In Vitro Evaluation of Antioxidant Potential of the Invasive Seagrass Halophila stipulacea


 ,
,  ,
,  , , ,
, , , 

Abstract
:1. Introduction
2. Results
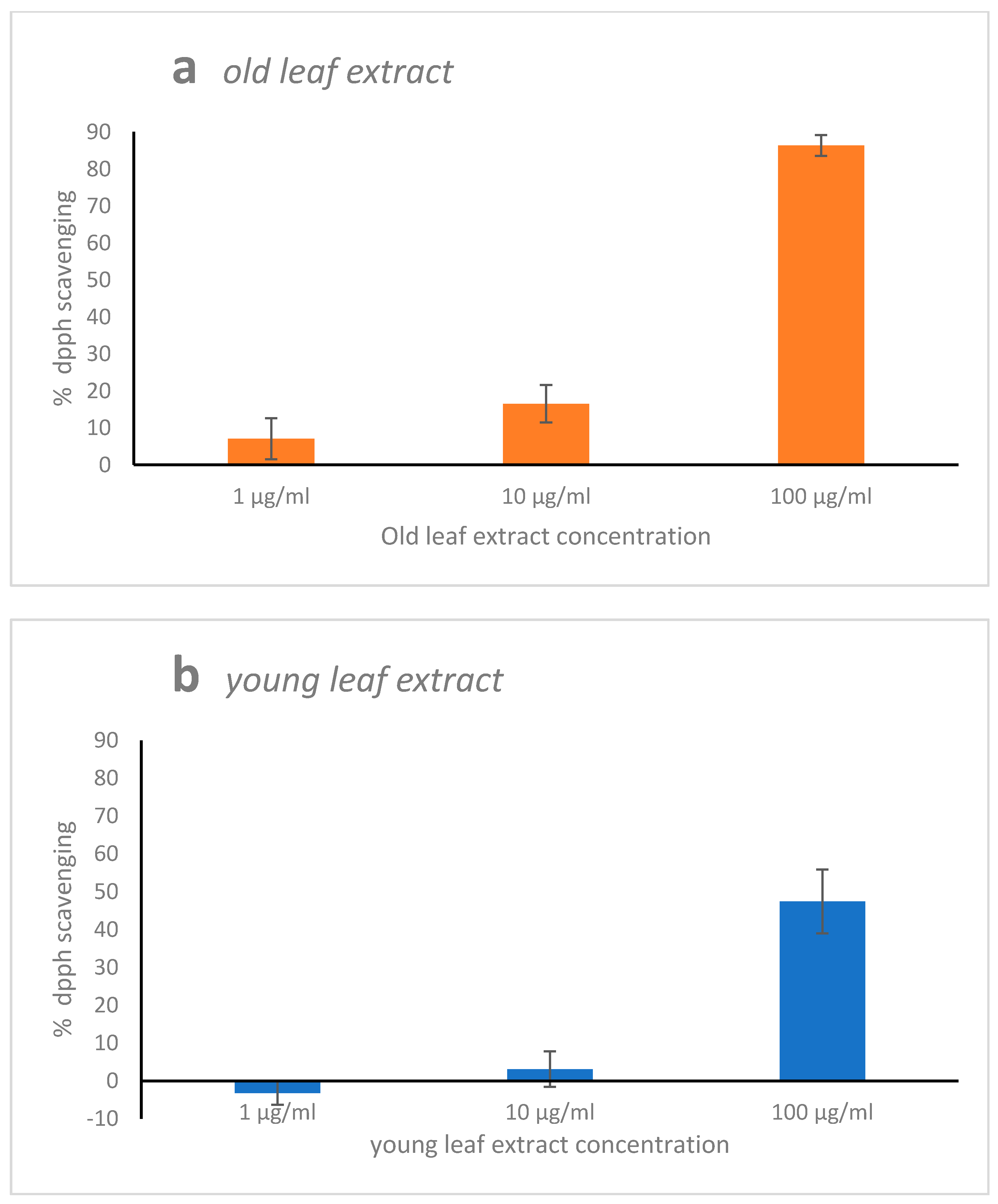
2.1. 2,2,1-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity
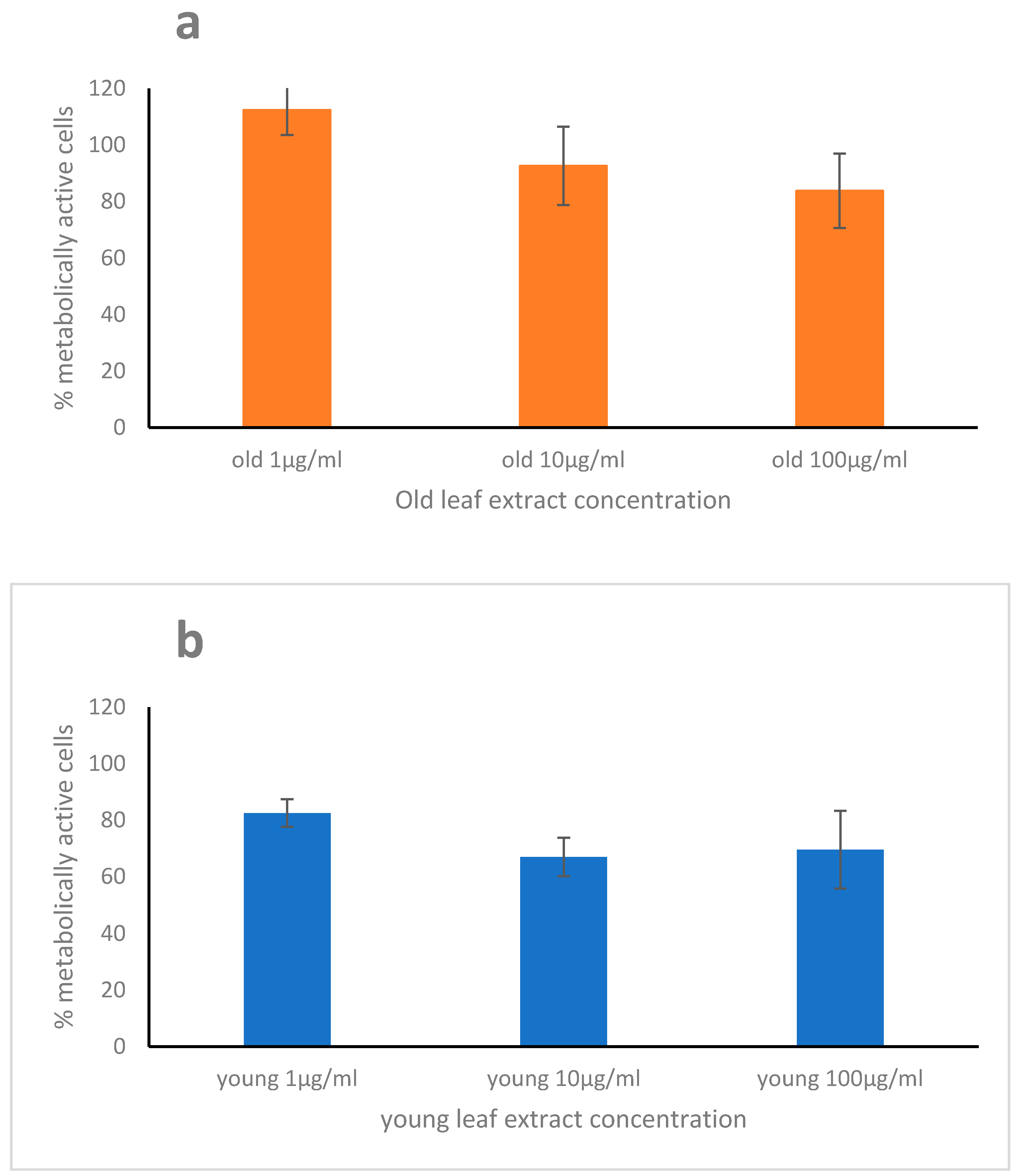
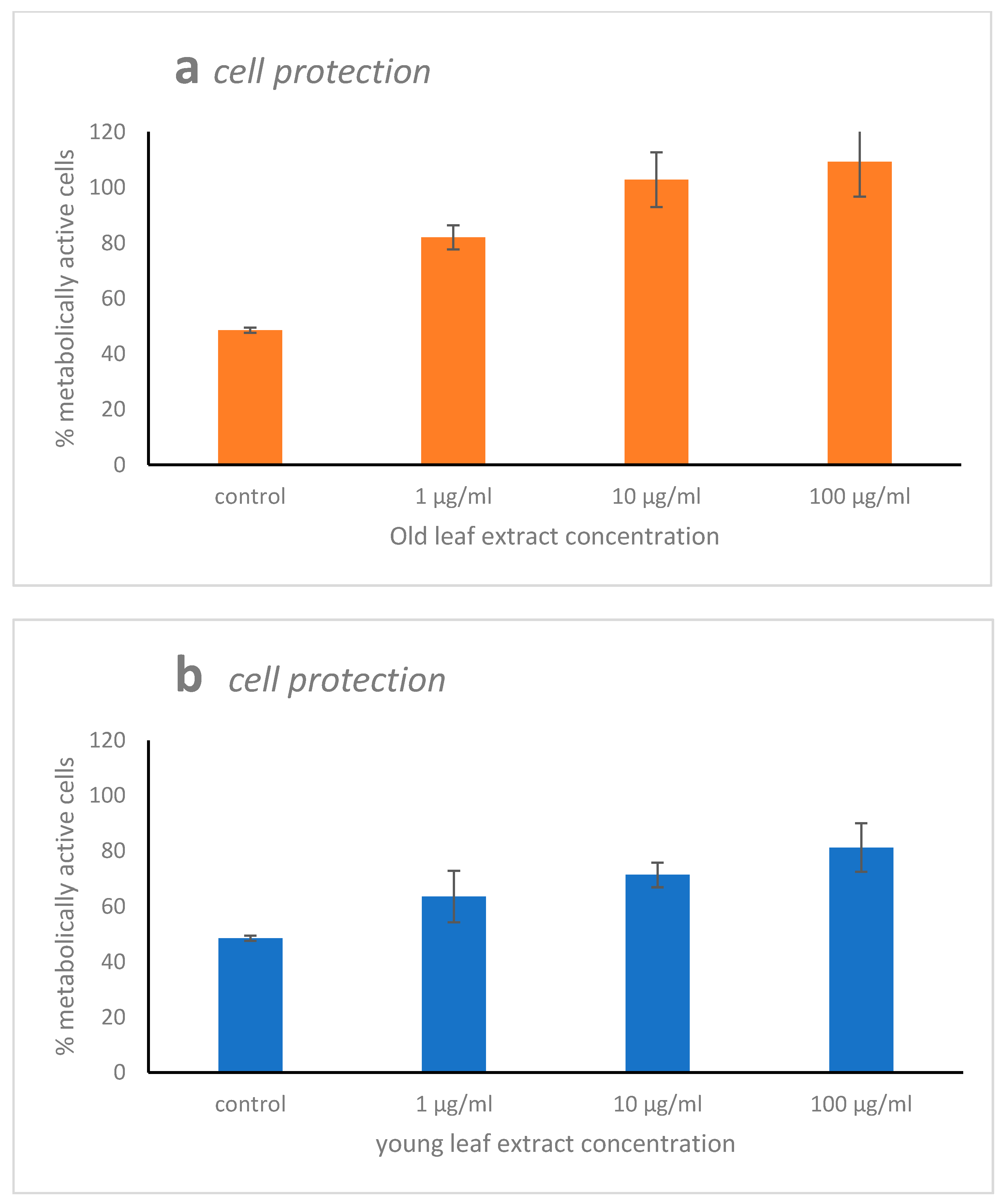
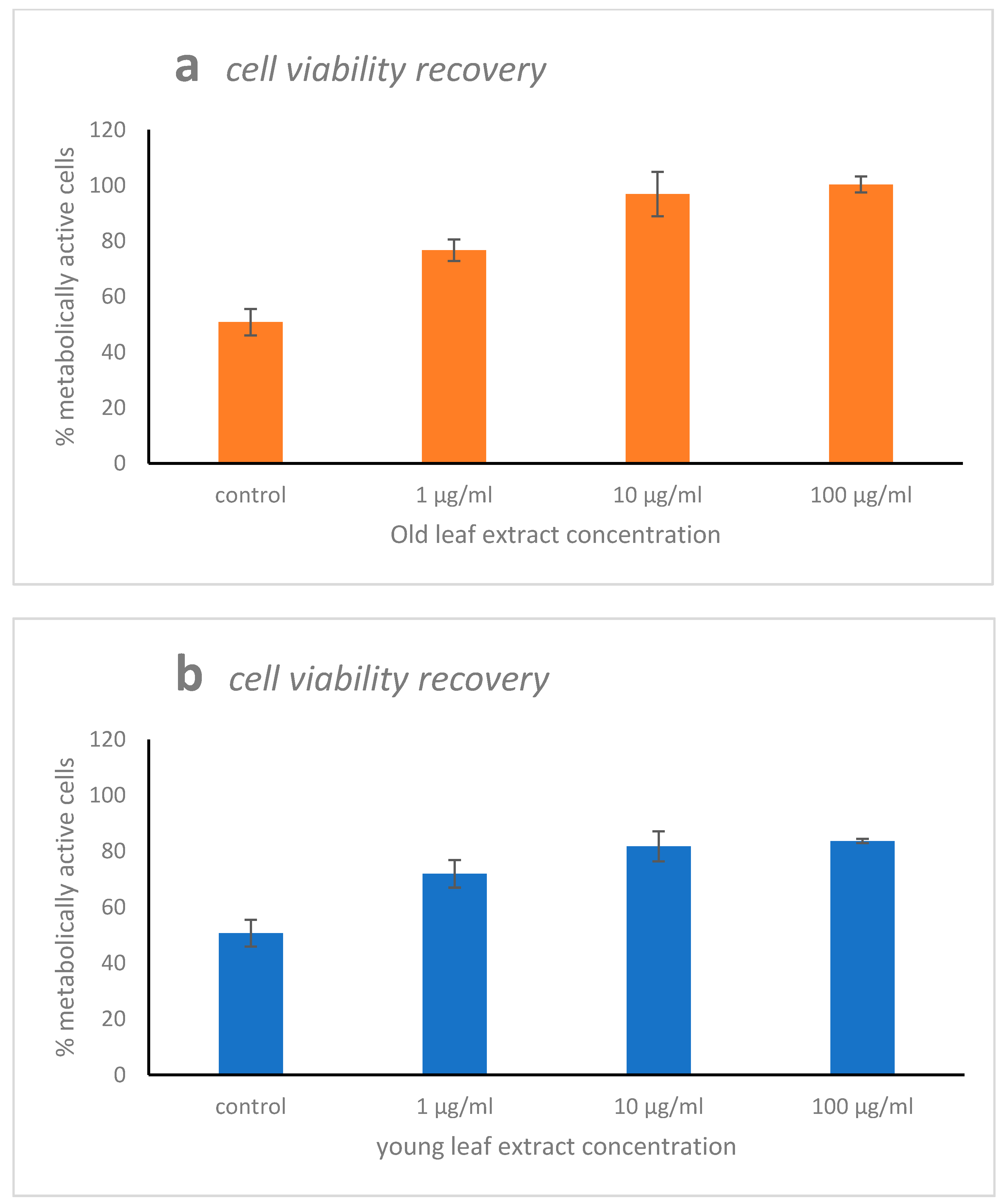
2.2. Cytotoxicity, Scavenging, and Repair Activities of the Leaf Extracts
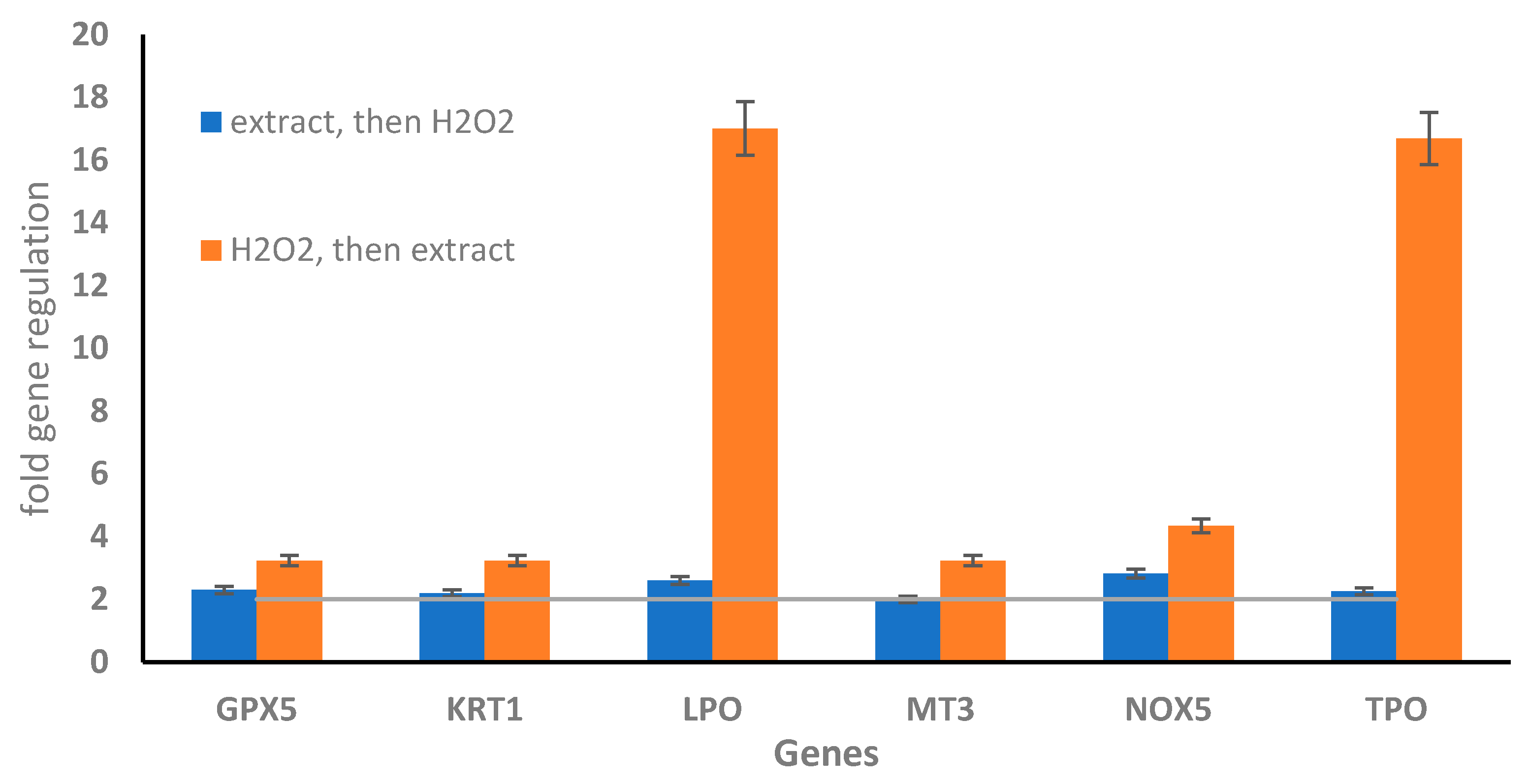
2.3. Oxidative Stress Gene Expression in WI-38 Cells Treated with Old Leaf Extracts
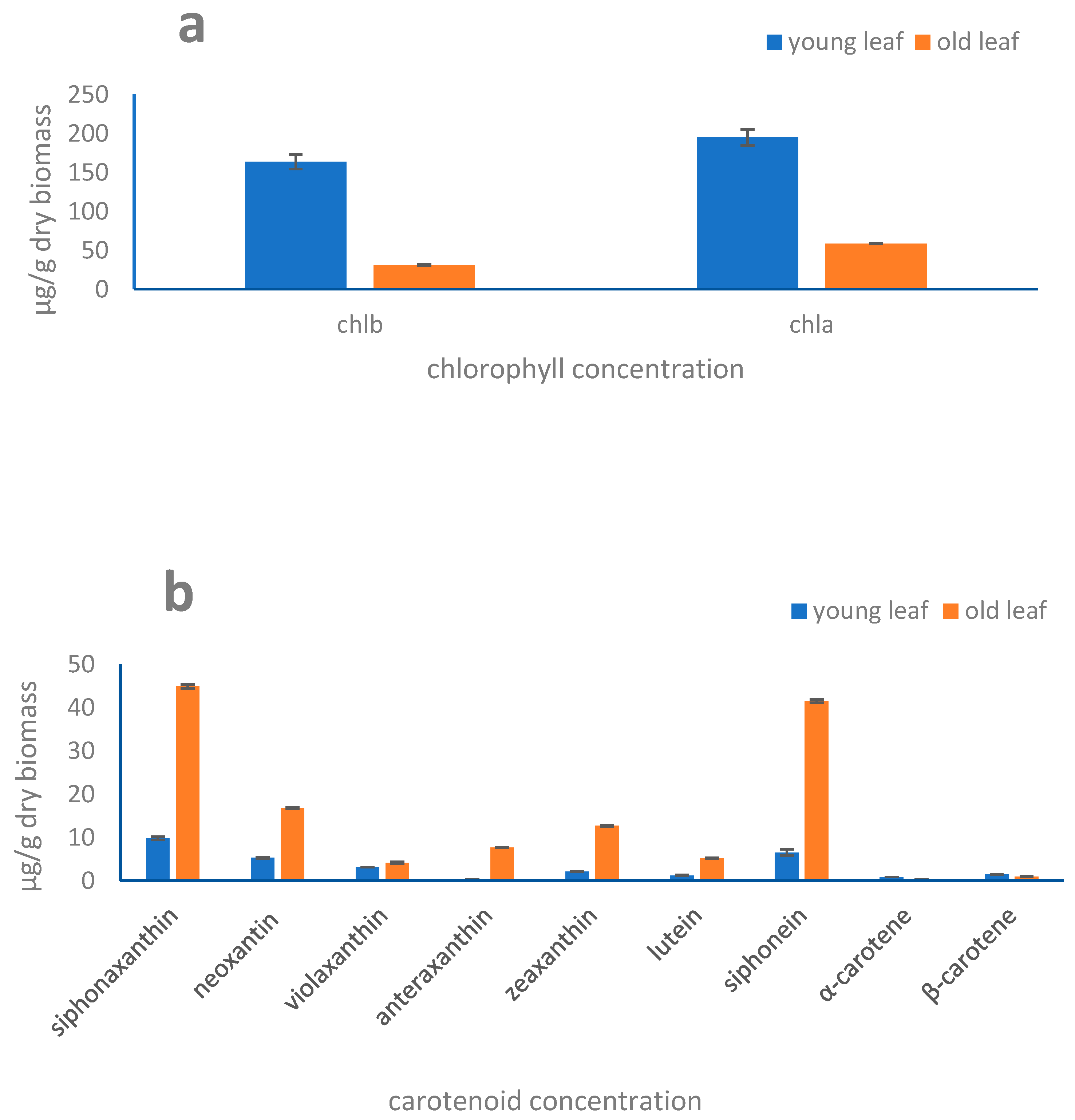
2.4. Pigments
2.5. Macromolecular Composition
3. Discussion
4. Materials and Methods
4.1. Halophila Stipulacea Sampling and Leaf Collection
4.2. Preparation of Ethanol/Water Extracts from Halophila Stipulacea Leaves
4.3. Pigment HPLC Analysis
4.4. Macromolecular Composition of the Extracts
4.5. DPPH Free Radical Scavenging Activity
4.6. Human WI-38 Cell Line Culture and Treatments
4.7. Cell Viability Assays
4.8. RNA Extraction and Real-Time qPCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pandel, R.; Poljšak, B.; Godic, A.; Dahmane, R. Skin Photoaging and the Role of Antioxidants in Its Prevention. ISRN Dermatol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McChesney, J.D.; Venkataraman, S.K.; Henri, J.T. Plant natural products: Back to the future or into extinction? Phytochemistry 2007, 68, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 9, 3012–3043. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Nuzzo, G.; Galasso, C.; Casotti, R.; Fontana, A.; Romano, G.; Ianora, A. The Marine Dinoflagellate Alexandrium andersoni Induces Cell Death in Lung and Colorectal Tumor Cell Lines. Mar. Biotechnol. 2018, 20, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Galasso, C.; Nuzzo, G.; Brunet, C.; Ianora, A.; Sardo, A.; Fontana, A.; Sansone, C. The marine dinoflagellate alexandrium minutum activates a mitophagic pathway in human lung cancer cells. Mar. Drugs 2018, 16, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zidorn, C. Secondary metabolites of seagrasses (Alismatales and Potamogetonales; Alismatidae): Chemical diversity, bioactivity, and ecological function. Phytochemistry 2016, 124, 5–28. [Google Scholar] [CrossRef]
