
Study on the Mechanism of the Blood-Glucose-Lowering Effect of Collagen Peptides from Sturgeon By-Products

 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
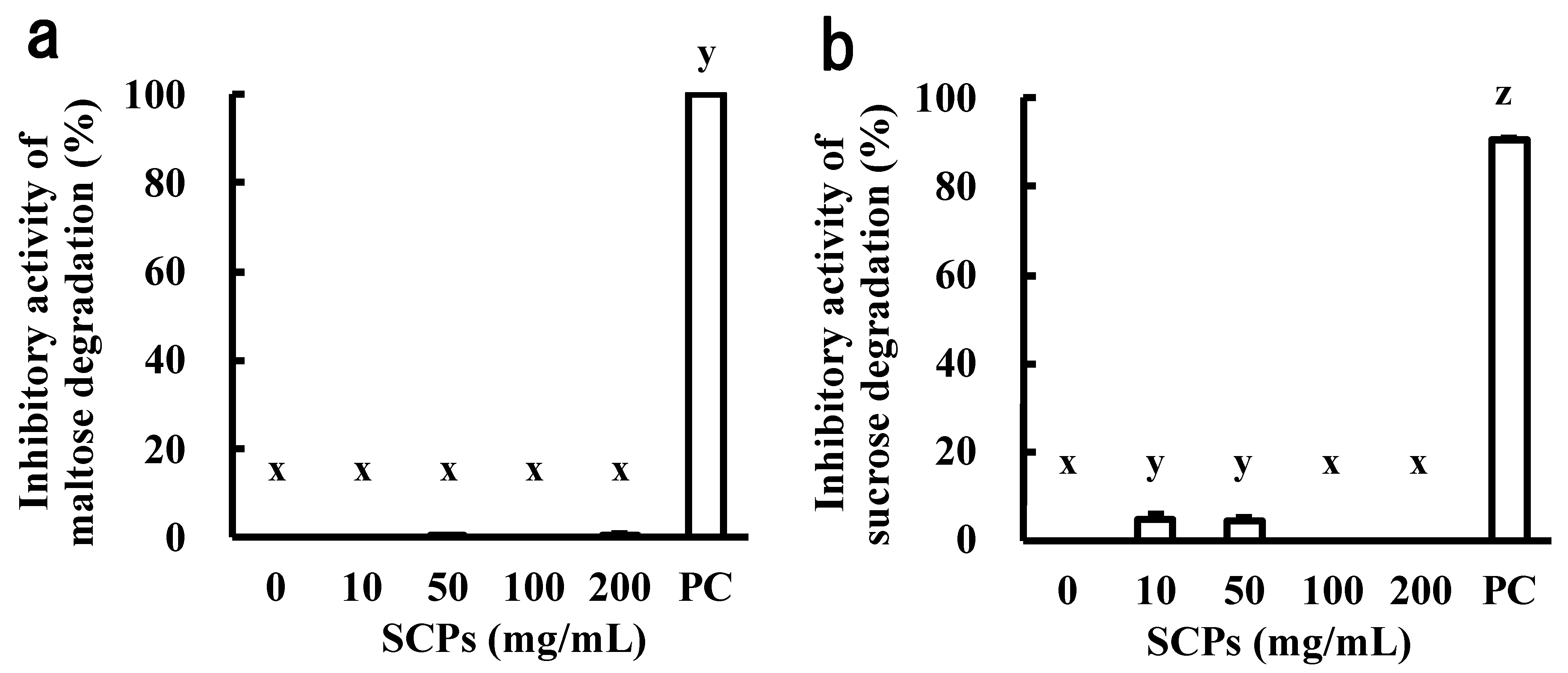
2.1. Effect of the SCPs on α-Glucosidase and Invertase Abilities
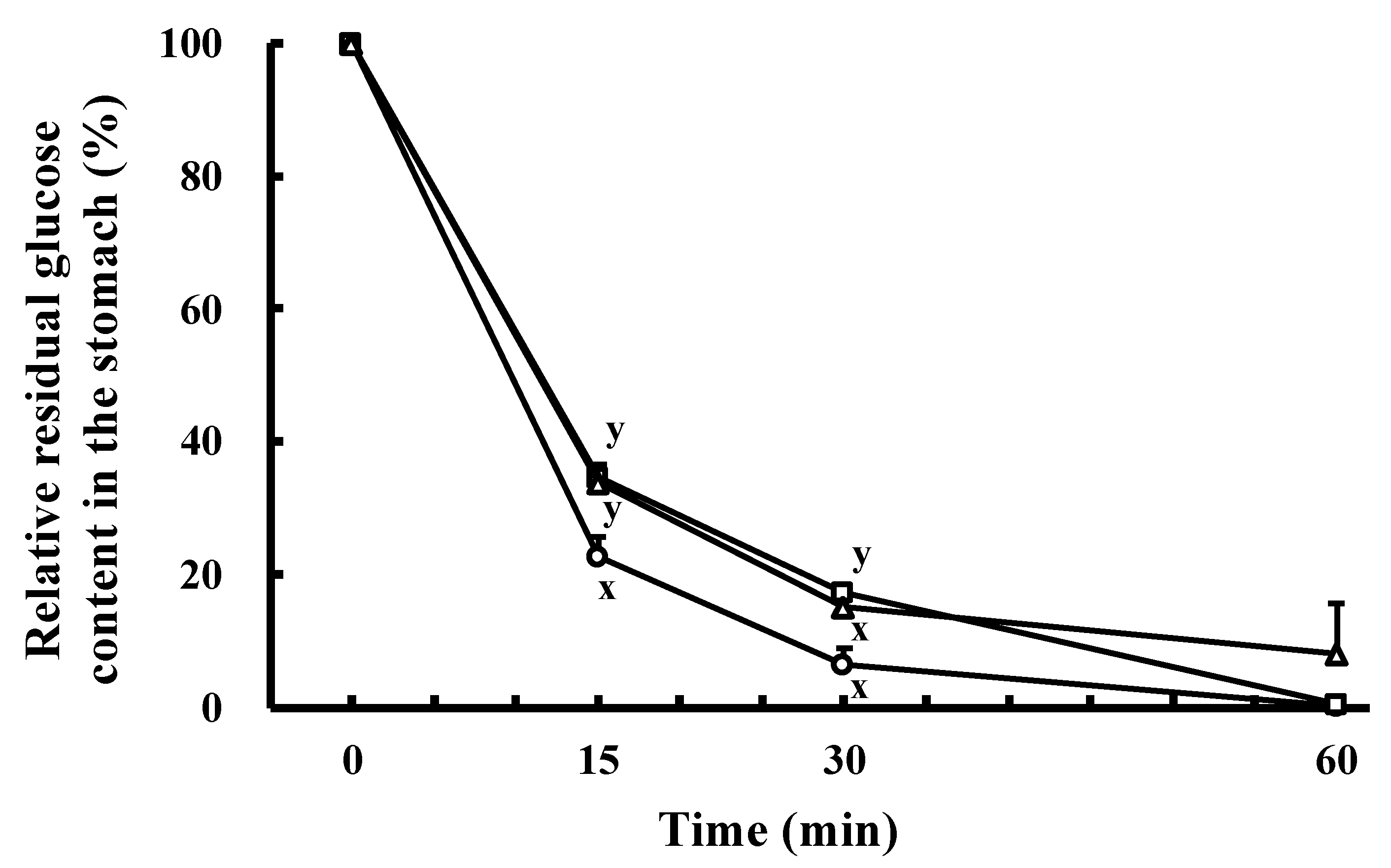
2.2. Effect of the Orally Administrated SCPs on Residual Glucose Content in the Stomach in the Oral Glucose Tolerance Test (OGTT)
2.3. Effect of the Orally Administrated SCPs on Glucose Absorption in the Small Intestine in the OGTT
2.4. Effect of the Orally Administrated SCPs on Blood Glucose and Insulin Levels in the Intraperitoneal Glucose Tolerance Test (IPGTT)
2.5. Effect of the Intravenously Injected SCPs on Blood Glucose and Insulin Levels in the IPGTT
