
Unravelling the Anti-Inflammatory and Antioxidant Potential of the Marine Sponge Cliona celata from the Portuguese Coastline

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Biological Activities of Cliona celata Crude Extracts
2.1.1. Antioxidant Activity
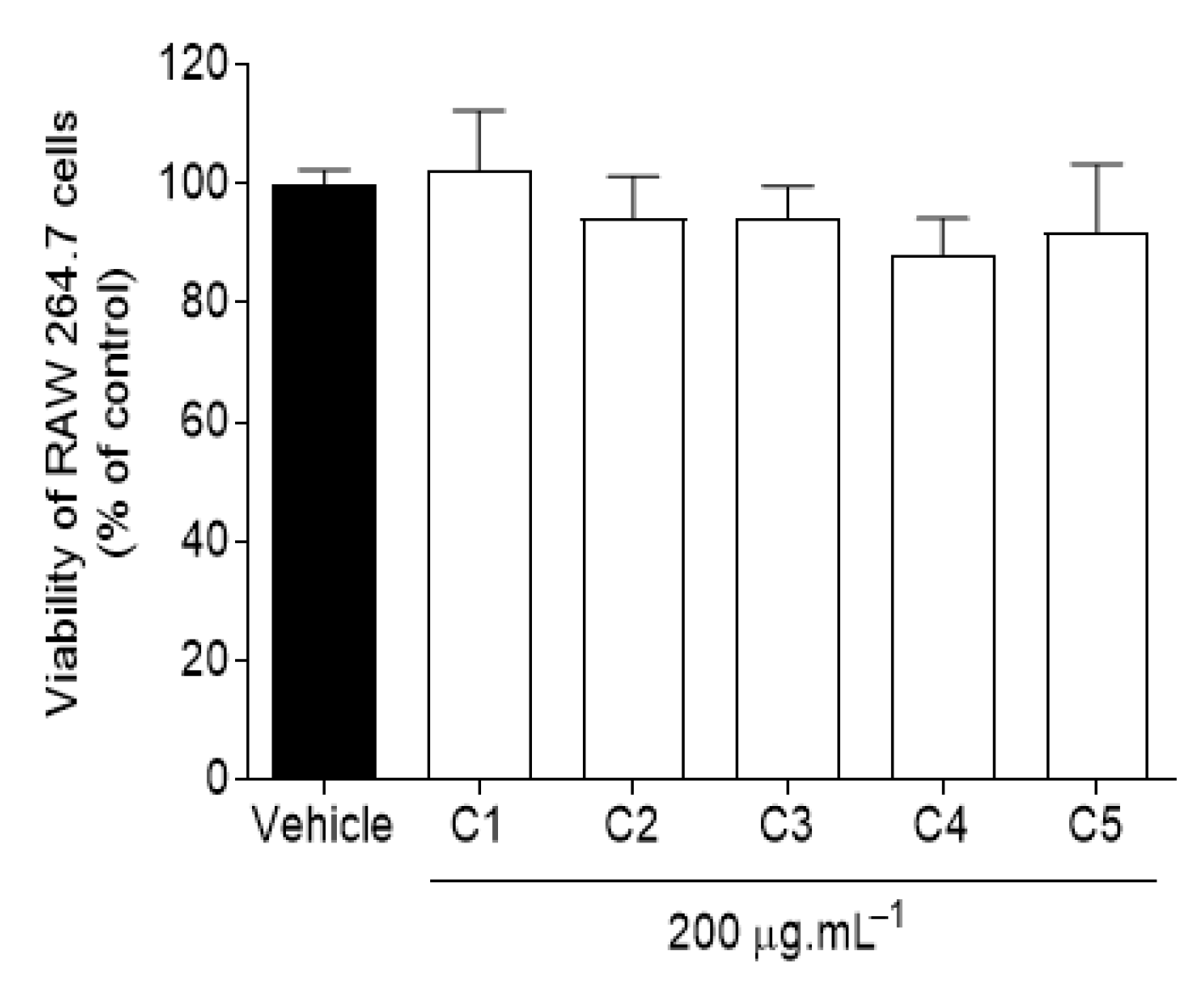
2.1.2. Cytotoxic Activity in RAW 264.7 Macrophages
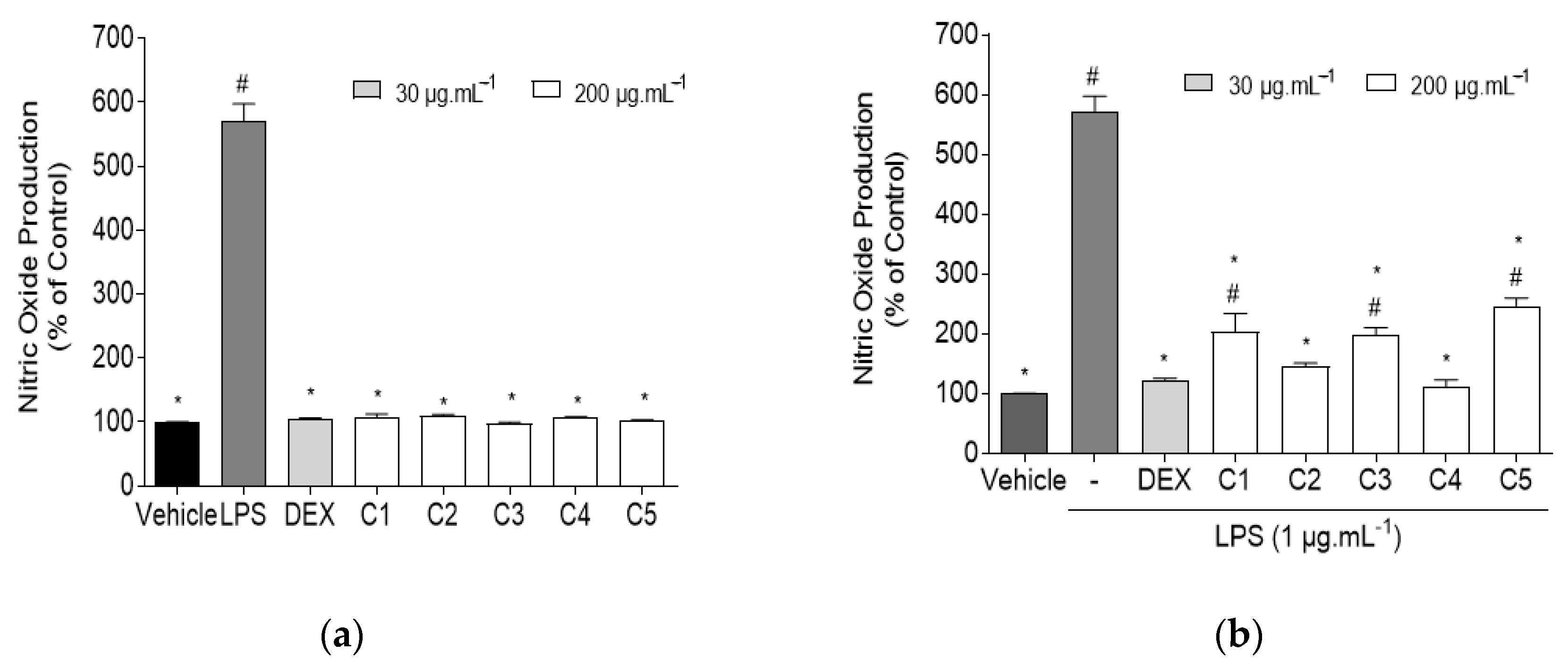
2.1.3. Quantification of Nitric Oxide (NO) in RAW 267.4 Cells
2.2. Evaluation of Biological Activities of Cliona celata Fractions
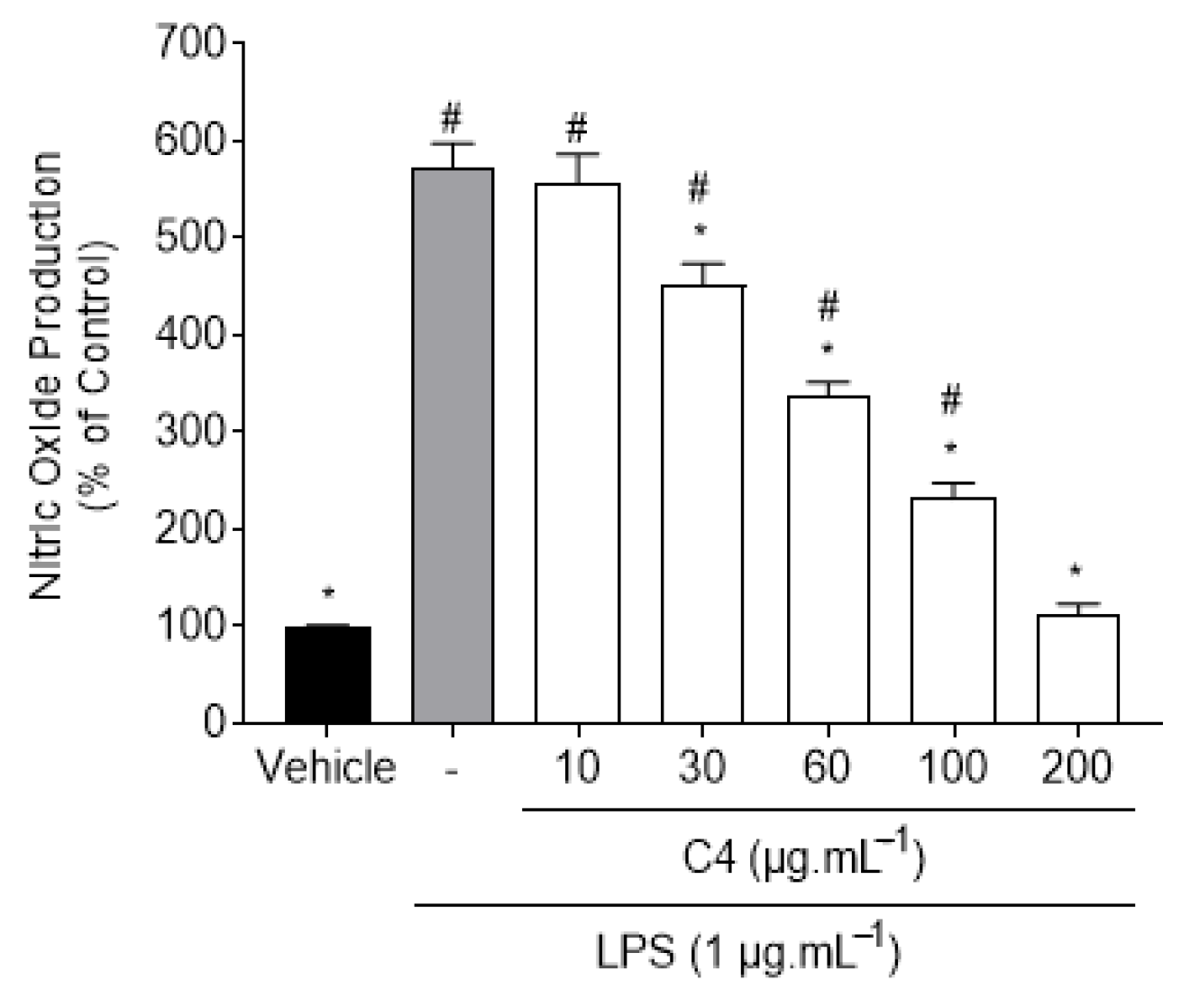
2.2.1. Cytotoxic Activity of C4 Fractions in RAW 264.7 Macrophages
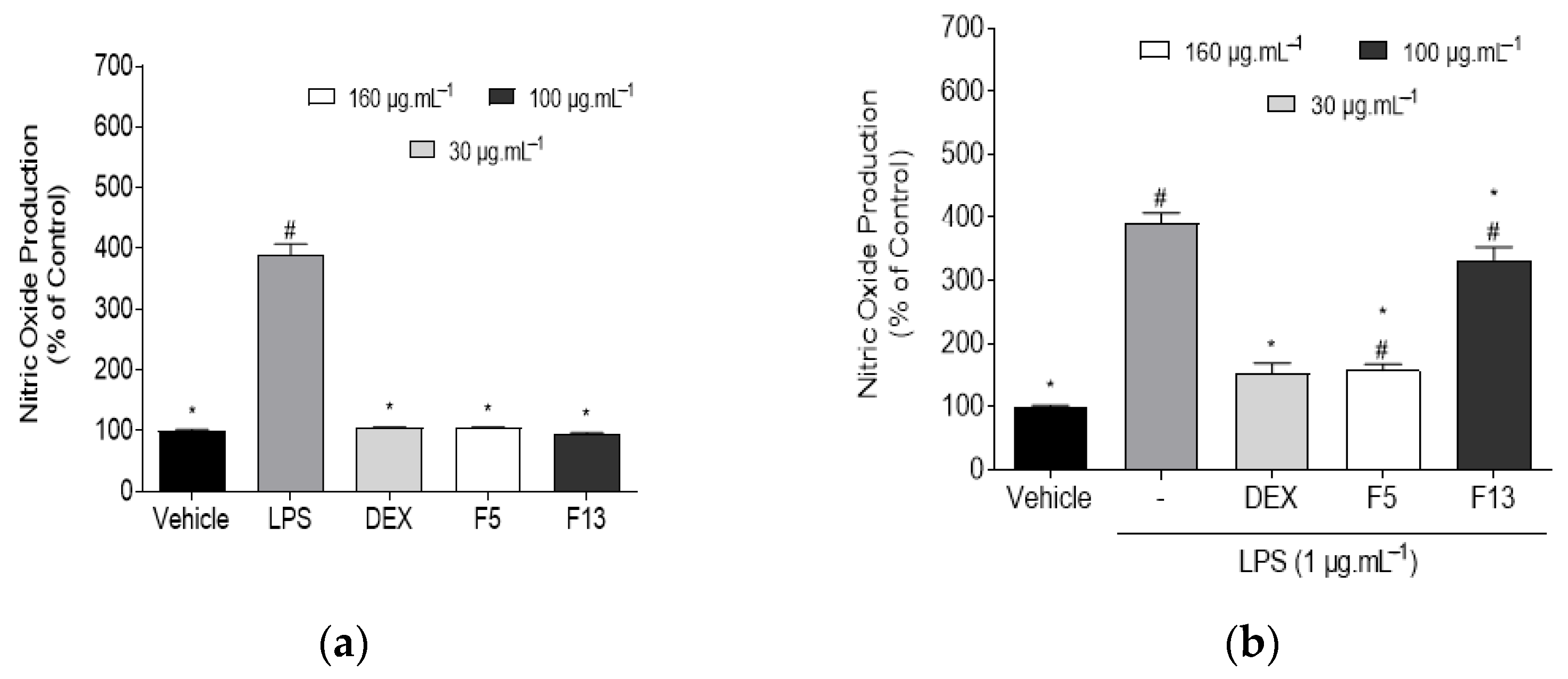
2.2.2. Effect of C4 Fractions (F5 and F13) on Nitric Oxide (NO) Levels in Normal and LPS-Induced RAW 267.4 Cells
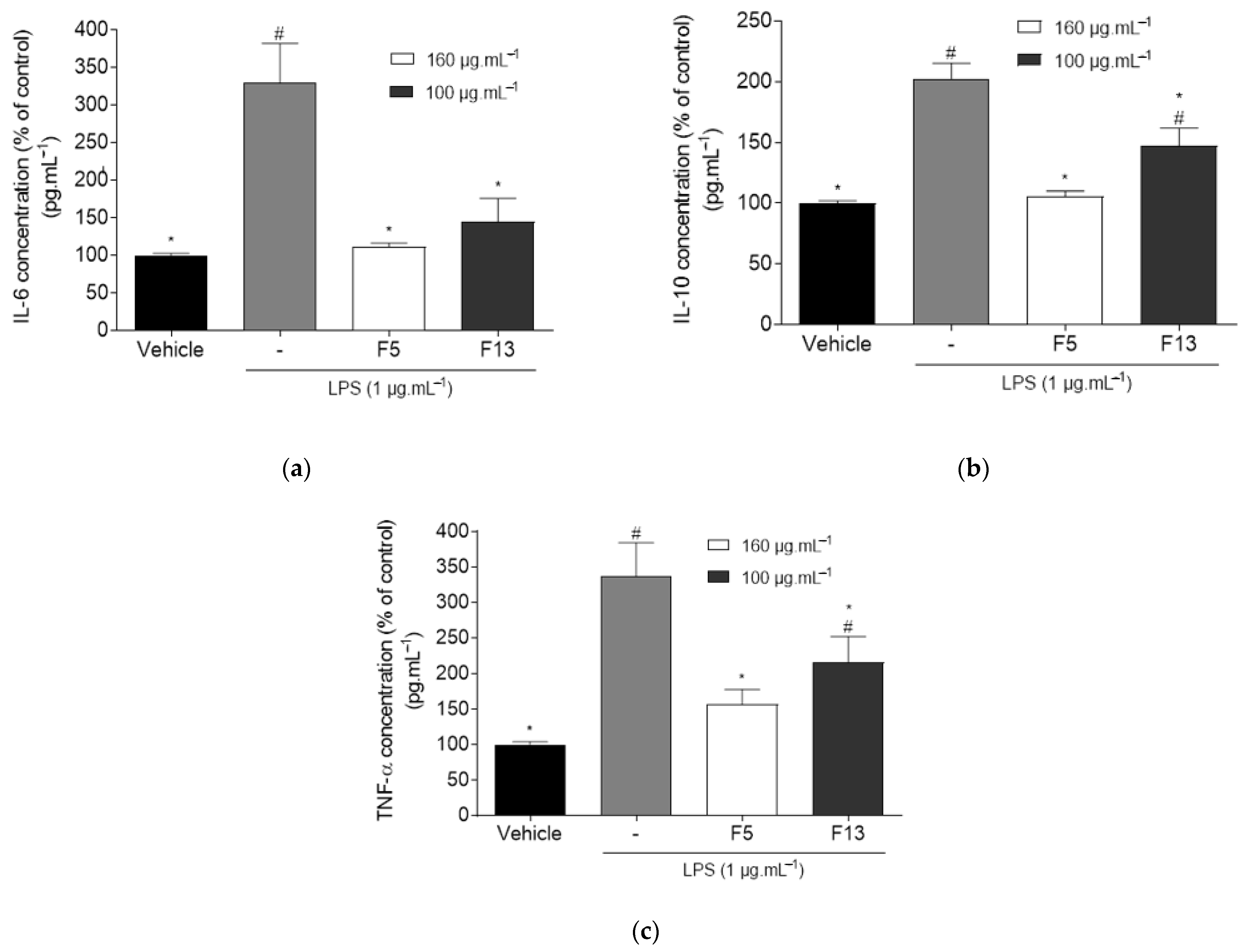
2.2.3. Effect of C4 Fractions (F5 and F13) on the Levels of TNF-α, IL-6 and IL-10 on LPS-Induced Inflammation on RAW 264.7 Macrophages
2.3. Chemical Characterization by GC-MS Analysis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
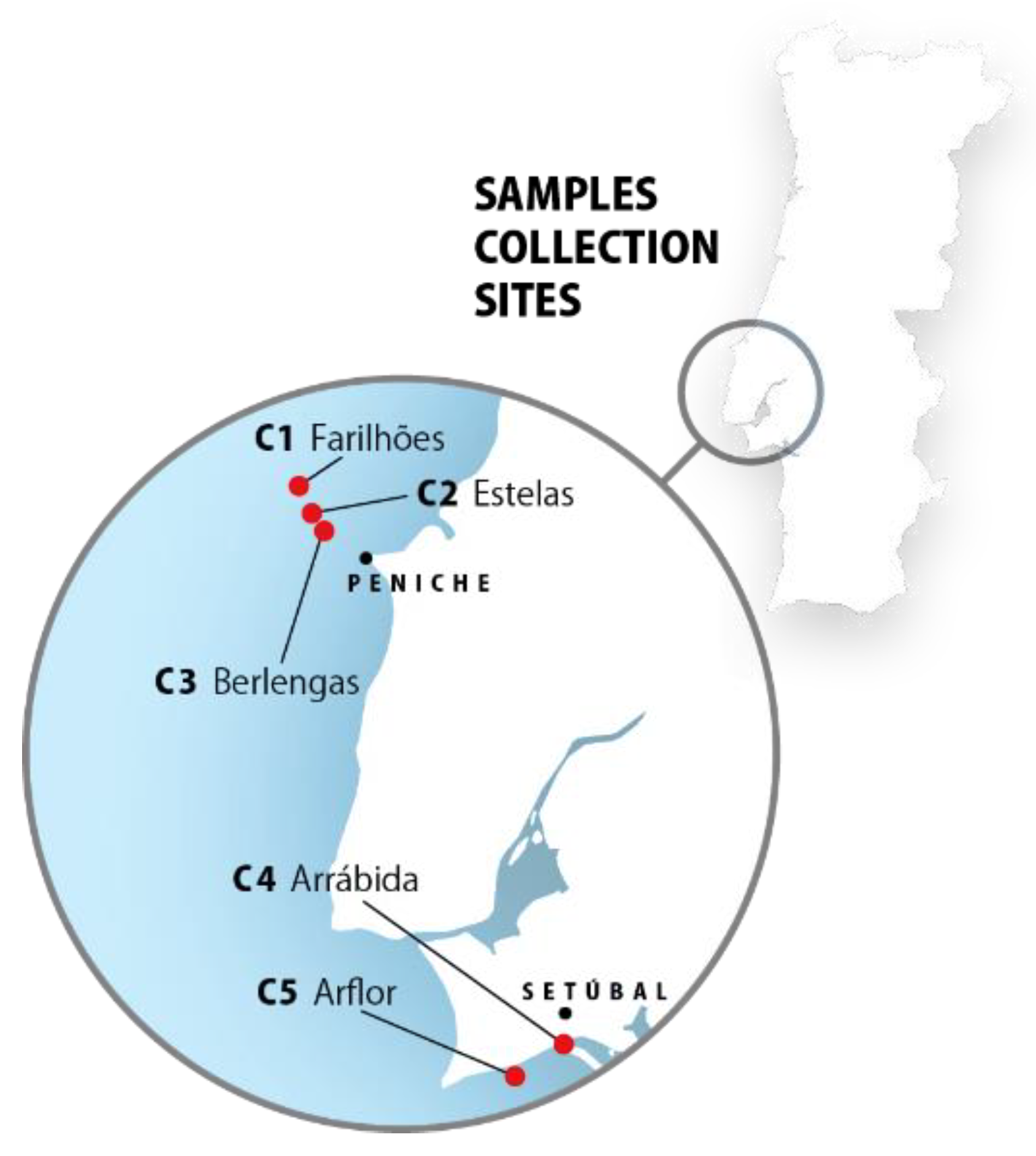
4.2. Marine Sponge Collection
4.3. Marine Sponge Extraction
4.4. Sponge Extracts Fractionation
4.5. Evaluation of Biological Activities of Cliona celata Crude Extracts and Fractions
4.5.1. Antioxidant Activity
2,2-Diphenyl-1-Picrylhydrazyl (DPPH) Radical Scavenging Activity
Ferric Reducing Antioxidant Power (FRAP)
Oxygen Radical Absorbance Capacity (ORAC)
4.5.2. Evaluation of Biological Activities of Cliona celata Crude Extracts and Fractions on In Vitro Cellular Models
Cell Culture Maintenance
Cytotoxic Activity in RAW 264.7 Macrophages