- Rocha, D.H.A.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed secondary metabolites in vitro and in vivo anticancer activity. Mar. Drugs 2018, 16, 410. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.; Lundqvist, L.C.E.; Jam, M.; Jeudy, A.; Barbeyron, T.; Sandström, C.; Michel, G.; Czjzek, M. Comparative characterization of two marine alginate lyases from zobellia galactanivorans reveals distinct modes of action and exquisite adaptation to their natural substrate. J. Biol. Chem. 2013, 288, 23021–23037. [Google Scholar] [CrossRef] [Green Version]
- Yuvaraj, N.; Kanmani, P.; Satishkumar, R.; Paari, A.; Pattukumar, V.; Arul, V. Seagrass as a potential source of natural antioxidant and anti-inflammatory agents. Pharm. Biol. 2012, 50, 458–467. [Google Scholar] [CrossRef] [Green Version]
- Saranya, K.; Balamani, K.; Kokila, P.; Maruthupandian, A.; Viji, M. Biochemical and bioactive potential of seagrass Halophila ovalis (R. Brown) J.D. Hooker and Thalassia hemprichii (Ehrenb. Ex solms) Asch. Int. J. Adv. Interdiscip. Res. 2017, 4, 51–55. [Google Scholar] [CrossRef]
- Manh Nguyen, H.; Savva, I.; Kleitou, P.; Kletou, D.; Lima, F.P.; Sapir, Y.; Winters, G. Seasonal dynamics of native and invasive Halophila stipulacea populations—A case study from the northern Gulf of Aqaba and the eastern Mediterranean Sea. Aquat. Bot. 2020, 162, 103205. [Google Scholar] [CrossRef] [Green Version]
- Winters, G.; Beer, S.; Willette, D.A.; Viana, I.G.; Chiquillo, K.L.; Beca-Carretero, P.; Villamayor, B.; Azcárate-García, T.; Shem-Tov, R.; Mwabvu, B.; et al. The tropical seagrass Halophila stipulacea: Reviewing what we know from its native and invasive habitats, alongside identifying knowledge gaps. Front. Mar. Sci. 2020, 7, 1–28. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the world’s worst invasive alien species. A selection from the Global Invasive Species Database. ISSG Spec. Group SSC World Conserv. Union IUCN 2000. [Google Scholar] [CrossRef]