2.6. Effect of the SCPs on DPP-IV Activity
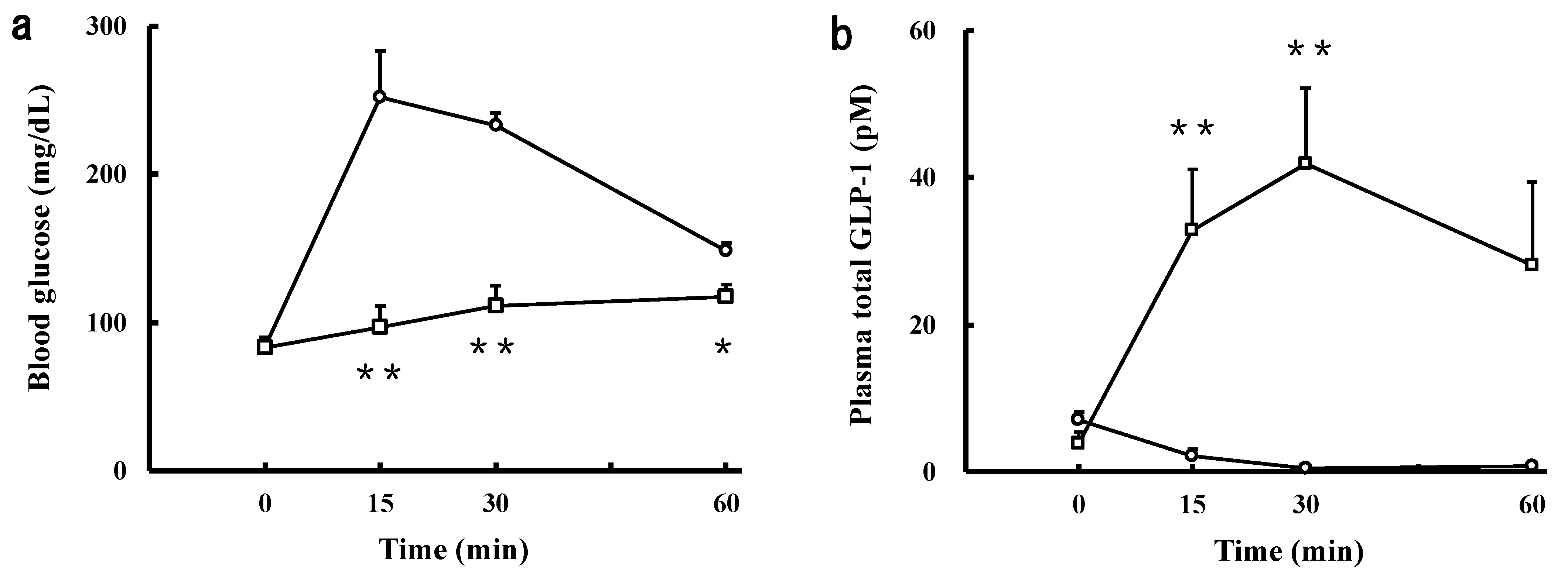
2.7. Effect of the Intravenously Injected SCPs on Blood Glucose and GLP-1 Levels in the IPGTT
3. Materials and Methods
3.1. Materials
3.2. α-Glucosidase and Invertase Inhibitory Assays
3.3. DPP-IV Inhibitory Assay
3.4. Measurement of Blood Glucose in OGTT
3.5. Measurement of Glucose in the Stomach in OGTT
3.6. Measurement of Glucose Content in Excrements in OGTT
3.7. Measurement of Blood Glucose and Insulin Levels in IPGTT
3.7.1. Oral Administration of the SCPs
3.7.2. Intravenous Injection of the SCPs
3.8. Measurement of Insulin Levels
3.9. Measurement of Total GLP-1 Levels
3.10. Measurement of Endotoxin Concentration
3.11. Preparation of Type 1 Diabetes Model Rats
3.12. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieffer, T.J.; McIntosh, C.H.; Pederson, R.A. Degradation of glucose-dependent insulinotropic polypeptide and truncated glu-cagon-like peptide 1 in vitro and in vivo by dipeptidyl peptidase IV. Endocrinology 1995, 136, 3585–3596. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Masuda, S.; Yamakage, H.; Inoue, T.; Ohue-Kitano, R.; Yokota, S.; Kusakabe, T.; Wada, H.; Sanada, K.; Ishii, K.; et al. Role of serum myostatin in the association between hyperinsulinemia and muscle atrophy in Japanese obese patients. Diabetes Res. Clin. Pract. 2018, 142, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Fact Sheet: Diabetes. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 7 October 2021).