Quantification of Nitric Oxide (NO) on RAW 267.4 Cells
Quantification of the Levels of TNF-α, IL-6, and IL-10 on LPS-Induced RAW 264.7 Macrophages
4.6. Chemical Characterization by GC-MS Analysis
4.7. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Du, C.; Bhatia, M.; Tang, S.C.W.; Zhang, M.; Steiner, T. Mediators of Inflammation: Inflammation in Cancer, Chronic Diseases, and Wound Healing. Mediators Inflamm. 2015, 2015, 570653. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Duque, G.A.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Denis Alexander, H.; Ross, O.A. Age and age-related diseases: Role of inflammation triggers and cytokines. Front. Immunol. 2018, 9, 586. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Hunter, P. The inflammation theory of disease. EMBO Rep. 2012, 13, 968–970. [Google Scholar] [CrossRef] [Green Version]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [Green Version]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.H.; Ma, J.N.; Park, J.H.; Jung, H.W.; Park, Y.K. Anti-inflammatory and antioxidant effects of MOK, a polyherbal extract, on lipopolysaccharide-stimulated RAW 264.7 macrophages. Int. J. Mol. Med. 2018, 43, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- El-Demerdash, F.M.; Tousson, E.M.; Kurzepa, J.; Habib, S.L. Xenobiotics, oxidative stress, and antioxidants. Oxidative Med. Cell. Longev. 2018, 2018, 9758951. [Google Scholar] [CrossRef] [Green Version]
- Stonik, V.A. Marine Natural Products: A Way to New Drugs. Acta Naturae 2009, 1, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [Green Version]