- Sghaier, Y.R.; Zakhama-Sraieb, R.; Benamer, I.; Charfi-Cheikhrouha, F. Occurrence of the seagrass Halophila stipulacea (Hydrocharitaceae) in the southern Mediterranean Sea. Bot. Mar. 2011, 54, 575–582. [Google Scholar] [CrossRef]
- Borghini, M.; Bryden, H.; Schroeder, K.; Sparnocchia, S.; Vetrano, A. The Mediterranean is becoming saltier. Ocean Sci. 2014, 10, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Mabrouk, S.B.; Reis, M.; Sousa, M.L.; Ribeiro, T.; Almeida, J.R.; Pereira, S.; Antunes, J.; Rosa, F.; Vasconcelos, V.; Achour, L.; et al. The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling. Mar. Drugs 2020, 18, 88. [Google Scholar] [CrossRef] [Green Version]
- Kannan Rengasamy, R.R.; Rajasekaran, A.; Micheline, G.D.; Perumal, A. Antioxidant activity of seagrasses of the Mandapam coast, India. Pharm. Biol. 2012, 50, 182–187. [Google Scholar] [CrossRef]
- Brunet, C.; Brylinski, J.M.; Lemoine, Y. In situ variations of the xanthophylls diatoxanthin and diadinoxanthin—Photoadaptation and relationships with a hydrodynamical system in the eastern English Channel. Mar. Ecol. Prog. Ser. 1993. [Google Scholar] [CrossRef]
- Raniello, R.; Lorenti, M.; Brunet, C.; Buia, M.C. Photoacclimation of the invasive alga Caulerpa racemosa var. cylindracea to depth and daylight patterns and a putative new role for siphonaxanthin. Mar. Ecol. 2006. [Google Scholar] [CrossRef]
- Toussaint, O.; Medrano, E.E.; Von Zglinicki, T. Cellular and molecular mechanisms of stress-induced premature senescence (SIPS) of human diploid fibroblasts and melanocytes. Exp. Gerontol. 2000, 35, 927–945. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.R.; Choi, M.J.; Choi, J.M.; Ko, J.C.; Ko, J.Y.; Cho, E.J. Malvidin Protects WI-38 Human Fibroblast Cells Against Stress-induced Premature Senescence. J. Cancer Prev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wolf, F.I.; Torsello, A.; Covacci, V.; Fasanella, S.; Montanari, M.; Boninsegna, A.; Cittadini, A. Oxidative DNA damage as a marker of aging in WI-38 human fibroblasts. Exp. Gerontol. 2002, 102, 69–77. [Google Scholar] [CrossRef]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J.C. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Van Tussenbroek, B.I.; van Katwijk, M.M.; Bouma, T.J.; van der Heide, T.; Govers, L.L.; Leuven, R.S.E.W. Non-native seagrass Halophila stipulacea forms dense mats under eutrophic conditions in the Caribbean. J. Sea Res. 2016, 115, 1–5. [Google Scholar] [CrossRef]