- Hirata, Y.; Nomura, K.; Senga, Y.; Okada, Y.; Kobayashi, K.; Okamoto, S.; Minokoshi, Y.; Imamura, M.; Takeda, S.; Hosooka, T.; et al. Hyperglycemia induces skeletal muscle atrophy via a WWP1/KLF15 axis. JCI Insight 2019, 4, e124952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanri, A.; Mizoue, T.; Noda, M.; Takahashi, Y.; Matsushita, Y.; Poudel-Tandukar, K.; Kato, M.; Oba, S.; Inoue, M.; Tsugane, S.; et al. Fish intake and type 2 diabetes in Japanese men and women: The Japan Public Health Center-based Pro-spective Study. Am. J. Clin. Nutr. 2011, 94, 884–891. [Google Scholar] [CrossRef] [Green Version]
- Liaset, B.; Øyen, J.; Jacques, H.; Kristiansen, K.; Madsen, L. Seafood intake and the development of obesity, insulin resistance and type 2 diabetes. Nutr. Res. Rev. 2019, 32, 146–167. [Google Scholar] [CrossRef] [Green Version]
- Kishimura, H.; Nagai, Y.; Fukumorita, K.; Adachi, K.; Chiba, S.; Nakajima, S.; Saeki, H.; Klomklao, S.; Nalinanon, S.; Benjakul, S.; et al. Acid- and heat-stable trypsin inhibitory peptide from the viscera of japanese common squid (Todarodes pacificus). J. Food Biochem. 2010, 34, 748–763. [Google Scholar] [CrossRef]
- Kishimura, H.; Fukumorita, K.; Adachi, K.; Chiba, S.; Nagai, Y.; Katayama, S.; Nakajima, S.; Saeki, H. A trypsin inhibitor in the viscera of japanese common squid (Todarodes pacificus) elicits insulinotropic effects in diabetic GK rats hypoglycemic effects of squid viscera. J. Food Biochem. 2012, 36, 93–98. [Google Scholar] [CrossRef]
- Adachi, K.; Fukumorita, K.; Araki, M.; Zaima, N.; Yang, Z.-H.; Chiba, S.; Kishimura, H.; Saeki, H. Transcriptome Analysis of the Duodenum, Pancreas, Liver, and Muscle from Diabetic Goto-Kakizaki Rats Fed a Trypsin Inhibitor Derived from Squid Viscera. J. Agric. Food Chem. 2012, 60, 5540–5546. [Google Scholar] [CrossRef]
- Billard, R.; Lecointre, G. Biology and conservation of sturgeon and paddlefish. Rev. Fish Biol. Fish. 2000, 10, 355–392. [Google Scholar] [CrossRef]
- WWF. Saving Sturgeons—A Grobal Report on Their Status and Suggested Conservation Strategy; WWF: Gland, Switzerland, 2016; pp. 2–3. [Google Scholar]
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Bronzi, P.; Rosenthal, H.; Gessner, J. Grobal sturgeon aquaculture production: An overview. J. Appl. Ichthyol. 2011, 27, 169–175. [Google Scholar] [CrossRef]
- Takahashi, S. Sturgeon conservation and the role of Japan. In The State of Wildlife Trade in Japan; Ishihara, A., Traffic East Asia-Japan, Eds.; TRAFFIC East Asia-Japan: Tokyo, Japan, 2010; pp. 42–47. [Google Scholar]
- Zhang, X.; Ookawa, M.; Tan, Y.; Ura, K.; Adachi, S.; Takagi, Y. Biochemical characterisation and assessment of fibril-forming ability of collagens extracted from Bester sturgeon Huso huso × Acipenser ruthenus. Food Chem. 2014, 160, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Sasaoka, Y.; Kishimura, H.; Adachi, S.; Takagi, Y. Collagen peptides derived from the triple helical region of sturgeon collagen improve glucose tolerance in normal mice. J. Food Biochem. 2017, 42, e12478. [Google Scholar] [CrossRef] [Green Version]
- Dooley, C.P.; Reznick, J.B.; Valenzuela, J.E. Variations in Gastric and Duodenal Motility During Gastric Emptying of Liquid Meals in Humans. Gastroenterology 1984, 87, 1114–1119. [Google Scholar] [CrossRef]