- Anjum, K.; Abbas, S.Q.; Shah, S.A.A.; Akhter, N.; Batool, S.; Hassan, S.S.U. Marine sponges as a drug treasure. Biomol. Ther. 2016, 24, 347–362. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K. Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 9783642539718. [Google Scholar]
- Yang, J.H.; Suh, S.J.; Lu, Y.; Li, X.; Lee, Y.K.; Chang, Y.C.; Na, M.K.; Choi, J.H.; Kim, C.H.; Son, J.K.; et al. Anti-inflammatory activity of ethylacetate fraction of Cliona celata. Immunopharmacol. Immunotoxicol. 2010, 33, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Keyzers, R.A.; Daoust, J.; Davies-Coleman, M.T.; Van Soest, R.; Balgi, R.; Donohue, E.; Roberge, M.; Andersen, R.J. Autophagy-modulating aminosteroids isolated from the sponge Cliona celata. Org. Lett. 2008, 10, 2959–2962. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C. Drug Discovery Handbook; John Wiley & Sons: Hoboken, NJ, USA, 2005; ISBN 0471213845. [Google Scholar]
- Hong, J. Natural product diversity and its role in chemical biology and drug discovery. Curr. Opin. Chem. Biol. 2011, 15, 350–354. [Google Scholar] [CrossRef] [Green Version]
- Seradj, H.; Moein, M.; Eskandari, M.; Maaref, F. Antioxidant Activity of Six Marine Sponges Collected from the Persian Gulf. Iran. J. Pharm. Sci. 2012, 8, 249–255. [Google Scholar]
- Shaaban, M.; Abd-Alla, H.I.; Hassan, A.Z.; Aly, H.F.; Ghani, M.A. Chemical characterization, antioxidant and inhibitory effects of some marine sponges against carbohydrate metabolizing enzymes. Org. Med. Chem. Lett. 2012, 2, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bary, K.; Elamraoui, B.; Bamhaoud, T. Chemical characterization of Cliona viridis: Sponge of Atlantic Moroccan Coast. Int. J. Innov. Sci. Res. 2016, 26, 14–22. [Google Scholar]
