Christensen, S.; Harvey, P.J.; Akcan, M.; Craik, D.J.; McIntosh, J.M. Characterization of a Novel α-Conotoxin TxID from Conus textile that potently blocks rat α3β4 Nicotinic acetylcholine receptors. J. Med. Chem. 2013, 56, 9655–9663. [Google Scholar] [PubMed] [Green Version]
- Yu, J.; Zhu, X.; Harvey, P.J.; Kaas, Q.; Zhangsun, D.; Craik, D.J.; Luo, S. Single amino acid substitution in α-Conotoxin TxID Reveals a Specific α3β4 Nicotinic acetylcholine receptor antagonist. J. Med. Chem. 2018, 61, 9256–9265. [Google Scholar]
- Whiteaker, P.; Christensen, S.; Yoshikami, D.; Dowell, C.; Watkins, M.; Gulyas, J.; Rivier, J.; Olivera, B.M.; McIntosh, J.M. Discovery, synthesis, and structure activity of a highly selective α7 nicotinic Acetylcholine receptor antagonist. Biochemistry 2007, 46, 6628–6638. [Google Scholar]
- Hone, A.J.; Whiteaker, P.; Mohn, J.L.; Jacob, M.H.; McIntosh, J.M. Alexa Fluor 546-ArIB[V11L;V16A] is a potent ligand for selectively labeling alpha 7 nicotinic acetylcholine receptors. J. Neurochem. 2010, 114, 994–1006. [Google Scholar]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef]
- Rafia, M.; Sajjad, K.; Mohammad Amjad, K.; Cornelia, M.W.; Zeenat, M. Conotoxins: Structure, therapeutic potential and pharmacological applications. Curr. Pharm. Des. 2016, 22, 582–589. [Google Scholar]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, synthesis, and structure–Activity relationships of conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef]
- El Hamdaoui, Y.; Wu, X.; Clark, R.J.; Giribaldi, J.; Anangi, R.; Craik, D.J.; King, G.F.; Dutertre, S.; Kaas, Q.; Herzig, V.; et al. Periplasmic expression of 4/7 α-conotoxin TxIA analogs in E. coli favors ribbon isomer formation—Suggestion of a binding mode at the α7 nAChR. Front. Pharmacol. 2019, 10, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Zhu, X.; Yang, Y.; Luo, S.; Zhangsun, D. Expression in Escherichia coli of fusion protein comprising α-conotoxin TxIB and preservation of selectivity to nicotinic acetylcholine receptors in the purified product. Chem. Biol. Drug Des. 2018, 91, 349–358. [Google Scholar] [CrossRef]
- Zhu, X.; Bi, J.; Yu, J.; Li, X.; Zhang, Y.; Zhangsun, D.; Luo, S. Recombinant expression and characterization of α-Conotoxin LvIA in Escherichia coli. Mar. Drugs 2016, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Wang, L.; Wu, Y.; Liu, J.; Sun, D.; Zhu, X.; Feng, Y.; Qin, M.; Chen, S.; Xu, A. Soluble expression and sodium channel activity of lt16a, a novel framework XVI conotoxin from the M-superfamily. Toxicon 2015, 98, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, L.; Zhou, M.; Jiang, X.; Zhu, X.; Chen, Y.; Luo, S.; You, Y.; Ren, Z.; Xu, A. Soluble expression, purification and functional identification of the framework XV conotoxins derived from different Conus species. Peptides 2014, 56, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhangsun, D.; Hu, Y.; Wu, Y.; Sheng, L.; Fang, L.; Wu, X.; Yu, J.; Luo, S. Expression and secretion of functional recombinant μO-conotoxin MrVIB-His-tag in Escherichia coli. Toxicon 2013, 72, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Zhangsun, D.; Wu, Y.; Lin, B.; Zhu, X.; Luo, S. Expression, renaturation and biological activity of recombinant conotoxin GeXIVAWT. Appl. Microbiol. Biotechnol. 2013, 97, 1223–1230. [Google Scholar] [CrossRef]