- Fox, M.; Georgi, G.; Boehm, G.; Menne, D.; Fried, M.; Thumshirn, M. Dietary protein precipitation properties have effects on gastric emptying in healthy volunteers. Clin. Nutr. 2004, 23, 641–646. [Google Scholar] [CrossRef]
- Karamanlis, A.; Chaikomin, R.; Doran, S.; Bellon, M.; Bartholomeusz, F.D.; Wishart, J.M.; Jones, K.; Horowitz, M.; Rayner, C.K. Effects of protein on glycemic and incretin responses and gastric emptying after oral glucose in healthy subjects. Am. J. Clin. Nutr. 2007, 86, 1364–1368. [Google Scholar] [CrossRef]
- Iba, Y.; Yokoi, K.; Eitoku, I.; Goto, M.; Koizumi, S.; Sugihara, F.; Oyama, H.; Yoshimoto, T. Oral Administration of Collagen Hydrolysates Improves Glucose Tolerance in Normal Mice Through GLP-1-Dependent and GLP-1-Independent Mechanisms. J. Med. Food 2016, 19, 836–843. [Google Scholar] [CrossRef]
- Imeryüz, N.; Yegen, B.; Bozkurt, A.; Coşkun, T.; Villanueva-Peñacarrillo, M.L.; Ulusoy, N.B. Glucagon-like peptide-1 inhibits gastric emptying via vagal afferent-mediated central mechanisms. Am. J. Physiol. Liver Physiol. 1997, 273, G920–G927. [Google Scholar] [CrossRef]
- Alemán, A.; Giménez, B.; Montero, P.; Gómez-Guillén, M. Antioxidant activity of several marine skin gelatins. LWT 2011, 44, 407–413. [Google Scholar] [CrossRef]
- Abdollahi, M.; Rezaei, M.; Jafarpour, A.; Undeland, I. Sequential extraction of gel-forming proteins, collagen and collagen hydrolysate from gutted silver carp (Hypophthalmichthys molitrix), a biorefinery approach. Food Chem. 2018, 242, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Iwai, K.; Hasegawa, T.; Taguchi, Y.; Morimatsu, F.; Sato, K.; Nakamura, Y.; Higashi, A.; Kido, Y.; Nakabo, Y.; Ohtsuki, K. Identification of Food-Derived Collagen Peptides in Human Blood after Oral Ingestion of Gelatin Hydrolysates. J. Agric. Food Chem. 2005, 53, 6531–6536. [Google Scholar] [CrossRef] [PubMed]
- Meilman, E.; Urivetzky, M.M.; Rapoport, C.M. Urinary Hydroxyproline Peptides. J. Clin. Investig. 1963, 42, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Baller, B.; Meier, J.J. Gastric Inhibitory Polypeptide and Glucagon-Like Peptide-1 in the Pathogenesis of Type 2 Diabetes. Diabetes 2004, 53, S190–S196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Aiello, G.; Bollati, C.; Bartolomei, M.; Arnoldi, A.; Lammi, C. Phycobiliproteins from Arthrospira Platensis (Spirulina): A New Source of Peptides with Dipeptidyl Peptidase-IV Inhibitory Activity. Nutrients 2020, 12, 794. [Google Scholar] [CrossRef] [Green Version]
- Li-Chan, E.C.; Hunag, S.L.; Jao, C.L.; Ho, K.P.; Hsu, K.C. Peptides derived from Atlantic salmon skin gelatin as dipep-tidyl-peptidase IV inhibitors. J. Agric. Food Chem. 2012, 60, 973–978. [Google Scholar] [CrossRef]
- Sila, A.; Martinez-Alvarez, O.; Haddar, A.; Gómez-Guillén, M.C.; Nasri, M.; Montero, M.P.; Bougatef, A. Recovery, viscoelastic and functional properties of Barbel skin gelatine: Investigation of anti-DPP-IV and anti-prolyl endopeptidase activities of generated gelatine polypeptides. Food Chem. 2015, 168, 478–486. [Google Scholar] [CrossRef] [Green Version]
- Baggio, L.L.; Drucker, D.J. Biology of Incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Mortensen, K.; Christensen, L.L.; Holst, J.J.; Orskov, C. GLP-1 and GIP are colocalized in a subset of endocrine cells in the small intestine. Regul. Pept. 2003, 114, 189–196. [Google Scholar] [CrossRef]