- Hyytiäinen, K.; Lehtiniemi, M.; Niemi, J.K.; Tikka, K. An optimization framework for addressing aquatic invasive species. Ecol. Econ. 2013, 91, 69–79. [Google Scholar] [CrossRef]
- Beer, S.; Waisel, Y. Effects of light and pressure on photosynthesis in two seagrasses. Aquat. Bot. 1982, 13, 331–337. [Google Scholar] [CrossRef]
- Schwarz, A.M.; Hellblom, F. The photosynthetic light response of Halophila stipulacea growing along a depth gradient in the Gulf of Aqaba, the Red Sea. Aquat. Bot. 2002, 74, 263–272. [Google Scholar] [CrossRef]
- Sharon, Y.; Silva, J.; Santos, R.; Runcie, J.W.; Chernihovsky, M.; Beer, S. Photosynthetic responses of Halophila stipulacea to a light gradient. II. Acclimations following transplantation. Aquat. Bot. 2009, 7, 153–157. [Google Scholar] [CrossRef]
- Mejia, A.Y.; Rotini, A.; Lacasella, F.; Bookman, R.; Thaller, M.C.; Shem-Tov, R.; Winters, G.; Migliore, L. Assessing the ecological status of seagrasses using morphology, biochemical descriptors and microbial community analyses. A study in Halophila stipulacea (Forsk.) Aschers meadows in the northern Red Sea. Ecol. Indic. 2016, 60, 1150–1163. [Google Scholar] [CrossRef]
- Santos, F.; Monteiro, J.P.; Duarte, D.; Melo, T.; Lopes, D.; da Costa, E.; Domingues, M.R. Unraveling the lipidome and antioxidant activity of native bifurcaria bifurcata and invasive sargassum muticum seaweeds: A lipid perspective on how systemic intrusion may present an opportunity. Antioxidants 2020, 9, 642. [Google Scholar] [CrossRef] [PubMed]
- Kannan, R.R.R.; Arumugam, R.; Anantharaman, P. In vitro antioxidant activities of ethanol extract from Enhalus acoroides (L.F.) Royle. Asian Pac. J. Trop. Med. 2010, 898–901. [Google Scholar] [CrossRef] [Green Version]
- Neelima, C.S.S.; Seenivasan, R. DPPH Radical Scavenging Activity of Selected Seagrasses from South East Coast of India. Int. J. Adv. Res. 2015, 3, 950–956. [Google Scholar]