- Yuan, G.; Wahlqvist, M.L.; He, G.; Yang, M.; Li, D. Natural products and anti-inflammatory activity. Asia Pac. J. Clin. Nutr. 2006, 15, 143–152. [Google Scholar]
- Fung, S.Y.; Sofiyev, V.; Schneiderman, J.; Hirschfeld, A.F.; Victor, R.E.; Woods, K.; Piotrowski, J.S.; Deshpande, R.; Li, S.C.; De Voogd, N.J.; et al. Unbiased screening of marine sponge extracts for antiinflammatory agents combined with chemical genomics identifies girolline as an inhibitor of protein synthesis. ACS Chem. Biol. 2013, 9, 247–257. [Google Scholar] [CrossRef] [Green Version]
- El-Demerdash, A.; Atanasov, A.G.; Horbanczuk, O.K.; Tammam, M.A.; Abdel-Mogib, M.; Hooper, J.N.A.; Sekeroglu, N.; Al-Mourabit, A.; Kijjoa, A. Chemical diversity and biological activities of marine sponges of the genus Suberea: A systematic review. Mar. Drugs 2019, 17, 115. [Google Scholar] [CrossRef] [Green Version]
- Koelman, L.; Pivovarova-Ramich, O.; Pfeiffer, A.F.H.; Grune, T.; Aleksandrova, K. Cytokines for evaluation of chronic inflammatory status in ageing research: Reliability and phenotypic characterisation. Immun. Ageing 2019, 16, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinne, R.W.; Stuhlmüller, B.; Burmester, G.R. Macrophages. In Rheumatoid Arthritis; Royal College of Physicians of London: London, UK, 2009; ISBN 9780323054751. [Google Scholar]