- Theodorakis, M.J.; Carlson, O.; Michopoulos, S.; Doyle, M.E.; Juhaszova, M.; Petraki, K.; Egan, J.M. Human duodenal entero-endocrine cells: Source of both incretin peptides, GLP-1 and GIP. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E550–E559. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.H.; Wang, T.Y.; Hung, C.C.; Chen, M.C.; Hsu, K.C. Improvement of glycemic control in streptozotocin-induced diabetic rats by Atlantic salmon skin gelatin hydrolysate as the dipeptidyl-peptidase IV inhibitor. Food Funct. 2015, 6, 1887–1892. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.; FitzGerald, R.J. Atlantic salmon (Salmo salar) co-product-derived protein hydrolysates: A source of antidiabetic peptides. Food Res. Int. 2018, 106, 598–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, T. α-Glucosidase inhibitory activity of a 70% methanol extract from Ezoishige (Pelvetia babingtonii de Toni) and its effect on the elevation of blood glucose level in rats. Biosci. Biotechnol. Biochem. 2002, 66, 1552–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lots | Endotoxin Concentrations |
|---|---|
| SCPs-1 | 0.014 ng/mg (0.112 endotoxin unit/mg) |
| SCPs-2 | 0.100 ng/mg (0.800 endotoxin unit/mg) |
| Peptides | Peptides MW (Da) | Enzymes Used for Preparation | IC50 Values | |
|---|---|---|---|---|
| (µg/mU) | (μmol/mU) | |||
| SCPs | 1400 [18] | Papain | 934 | 0.667 |
| Collagen peptides (Atlantic salmon skin) [30] | <1000 | Flavourzyme | 270 | 0.270 |
| Gly-Pro-Ala-Glu [30] | 372.4 | (Synthetic peptide) | 0.924 | 0.248 × 10−2 |
| Gly-Pro-Gly-Ala [30] | 300.4 | (Synthetic peptide) | 1.26 | 0.210 × 10−2 |
| Collagen peptides (tilapia scale) [22] | - | - | 616 | - |
| Collagen peptides (carp skin) [31] | 2935 | Esperase | 425 | 0.145 |
| 2322 | Savinase | 521 | 0.224 | |
| 1995 | Alcalase | 504 | 0.253 | |
| 3925 | Trypsin | 479 | 0.122 | |
| 5797 | Izyme G | 714 | 0.123 | |
| 2325 | Protamex | 467 | 0.201 | |
| 7878 | Neutrase | 529 | 0.067 | |
| 15,053 | Peptidase | 527 | 0.035 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaoka, Y.; Takagi, T.; Michiba, S.; Yamamoto, Y.; Kumagai, Y.; Kishimura, H. Study on the Mechanism of the Blood-Glucose-Lowering Effect of Collagen Peptides from Sturgeon By-Products. Mar. Drugs 2021, 19, 584. https://doi.org/10.3390/md19100584
Sasaoka Y, Takagi T, Michiba S, Yamamoto Y, Kumagai Y, Kishimura H. Study on the Mechanism of the Blood-Glucose-Lowering Effect of Collagen Peptides from Sturgeon By-Products. Marine Drugs. 2021; 19(10):584. https://doi.org/10.3390/md19100584
Chicago/Turabian StyleSasaoka, Yukiho, Taichi Takagi, Shunta Michiba, Yohei Yamamoto, Yuya Kumagai, and Hideki Kishimura. 2021. "Study on the Mechanism of the Blood-Glucose-Lowering Effect of Collagen Peptides from Sturgeon By-Products" Marine Drugs 19, no. 10: 584. https://doi.org/10.3390/md19100584
APA StyleSasaoka, Y., Takagi, T., Michiba, S., Yamamoto, Y., Kumagai, Y., & Kishimura, H. (2021). Study on the Mechanism of the Blood-Glucose-Lowering Effect of Collagen Peptides from Sturgeon By-Products. Marine Drugs, 19(10), 584. https://doi.org/10.3390/md19100584