- Jeyapragash, D.; Subhashini, P.; Raja, S.; Abirami, K.; Thangaradjou, T. Evaluation of In-vitro Antioxidant Activity of Seagrasses: Signals for Potential Alternate Source. Free Radic. Antioxid. 2016, 6, 77–89. [Google Scholar] [CrossRef]
- Pushpa Bharathi, N.; Amudha, P.; Vanitha, V. Sea grasses—Novel marine nutraceuticals. Int. J. Pharm. Bio. Sci. 2016, 7, 567–573. [Google Scholar]
- Gullón, B.; Eibes, G.; Moreira, M.T.; Herrera, R.; Labidi, J.; Gullón, P. Yerba mate waste: A sustainable resource of antioxidant compounds. Ind. Crops Prod. 2018, 113, 398–405. [Google Scholar] [CrossRef]
- Santhanam, R.; Santhanam, R.; Suleria, H.; Santhanam, R.; Santhanam, R.; Suleria, H. Biology and Ecology of Pharmaceutical Marine Plants. Biol. Ecol. Pharm. Mar. Plants 2018. [Google Scholar] [CrossRef]
- Woo, H.R.; Masclaux-Daubresse, C.; Lim, P.O. Plant senescence: How plants know when and how to die. J. Exp. Bot. 2018, 69, 715–718. [Google Scholar] [CrossRef]
- Drew, E.A. Factors affecting photosynthesis and its seasonal variation in the seagrasses Cymodocea nodosa (Ucria) Aschers, and Posidonia oceanica (L.) Delile in the mediterranean. J. Exp. Mar. Biol. Ecol. 1978, 31, 173–194. [Google Scholar] [CrossRef]
- Pourtau, N.; Jennings, R.; Pelzer, E.; Pallas, J.; Wingler, A. Effect of sugar-induced senescence on gene expression and implications for the regulation of senescence in Arabidopsis. Planta 2006, 224, 556–568. [Google Scholar] [CrossRef] [PubMed]
- Socaci, S.A.; Rugină, D.O.; Diaconeasa, Z.M.; Pop, O.L.; Fărcaș, A.C.; Păucean, A.; Tofană, M.; Pintea, A. Antioxidant Compounds Recovered from Food Wastes. Funct. Food Improv. Health Adequate Food 2017. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A.; Purdy, S.; MacLean, J.A.; Pourtau, N. The role of sugars in integrating environmental signals during the regulation of leaf senescence. Proc. J. Exp. Bot. 2006, 57, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A.; Masclaux-Daubresse, C.; Fischer, A.M. Sugars, senescence, and ageing in plants and heterotrophic organisms. Proc. J. Exp. Bot. 2009, 60, 1063–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Doorn, W.G. Is the onset of senescence in leaf cells of intact plants due to low or high sugar levels? J. Exp. Bot. 2008, 57, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Quiles, D.; Tovar-Sánchez, A. Sunscreens as a source of hydrogen peroxide production in coastal waters. Environ. Sci. Technol. 