- Ravussin, E.; Smith, S.R. Role of the Adipocyte in Metabolism and Endocrine Function. In Endocrinology: Adult and Pediatric; Elsevier: Philadelphia, PA, USA, 2015; ISBN 9780323189071. [Google Scholar]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. Ther. 2006, 8, S3. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Il-6 in inflammation, Immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Barnes, P.J. Cytokine modulators as novel therapies for airway disease. Eur. Respir. J. 2001, 18, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, X.; Oskarsson, J.T.; Omarsdottir, S.; Freysdottir, J.; Hardardottir, I. Lipophilic fractions from the marine sponge Halichondria sitiens decrease secretion of pro-inflammatory cytokines by dendritic cells and decrease their ability to induce a Th1 type response by allogeneic CD4+ T cells. Pharm. Biol. 2017, 55, 2116–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costantini, S.; Romano, G.; Rusolo, F.; Capone, F.; Guerriero, E.; Colonna, G.; Ianora, A.; Ciliberto, G.; Costantini, M. Anti-Inflammatory Effects of a Methanol Extract from the Marine Sponge Geodia cydonium on the Human Breast Cancer MCF-7 Cell Line. Mediat. Inflamm. 2015, 2015, 204975. [Google Scholar] [CrossRef] [Green Version]
- Prabakaran, R.; Kumar, T.S.; Rao, M.V. GC-MS Analysis and In vitro Cytotoxicity Studies of Root Bark Exudates of Hardwickia binata Roxb. Am. J. Phytomed. Clin. Ther. 2014, 2, 723–733. [Google Scholar]
- Stonard, R.J.; Andersen, R.J. Celenamides A and B, Linear Peptide Alkaloids from the Sponge Cliona celata. J. Org. Chem. 1980, 45, 3687–3691. [Google Scholar] [CrossRef]
- Stonard, R.J.; Andersen, R.J. Linear peptide alkaloids from the sponge Cliona celata (Grant). Celenamides C and D. Can. J. Chem. 1980, 58, 2121–2126. [Google Scholar] [CrossRef]