- Spiezia, M.C.; Chiarabelli, C.; Polticelli, F. Recombinant expression and insecticidal properties of a Conus ventricosus conotoxin-GST fusion protein. Toxicon 2012, 60, 744–751. [Google Scholar] [CrossRef]
- Pi, C.; Liu, J.; Wang, L.; Jiang, X.; Liu, Y.; Peng, C.; Chen, S.; Xu, A. Soluble expression, purification and functional identification of a disulfide-rich conotoxin derived from Conus litteratus. J. Biotechnol. 2007, 128, 184–193. [Google Scholar] [CrossRef]
- Zhan, J.; Chen, X.; Wang, C.; Qiu, J.; Ma, F.; Wang, K.; Zheng, S. A fusion protein of conotoxin MVIIA and thioredoxin expressed in Escherichia coli has significant analgesic activity. Biochem. Bioph. Res. Co. 2003, 311, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Cuebas, L.M.; White, M.M. Expression of a biologically-active conotoxin PrIIIE in Escherichia coli. Protein Expr. Purif. 2012, 82, 6–10. [Google Scholar] [CrossRef]
- Bruce, C.; Fitches Ec Fau-Chougule, N.; Chougule N Fau-Bell, H.A.; Bell Ha Fau-Gatehouse, J.A.; Gatehouse, J.A. Recombinant conotoxin, TxVIA, produced in yeast has insecticidal activity. Toxicon 2011, 58, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Klint, J.K.; Senff S Fau-Saez, N.J.; Saez Nj Fau-Seshadri, R.; Seshadri R Fau-Lau, H.Y.; Lau Hy Fau-Bende, N.S.; Bende Ns Fau-Undheim, E.A.B.; Undheim Ea Fau-Rash, L.D.; Rash Ld Fau-Mobli, M.; Mobli M Fau-King, G.F.; King, G.F. Production of recombinant disulfide-rich venom peptides for structural and functional analysis via expression in the periplasm of E. coli. PLoS ONE 2013, 8, e63865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, J.M.; Gardner, S.; Luo, S.; Garrett, J.E.; Yoshikami, D. Conus peptides: Novel probes for nicotinic acetylcholine receptor structure and function. Eur. J. Pharmacol. 2000, 393, 205–208. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| ArIB Forward primer 1 | 5′-CGACGACGACGACAAGGATGAATGCTGTAGCAACCCGGCGTGCCGCGTGAACAATCCGCATGTTTGTCGTCGCCGTTAAG -3′ |
| Reverse primer 1 | 5′-AATTCTTAACGGCGACGACAAACATGCGGATTGTTCACGCGGCACGCCGGGTTGCTACAGCATTCATCCTTGTCGTCGTCGTCGGTAC -3′ |
| ArIB (V11L, V16A) Forward primer 2 | 5′-CGACGACGACGACAAGGATGAATGCTGTAGCAACCCGGCGTGCCGCCTGAACAATCCGCATGCATGTCGTCGCCGTTAAG-3′ |
| Reverse primer 2 | 5′-AATTCTTAACGGCGACGACATGCATGCGGATTGTTCAGGCGGCACGCCGGGTTGCTACAGCATTCATCCTTGTCGTCGTCGTCGGTAC -3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yin, Y.; Song, Y.; Wang, K.; Wu, F.; Jiang, H. α-Conotoxin as Potential to α7-nAChR Recombinant Expressed in Escherichia coli. Mar. Drugs 2020, 18, 422. https://doi.org/10.3390/md18080422
Liu Y, Yin Y, Song Y, Wang K, Wu F, Jiang H. α-Conotoxin as Potential to α7-nAChR Recombinant Expressed in Escherichia coli. Marine Drugs. 2020; 18(8):422. https://doi.org/10.3390/md18080422
Chicago/Turabian StyleLiu, Yanli, Yifeng Yin, Yunyang Song, Kang Wang, Fanghui Wu, and Hui Jiang. 2020. "α-Conotoxin as Potential to α7-nAChR Recombinant Expressed in Escherichia coli" Marine Drugs 18, no. 8: 422. https://doi.org/10.3390/md18080422
APA StyleLiu, Y., Yin, Y., Song, Y., Wang, K., Wu, F., & Jiang, H. (2020). α-Conotoxin as Potential to α7-nAChR Recombinant Expressed in Escherichia coli. Marine Drugs, 18(8), 422. https://doi.org/10.3390/md18080422