2014, 48, 9037–9042. [Google Scholar] [CrossRef] [Green Version]
- Raniello, R.; Lorenti, M.; Brunet, C.; Buia, M.C. Photosynthetic plasticity of an invasive variety of Caulerpa racemosa in a coastal Mediterranean area: Light harvesting capacity and seasonal acclimation. Mar. Ecol. Prog. Ser. 2004. [Google Scholar] [CrossRef] [Green Version]
- Uragami, C.; Galzerano, D.; Gall, A.; Shigematsu, Y.; Meisterhans, M.; Oka, N.; Iha, M.; Fujii, R.; Robert, B.; Hashimoto, H. Light-dependent conformational change of neoxanthin in a siphonous green alga, Codium intricatum, revealed by Raman spectroscopy. Photosynth. Res. 2014, 121, 69–77. [Google Scholar] [CrossRef]
- Sugawara, T.; Ganesan, P.; Li, Z.; Manabe, Y.; Hirata, T. Siphonaxanthin, a green algal carotenoid, as a novel functional compound. Mar. Drugs 2014, 12, 3660–3668. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, G.I.; Baker, N.R.; Woolhouse, H.W. Changes in chlorophyll content and organization during senescence of the primary leaves of Phaseolus vulgaris L. In relation to photosynthetic electron transport. J. Exp. Bot. 1981, 32, 1009–1020. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Proc. Biochim. Biophys. Acta Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Lin, X.M. Effects of lutein and zeaxanthin on aspects of eye health. J. Sci. Food Agric. 2010, 90, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Galasso, C.; Gentile, A.; Orefice, I.; Ianora, A.; Bruno, A.; Noonan, D.M.; Sansone, C.; Albini, A.; Brunet, C. Microalgal derivatives as potential nutraceutical and food supplements for human health: A focus on cancer prevention and interception. Nutrients 2019, 11, 1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricketts, T.R. The structures of siphonein and siphonaxanthin from Codium fragile. Phytochemistry 1971, 10, 155–160. [Google Scholar] [CrossRef]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a