- Lenis, L.A.; Nuñez, L.; Jiménez, C.; Riguera, R. Isonitenin and acetylhomoagmatine new metabolites from the sponges Spongia officinalis and Cliona celata collected at the galician coast (NW Spain). Nat. Prod. Lett. 1996, 8, 15–23. [Google Scholar] [CrossRef]
- Castellanos, L.; Duque, C.; Rodríguez, J.; Jiménez, C. Synthesis of acetylhomoagmatine. Mar. Drugs 2006, 4, 286–289. [Google Scholar] [CrossRef] [Green Version]
- Siodłak, D. α,β-Dehydroamino acids in naturally occurring peptides. Amino Acids 2015, 47, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, N.; Costantini, S.; Costantini, M. Blue-print autophagy: Potential for cancer treatment. Mar. Drugs 2016, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pinteus, S.; Silva, J.; Alves, C.; Horta, A.; Fino, N.; Rodrigues, A.I.; Mendes, S.; Pedrosa, R. Cytoprotective effect of seaweeds with high antioxidant activity from the Peniche coast (Portugal). Food Chem. 2017, 218, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.; Alves, C.; Freitas, R.; Martins, A.; Pinteus, S.; Ribeiro, J.; Gaspar, H.; Alfonso, A.; Pedrosa, R. Antioxidant and neuroprotective potential of the brown seaweed Bifurcaria bifurcata in an in vitro Parkinson’s disease model. Mar. Drugs 2019, 17, 85. [Google Scholar] [CrossRef] [Green Version]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending Applicability of the Oxygen Radical Absorbance Capacity (ORAC-Fluorescein) Assay. Agric. Food Chem. 2003, 52, 48–54. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Yang, E.J.; Yim, E.Y.; Song, G.; Kim, G.O.; Hyun, C.G. Inhibition of nitric oxide production in lipopolysaccharide-activated RAW 264.7 macrophages by Jeju plant extracts. Interdiscip. Toxicol. 2009, 2, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Freitas, R.; Martins, A.; Silva, J.; Alves, C.; Pinteus, S.; Alves, J.; Teodoro, F.; Ribeiro, H.M.; Gonçalves, L.; Petrovski, Ž.; et al. Highlighting the biological potential of the brown seaweed Fucus spiralis for skin applications. Antioxidants 2020, 9, 611. [Google Scholar] [CrossRef]
- Silva, J.; Martins, A.; Alves, C.; Pinteus, S.; Gaspar, H.; Alfonso, A.; Pedrosa, R. Natural Approaches for Neurological Disorders-The Neuroprotective Potential of Codium tomentosum. Molecules 2020, 25, 5478. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | DPPH 1 | FRAP 2 | ORAC 3 |
|---|---|---|---|
| C1 | 96.56 ± 0.85 | 24.32 ± 2.24 | 173.63 ± 7.07 |
| C2 | 95.56 ± 1.63 | 31.93 ± 5.45 | 120.22 ± 5.24 |
| C3 | 94.35 ± 1.02 | 35.41 ± 3.62 | 248.91 ± 6.74 |
| C4 | 89.98 ± 1.18 | 77.63 ± 4.10 | 239.37 ± 7.50 |
| C5 | 98.04 ± 0.71 | 38.54 ± 3.38 | 171.67 ± 6.17 |
| BHT | 39.10 ± 2.97 | 2821.50 ± 63.04 | 136.38 ± 9.09 |
| Fraction | Yield (%) | Cells Viability (% of Control) | ||
|---|---|---|---|---|
| 200 μg·mL−1 | 160 μg·mL−1 | 100 μg·mL−1 | ||
| F1 | 0.08 | - | - | - |
| F2 | 0.05 | - | - | - |
| F3 | 0.11 | - | - | - |
| F4 | 0.63 | - | - | - |
| F5 | 7.29 | 64.41 ± 7.23 * | 118.35 ± 2.83 | 101.00 ± 7.16 |
| F6 | 4.90 | 7.31 ± 2.89 * | 19.13 ± 2.84 * | 46.83 ± 2.09 * |
| F7 | 2.32 | 0.00 ± 0.00 * | 15.18 ± 3.96 * | 36.76 ± 3.46 * |
| F8.1 | 1.47 | 0.00 ± 0.00 * | 4.71 ± 1.36 * | 15.69 ± 1.88 * |
| F8.2 | 5.60 | 0.00 ± 0.00 * | 22.99 ± 3.81 * | 29.84 ± 3.76 * |
| F9 | 3.58 | 0.08 ± 0.09 * | 8.80 ± 1.72 * | 50.19 ± 11.71 * |