marine carotenoid from green algae, effectively induces apoptosis in human leukemia (HL-60) cells. Biochim. Biophys. Acta 2011, 1810, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, P.; Matsubara, K.; Ohkubo, T.; Tanaka, Y.; Noda, K.; Sugawara, T.; Hirata, T. Anti-angiogenic effect of siphonaxanthin from green alga, Codium fragile. Phytomedicine 2010, 17, 1140–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.-S.; Noda, K.; Fujita, E.; Manabe, Y.; Hirata, T.; Sugawara, T. The Green Algal Carotenoid Siphonaxanthin Inhibits Adipogenesis in 3T3-L1 Preadipocytes and the Accumulation of Lipids in White Adipose Tissue of KK-Ay Mice. J. Nutr. 2015, 145, 490–498. [Google Scholar] [CrossRef] [Green Version]
- Izumi, M.; Ishida, H. The changes of leaf carbohydrate contents as a regulator of autophagic degradation of chloroplasts via rubisco-containing bodies during leaf senescence. Plant Signal. Behav. 2011, 6, 685–687. [Google Scholar] [CrossRef] [Green Version]
- Leone, A.; Roca, M.S.; Ciardiello, C.; Costantini, S.; Budillon, A. Oxidative Stress Gene Expression Profile Correlates with Cancer Patient Poor Prognosis: Identification of Crucial Pathways Might Select Novel Therapeutic Approaches. Oxid. Med. Cell Longev. 2017, 2597581. [Google Scholar] [CrossRef] [Green Version]
- Touyz, R.M.; Anagnostopoulou, A.; Rios, F.; Montezano, A.C.; Camargo, L.L. NOX5: Molecular biology and pathophysiology. Exp. Physiol. 2019, 104, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Taso, O.V.; Philippou, A.; Moustogiannis, A.; Zevolis, E.; Koutsilieris, M. Lipid peroxidation products and their role in neurodegenerative diseases. Ann. Res. Hosp. 2019, 3. [Google Scholar] [CrossRef]
- Sansone, C.; Galasso, C.; Orefice, I.; Nuzzo, G.; Luongo, E.; Cutignano, A.; Romano, G.; Brunet, C.; Fontana, A.; Esposito, F.; et al. The green microalga Tetraselmis suecica reduces oxidative stress and induces repairing mechanisms in human cells. Sci. Rep. 2017, 7, 41215. [Google Scholar] [CrossRef] [PubMed]