| F10 | 0.87 | - | - | - |
| F11 | 7.75 | 24.33 ± 7.65 * | 32.97 ± 5.84 * | 63.64 ± 3.79 * |
| F12 | 7.95 | 4.81 ± 2.07 * | 18.08 ± 4.06 * | 50.59 ± 3.51 * |
| F13 | 6.21 | 8.45 ± 1.71 * | 68.62 ± 2.98 * | 82.61 ± 4.46 |
| F14.1 | 30.25 | 82.50 ± 4.03 * | 79.91 ± 4.09 * | 68.39 ± 8.09 * |
| F14.2 | 14.25 | 82.30 ± 3.13 * | 71.40 ± 1.83 * | 61.43 ± 3.95 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, J.; Gaspar, H.; Silva, J.; Alves, C.; Martins, A.; Teodoro, F.; Susano, P.; Pinteus, S.; Pedrosa, R. Unravelling the Anti-Inflammatory and Antioxidant Potential of the Marine Sponge Cliona celata from the Portuguese Coastline. Mar. Drugs 2021, 19, 632. https://doi.org/10.3390/md19110632
Alves J, Gaspar H, Silva J, Alves C, Martins A, Teodoro F, Susano P, Pinteus S, Pedrosa R. Unravelling the Anti-Inflammatory and Antioxidant Potential of the Marine Sponge Cliona celata from the Portuguese Coastline. Marine Drugs. 2021; 19(11):632. https://doi.org/10.3390/md19110632
Chicago/Turabian StyleAlves, Joana, Helena Gaspar, Joana Silva, Celso Alves, Alice Martins, Fernando Teodoro, Patrícia Susano, Susete Pinteus, and Rui Pedrosa. 2021. "Unravelling the Anti-Inflammatory and Antioxidant Potential of the Marine Sponge Cliona celata from the Portuguese Coastline" Marine Drugs 19, no. 11: 632. https://doi.org/10.3390/md19110632
APA StyleAlves, J., Gaspar, H., Silva, J., Alves, C., Martins, A., Teodoro, F., Susano, P., Pinteus, S., & Pedrosa, R. (2021). Unravelling the Anti-Inflammatory and Antioxidant Potential of the Marine Sponge Cliona celata from the Portuguese Coastline. Marine Drugs, 19(11), 632. https://doi.org/10.3390/md19110632