- Smerilli, A.; Balzano, S.; Maselli, M.; Blasio, M.; Orefice, I.; Galasso, C.; Sansone, C.; Brunet, C. Antioxidant and photoprotection networking in the coastal diatom skeletonema marinoi. Antioxidants 2019, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Hartree, E.F. Determination of protein: A modification of the lowry method that gives a linear photometric response. Anal. Biochem. 1972, 48, 422–427. [Google Scholar] [CrossRef]
- Rice, D. The Detritus Nitrogen Problem: New Observations and Perspectives from Organic Geochemistry. Mar. Ecol. Prog. Ser. 1982, 9, 153–162. [Google Scholar] [CrossRef]
- Gerchakov, S.M.; Hatcher, P.G. Improved Technique for Analysis of Carbohydrates in Sediments. Limnol. Oceanogr. 1972, 17, 938–943. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37. [Google Scholar] [CrossRef] [Green Version]
- Marsh, J.B.; Weinstein, D.B. Simple charring method for determination of lipids. J. Lipid Res. 1966, 7, 574–576. [Google Scholar] [CrossRef]
- Olshansky, S.J.; Hayflick, L. The Role of the WI-38 Cell Strain in Saving Lives and Reducing Morbidity. AIMS Public Health 2017, 4, 127–138. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Carbohydrates | Lipids | |
|---|---|---|---|
| Young leaves | 21.3 ± 0.5 | 19.0 ± 0.6 | 5.8 ± 0.1 |
| Old leaves | 8.2 ± 0.6 | 10.9 ± 1.1 | 1.6 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sansone, C.; Galasso, C.; Lo Martire, M.; Fernández, T.V.; Musco, L.; Dell’Anno, A.; Bruno, A.; Noonan, D.M.; Albini, A.; Brunet, C. In Vitro Evaluation of Antioxidant Potential of the Invasive Seagrass Halophila stipulacea. Mar. Drugs 2021, 19, 37. https://doi.org/10.3390/md19010037
Sansone C, Galasso C, Lo Martire M, Fernández TV, Musco L, Dell’Anno A, Bruno A, Noonan DM, Albini A, Brunet C. In Vitro Evaluation of Antioxidant Potential of the Invasive Seagrass Halophila stipulacea. Marine Drugs. 2021; 19(1):37. https://doi.org/10.3390/md19010037
Chicago/Turabian StyleSansone, Clementina, Christian Galasso, Marco Lo Martire, Tomás Vega Fernández, Luigi Musco, Antonio Dell’Anno, Antonino Bruno, Douglas M. Noonan, Adriana Albini, and Christophe Brunet. 2021. "In Vitro Evaluation of Antioxidant Potential of the Invasive Seagrass Halophila stipulacea" Marine Drugs 19, no. 1: 37. https://doi.org/10.3390/md19010037